
Methotrexate Ameliorates Systemic Inflammation and Septic Associated-Lung Damage in a Cecal Ligation and Puncture Septic Rat Model

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
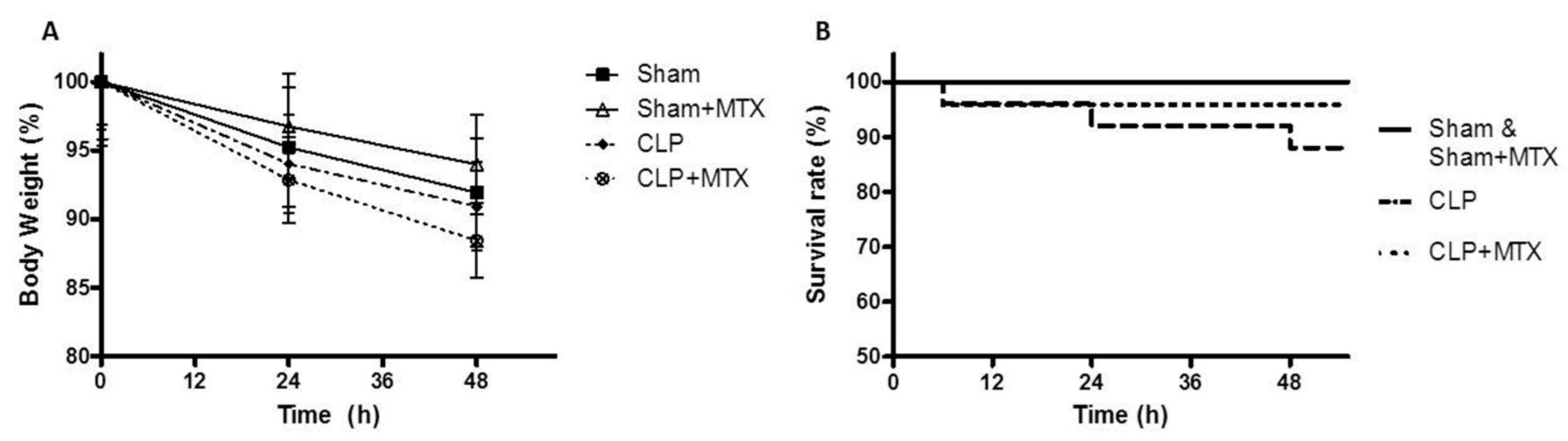
2.1. Survival and Body Weight
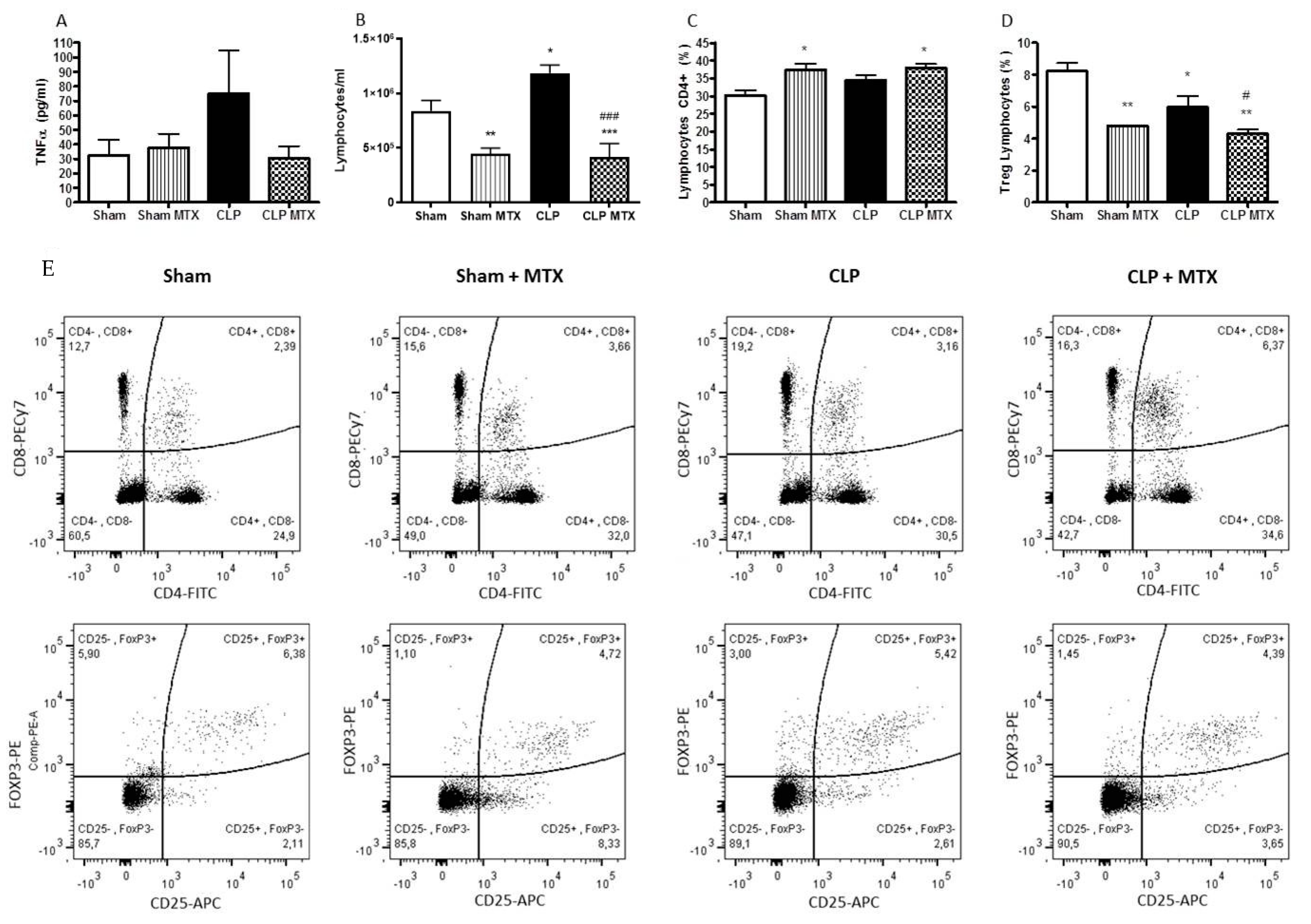
2.2. Systemic Inflammation
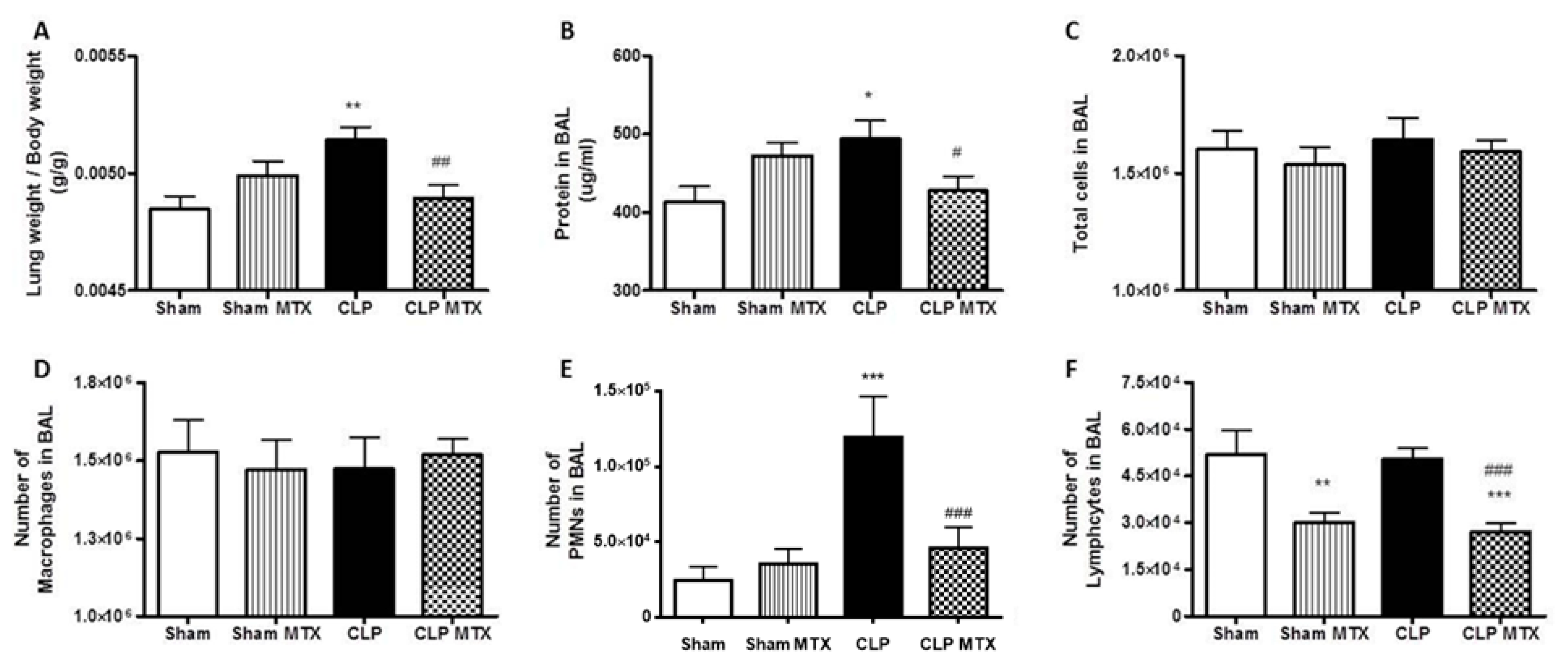
2.3. Lung Weight and Bronchoalveolar Lavage Analysis
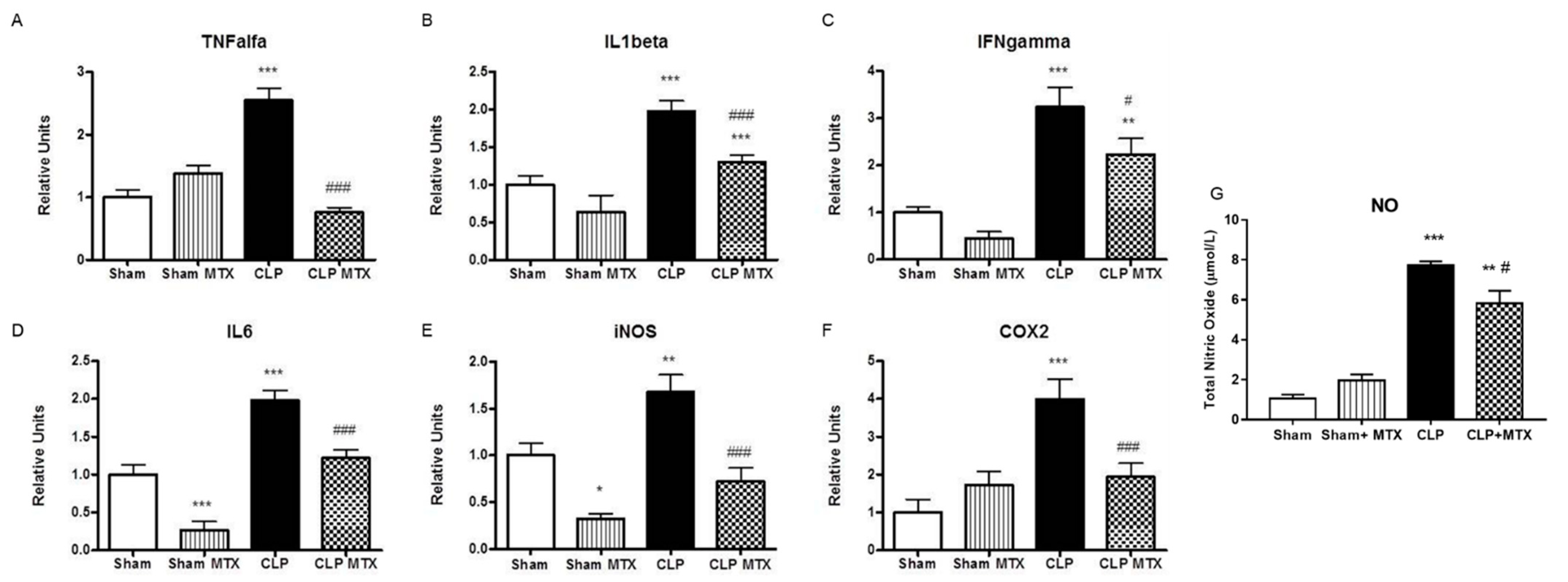
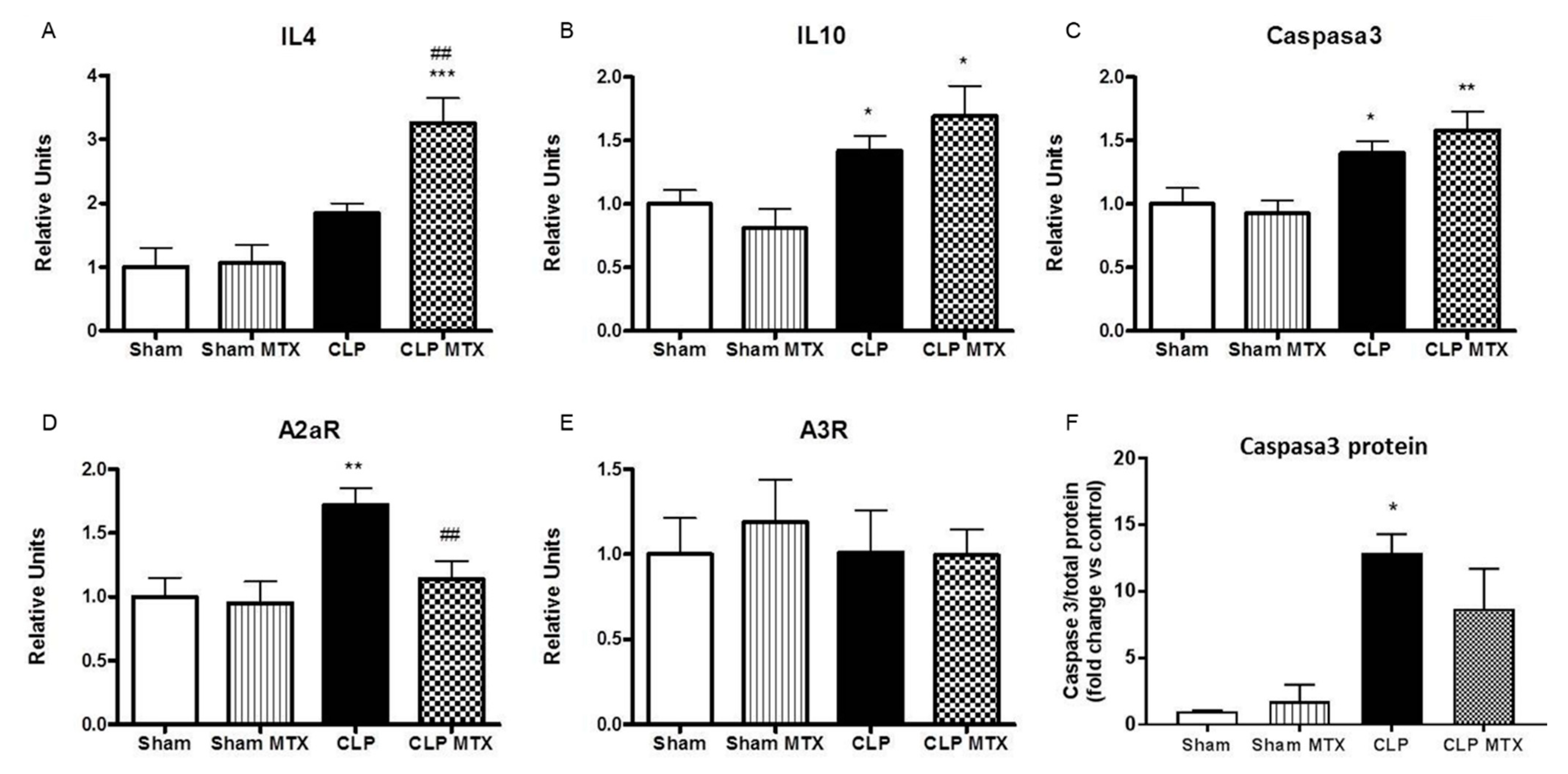
2.4. Inflammatory and Anti-Inflammatory Pathways in Lung
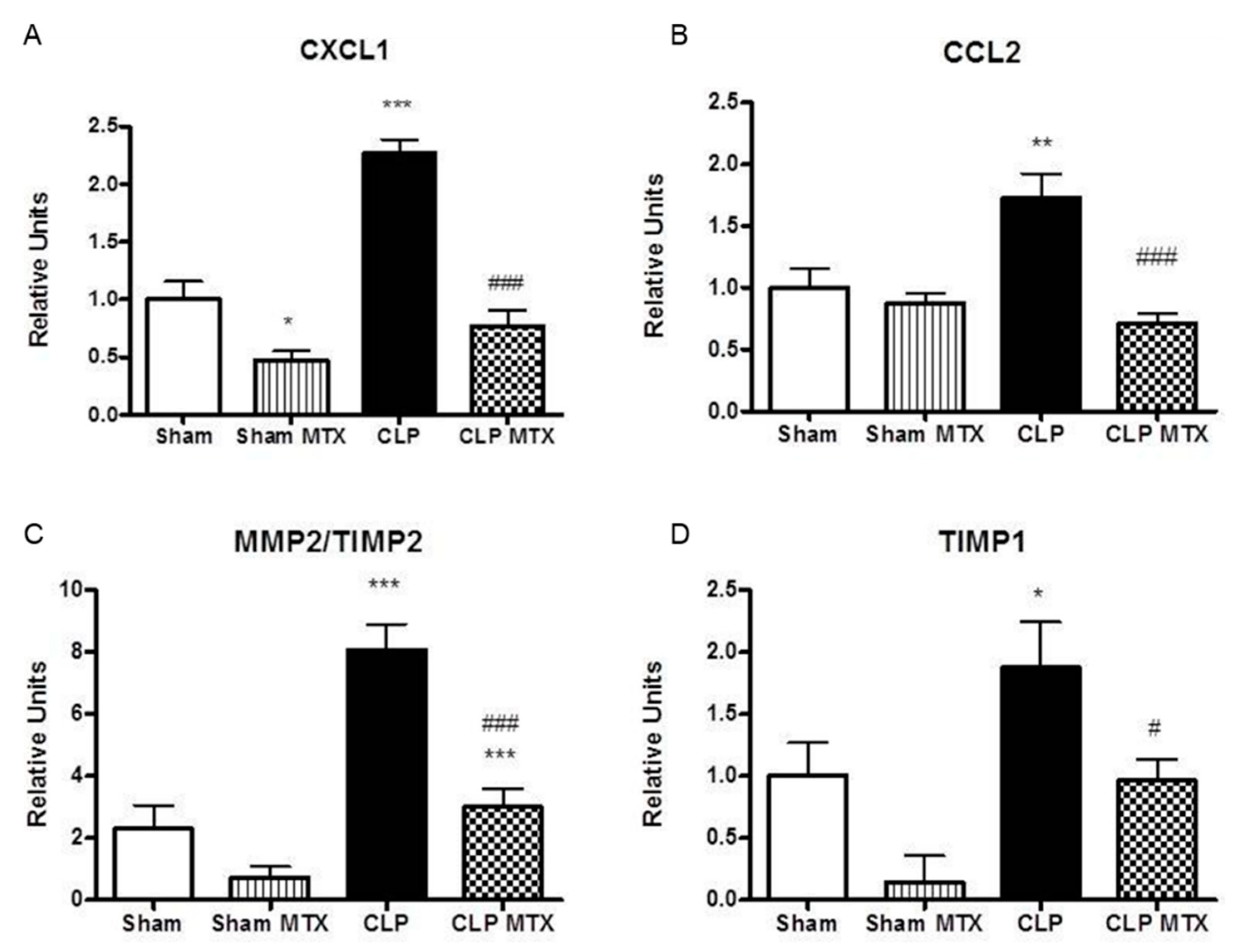
2.5. Recruitment Markers and Metalloproteinase Activation in Lung Tissue
3. Discussion
4. Materials and Methods
4.1. Animal
4.2. Sepsis Induction and Treatment
4.3. RNA Isolation and Gene Expression Analysis by qRT-PCR
4.4. Western Blot of Caspase 3
4.5. Cellular Quantification of Bronchoalveolar Lavage, Cytokine and Total Nitric Oxide Measurement
4.6. Flow Cytometry Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dellinger, R.P.; Levy, M.M.; Rhodes, A.; Annane, D.; Gerlach, H.; Opal, S.M.; Sevransky, J.E.; Sprung, C.L.; Douglas, I.S.; Jaeschke, R.; et al. Surviving Sepsis Campaign: International Guidelines for Management of Severe Sepsis and Septic Shock: 2012. Crit. Care Med. 2013, 41, 580–637. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.M.; Fink, M.P.; Marshall, J.C.; Abraham, E.; Angus, D.; Cook, D.; Cohen, J.; Opal, S.M.; Vincent, J.-L.; Ramsay, G.; et al. 2001 SCCM/ESICM/ACCP/ATS/SIS International Sepsis Definitions Conference. Intensive Care Med. 2003, 29, 530–538. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Reinhart, K.; Daniels, R.; Kissoon, N.; Machado, F.R.; Schachter, R.D.; Finfer, S. Recognizing Sepsis as a Global Health Priority—A WHO Resolution. N. Engl. J. Med. 2017, 377, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.L.; Ferrer, R.; Torrents, E.; Guillamat-Prats, R.; Gomà, G.; Suárez, D.; Álvarez-Rocha, L.; Pozo Laderas, J.C.; Martín-Loeches, I.; Levy, M.M.; et al. Impact of Source Control in Patients With Severe Sepsis and Septic Shock. Crit. Care Med. 2017, 45, 11–19. [Google Scholar] [CrossRef]
- Artigas, A.; Carlet, J.; Ferrer, R.; Niederman, M.; Torres, A. 25th International Symposium on Infections in the Critically Ill Patient. Med. Sci. 2020, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, R.; Martínez, M.L.; Gomà, G.; Suárez, D.; Álvarez-Rocha, L.; de la Torre, M.V.; González, G.; Zaragoza, R.; Borges, M.; Blanco, J.; et al. Improved Empirical Antibiotic Treatment of Sepsis after an Educational Intervention: The ABISS-Edusepsis Study. Crit. Care 2018, 22, 167. [Google Scholar] [CrossRef] [Green Version]
- Seymour, C.W.; Rosengart, M.R. Septic Shock: Advances in Diagnosis and Treatment. JAMA 2015, 314, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Angus, D.C.; van der Poll, T. Severe Sepsis and Septic Shock. N. Engl. J. Med. 2013, 369, 2063. [Google Scholar] [CrossRef]
- Cohen, J.; Vincent, J.-L.; Adhikari, N.K.J.; Machado, F.R.; Angus, D.C.; Calandra, T.; Jaton, K.; Giulieri, S.; Delaloye, J.; Opal, S.; et al. Sepsis: A Roadmap for Future Research. Lancet Infect. Dis. 2015, 15, 581–614. [Google Scholar] [CrossRef]
- Dushianthan, A.; Grocott, M.P.W.; Postle, A.D.; Cusack, R. Acute Respiratory Distress Syndrome and Acute Lung Injury. Postgrad. Med. J. 2011, 87, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Rossaint, J.; Zarbock, A. Pathogenesis of Multiple Organ Failure in Sepsis. Crit. Rev. Immunol. 2015, 35, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Stearns-Kurosawa, D.J.; Osuchowski, M.F.; Valentine, C.; Kurosawa, S.; Remick, D.G. The Pathogenesis of Sepsis. Annu. Rev. Pathol. 2011, 6, 19–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- László, I.; Trásy, D.; Molnár, Z.; Fazakas, J. Sepsis: From Pathophysiology to Individualized Patient Care. J. Immunol. Res. 2015, 2015, 510436. [Google Scholar] [CrossRef]
- Mühl, D.; Nagy, B.; Woth, G.; Falusi, B.; Bogár, L.; Weber, G.; Lantos, J. Dynamic Changes of Matrix Metalloproteinases and Their Tissue Inhibitors in Severe Sepsis. J. Crit. Care 2011, 26, 550–555. [Google Scholar] [CrossRef]
- Hoffmann, U.; Bertsch, T.; Dvortsak, E.; Liebetrau, C.; Lang, S.; Liebe, V.; Huhle, G.; Borggrefe, M.; Brueckmann, M. Matrix-Metalloproteinases and Their Inhibitors Are Elevated in Severe Sepsis: Prognostic Value of TIMP-1 in Severe Sepsis. Scand. J. Infect. Dis. 2006, 38, 867–872. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in Sepsis: A Novel Understanding of the Disorder and a New Therapeutic Approach. Lancet Infect. Dis. 2013, 13, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Boomer, J.S.; Green, J.M.; Hotchkiss, R.S. The Changing Immune System in Sepsis: Is Individualized Immuno-Modulatory Therapy the Answer? Virulence 2014, 5, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira, J.C.; Gentile, L.F.; Mathias, B.J.; Efron, P.A.; Brakenridge, S.C.; Mohr, A.M.; Moore, F.A.; Moldawer, L.L. Sepsis Pathophysiology, Chronic Critical Illness, and Persistent Inflammation-Immunosuppression and Catabolism Syndrome. Crit. Care Med. 2017, 45, 253–262. [Google Scholar] [CrossRef]
- van Vught, L.A.; Klein Klouwenberg, P.M.C.; Spitoni, C.; Scicluna, B.P.; Wiewel, M.A.; Horn, J.; Schultz, M.J.; Nürnberg, P.; Bonten, M.J.M.; Cremer, O.L.; et al. Incidence, Risk Factors, and Attributable Mortality of Secondary Infections in the Intensive Care Unit After Admission for Sepsis. JAMA 2016, 315, 1469–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angus, D.C.; Opal, S. Immunosuppression and Secondary Infection in Sepsis: Part, Not All, of the Story. JAMA 2016, 315, 1457–1459. [Google Scholar] [CrossRef]
- Monneret, G.; Venet, F. A Rapidly Progressing Lymphocyte Exhaustion after Severe Sepsis. Crit. Care 2012, 16, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boomer, J.S.; To, K.; Chang, K.C.; Takasu, O.; Osborne, D.F.; Walton, A.H.; Bricker, T.L.; Jarman, S.D.; Kreisel, D.; Krupnick, A.S.; et al. Immunosuppression in Patients Who Die of Sepsis and Multiple Organ Failure. JAMA 2011, 306, 2594–2605. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Chung, C.-S.; Kherouf, H.; Geeraert, A.; Malcus, C.; Poitevin, F.; Bohé, J.; Lepape, A.; Ayala, A.; Monneret, G. Increased Circulating Regulatory T Cells (CD4(+)CD25 (+)CD127 (-)) Contribute to Lymphocyte Anergy in Septic Shock Patients. Intensive Care Med. 2009, 35, 678–686. [Google Scholar] [CrossRef] [Green Version]
- Hotchkiss, R.S.; Tinsley, K.W.; Swanson, P.E.; Schmieg, R.E.; Hui, J.J.; Chang, K.C.; Osborne, D.F.; Freeman, B.D.; Cobb, J.P.; Buchman, T.G.; et al. Sepsis-Induced Apoptosis Causes Progressive Profound Depletion of B and CD4+ T Lymphocytes in Humans. J. Immunol. 2001, 166, 6952–6963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heming, N.; Azabou, E.; Cazaumayou, X.; Moine, P.; Annane, D. Sepsis in the Critically Ill Patient: Current and Emerging Management Strategies. Expert Rev. Anti-Infect. Ther. 2021, 19, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Aggarwal, B.B. Methotrexate Suppresses NF-KappaB Activation through Inhibition of IkappaBalpha Phosphorylation and Degradation. J. Immunol. 2001, 167, 2911–2920. [Google Scholar] [CrossRef] [Green Version]
- Cronstein, B.N.; Levin, R.I.; Belanoff, J.; Weissmann, G.; Hirschhorn, R. Adenosine: An Endogenous Inhibitor of Neutrophil-Mediated Injury to Endothelial Cells. J. Clin Investig. 1986, 78, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Wollner, A.; Wollner, S.; Smith, J.B. Acting via A2 Receptors, Adenosine Inhibits the Upregulation of Mac-1 (Cd11b/CD18) Expression on FMLP-Stimulated Neutrophils. Am. J. Respir. Cell Mol. Biol. 1993, 9, 179–185. [Google Scholar] [CrossRef]
- Alqarni, A.M.; Zeidler, M.P. How Does Methotrexate Work? Biochem. Soc. Trans. 2020, 48, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, F.; Matsuno, H.; Yudoh, K.; Katayama, R.; Sawai, T.; Uzuki, M.; Kimura, T. Methotrexate Inhibits Rheumatoid Synovitis by Inducing Apoptosis. J. Rheumatol. 2001, 28, 1800–1808. [Google Scholar]
- Wascher, T.C.; Hermann, J.; Brezinschek, H.P.; Brezinschek, R.; Wilders-Truschnig, M.; Rainer, F.; Krejs, G.J. Cell-Type Specific Response of Peripheral Blood Lymphocytes to Methotrexate in the Treatment of Rheumatoid Arthritis. Clin. Investig. 1994, 72, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Quéméneur, L.; Gerland, L.-M.; Flacher, M.; Ffrench, M.; Revillard, J.-P.; Genestier, L. Differential Control of Cell Cycle, Proliferation, and Survival of Primary T Lymphocytes by Purine and Pyrimidine Nucleotides. J. Immunol. 2003, 170, 4986–4995. [Google Scholar] [CrossRef] [Green Version]
- Arslan, A.; Ozcicek, A.; Suleyman, B.; Coban, T.A.; Cimen, F.K.; Nalkiran, H.S.; Kuzucu, M.; Altuner, D.; Cetin, N.; Suleyman, H. Effects of Nimesulide on the Small Intestine Mucositis Induced by Methotrexate in Rats. Exp. Anim. 2016, 65, 329–336. [Google Scholar] [CrossRef]
- Refaat, R.; Salama, M.; Abdel Meguid, E.; El Sarha, A.; Gowayed, M. Evaluation of the Effect of Losartan and Methotrexate Combined Therapy in Adjuvant-Induced Arthritis in Rats. Eur. J. Pharmacol. 2013, 698, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Friebe, D.; Yang, T.; Schmidt, T.; Borg, N.; Steckel, B.; Ding, Z.; Schrader, J. Purinergic Signaling on Leukocytes Infiltrating the LPS-Injured Lung. PLoS ONE 2014, 9, e95382. [Google Scholar] [CrossRef] [PubMed]
- Ali, R.A.; Gandhi, A.A.; Meng, H.; Yalavarthi, S.; Vreede, A.P.; Estes, S.K.; Palmer, O.R.; Bockenstedt, P.L.; Pinsky, D.J.; Greve, J.M.; et al. Adenosine Receptor Agonism Protects against NETosis and Thrombosis in Antiphospholipid Syndrome. Nat. Commun. 2019, 10, 1916. [Google Scholar] [CrossRef] [PubMed]
- Leppkes, M.; Knopf, J.; Naschberger, E.; Lindemann, A.; Singh, J.; Herrmann, I.; Stürzl, M.; Staats, L.; Mahajan, A.; Schauer, C.; et al. Vascular Occlusion by Neutrophil Extracellular Traps in COVID-19. EBioMedicine 2020, 58, 102925. [Google Scholar] [CrossRef] [PubMed]
- Dejager, L.; Pinheiro, I.; Dejonckheere, E.; Libert, C. Cecal Ligation and Puncture: The Gold Standard Model for Polymicrobial Sepsis? Trends Microbiol. 2011, 19, 198–208. [Google Scholar] [CrossRef]
- Rittirsch, D.; Huber-Lang, M.S.; Flierl, M.A.; Ward, P.A. Immunodesign of Experimental Sepsis by Cecal Ligation and Puncture. Nat. Protoc. 2009, 4, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Leone, M.; Viviand, X.; Ayem, M.L.; Guieu, R. High Adenosine Plasma Concentration as a Prognostic Index for Outcome in Patients with Septic Shock. Crit. Care Med. 2000, 28, 3198–3202. [Google Scholar] [CrossRef]
- Capcha, J.M.C.; Moreira, R.S.; Rodrigues, C.E.; Silveira, M.A.D.; Andrade, L.; Gomes, S.A. Using the Cecal Ligation and Puncture Model of Sepsis to Induce Rats to Multiple Organ Dysfunction. Bio Protoc. 2021, 11, e3979. [Google Scholar] [CrossRef]
- Rittirsch, D.; Hoesel, L.M.; Ward, P.A. The disconnect between animal models of sepsis and human sepsis. J. Leukoc. Biol. 2007, 81, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downey, J.S.; Han, J. Cellular Activation Mechanisms in Septic Shock. Front. Biosci. 1998, 3, d468–d476. [Google Scholar] [CrossRef] [Green Version]
- Mercurio, F.; Manning, A.M. Multiple Signals Converging on NF-KappaB. Curr. Opin. Cell Biol. 1999, 11, 226–232. [Google Scholar] [CrossRef]
- Liu, S.F.; Malik, A.B. NF-Kappa B Activation as a Pathological Mechanism of Septic Shock and Inflammation. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 290, L622–L645. [Google Scholar] [CrossRef] [PubMed]
- Kirkebøen, K.A.; Strand, O.A. The Role of Nitric Oxide in Sepsis—An Overview. Acta Anaesthesiol. Scand. 1999, 43, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Parratt, J.R. Nitric Oxide. A Key Mediator in Sepsis and Endotoxaemia? J. Physiol. Pharmacol. 1997, 48, 493–506. [Google Scholar]
- Parratt, J.R. Nitric Oxide in Sepsis and Endotoxaemia. J. Antimicrob. Chemother. 1998, 41, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Salvemini, D.; Korbut, R.; Anggård, E.; Vane, J. Immediate Release of a Nitric Oxide-like Factor from Bovine Aortic Endothelial Cells by Escherichia Coli Lipopolysaccharide. Proc. Natl. Acad. Sci. USA 1990, 87, 2593–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Adcock, I.M.; Old, R.W.; Barnes, P.J.; Evans, T.W. Lipopolysaccharide Treatment in Vivo Induces Widespread Tissue Expression of Inducible Nitric Oxide Synthase MRNA. Biochem. Biophys. Res. Commun. 1993, 196, 1208–1213. [Google Scholar] [CrossRef]
- Benjamim, C.F.; Silva, J.S.; Fortes, Z.B.; Oliveira, M.A.; Ferreira, S.H.; Cunha, F.Q. Inhibition of Leukocyte Rolling by Nitric Oxide during Sepsis Leads to Reduced Migration of Active Microbicidal Neutrophils. Infect. Immun. 2002, 70, 3602–3610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Liu, Z.; Wu, W.; Rozo, C.; Bowdridge, S.; Millman, A.; Van Rooijen, N.; Urban, J.F.; Wynn, T.A.; Gause, W.C. An Essential Role for TH2-Type Responses in Limiting Acute Tissue Damage during Experimental Helminth Infection. Nat. Med. 2012, 18, 260–266. [Google Scholar] [CrossRef]
- Jenkins, S.J.; Ruckerl, D.; Cook, P.C.; Jones, L.H.; Finkelman, F.D.; van Rooijen, N.; MacDonald, A.S.; Allen, J.E. Local Macrophage Proliferation, Rather than Recruitment from the Blood, Is a Signature of TH2 Inflammation. Science 2011, 332, 1284–1288. [Google Scholar] [CrossRef] [Green Version]
- Pelekanou, A.; Tsangaris, I.; Kotsaki, A.; Karagianni, V.; Giamarellou, H.; Armaganidis, A.; Giamarellos-Bourboulis, E.J. Decrease of CD4-Lymphocytes and Apoptosis of CD14-Monocytes Are Characteristic Alterations in Sepsis Caused by Ventilator-Associated Pneumonia: Results from an Observational Study. Crit. Care 2009, 13, R172. [Google Scholar] [CrossRef] [Green Version]
- Gagliani, N.; Amezcua Vesely, M.C.; Iseppon, A.; Brockmann, L.; Xu, H.; Palm, N.W.; de Zoete, M.R.; Licona-Limón, P.; Paiva, R.S.; Ching, T.; et al. Th17 Cells Transdifferentiate into Regulatory T Cells during Resolution of Inflammation. Nature 2015, 523, 221–225. [Google Scholar] [CrossRef]
- Tian, H.; Cronstein, B.N. Understanding the Mechanisms of Action of Methotrexate: Implications for the Treatment of Rheumatoid Arthritis. Bull. NYU Hosp. Jt. Dis. 2007, 65, 168–173. [Google Scholar] [PubMed]
- Koshiba, M.; Rosin, D.L.; Hayashi, N.; Linden, J.; Sitkovsky, M.V. Patterns of A2A Extracellular Adenosine Receptor Expression in Different Functional Subsets of Human Peripheral T Cells. Flow Cytometry Studies with Anti-A2A Receptor Monoclonal Antibodies. Mol. Pharmacol. 1999, 55, 614–624. [Google Scholar]
- Antonioli, L.; Fornai, M.; Blandizzi, C.; Pacher, P.; Haskó, G. Adenosine Signaling and the Immune System: When a Lot Could Be Too Much. Immunol. Lett. 2019, 205, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Naamani, O.; Chaimovitz, C.; Douvdevani, A. Pharmacological Preconditioning with Adenosine A(1) Receptor Agonist Suppresses Cellular Immune Response by an A(2A) Receptor Dependent Mechanism. Int. Immunopharmacol. 2014, 20, 205–212. [Google Scholar] [CrossRef]
- Belikoff, B.; Hatfield, S.; Sitkovsky, M.; Remick, D.G. Adenosine Negative Feedback on A2A Adenosine Receptors Mediates Hyporesponsiveness in Chronically Septic Mice. Shock 2011, 35, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Németh, Z.H.; Csóka, B.; Wilmanski, J.; Xu, D.; Lu, Q.; Ledent, C.; Deitch, E.A.; Pacher, P.; Spolarics, Z.; Haskó, G. Adenosine A2A Receptor Inactivation Increases Survival in Polymicrobial Sepsis. J. Immunol. 2006, 176, 5616–5626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belikoff, B.G.; Hatfield, S.; Georgiev, P.; Ohta, A.; Lukashev, D.; Buras, J.A.; Remick, D.G.; Sitkovsky, M. A2B Adenosine Receptor Blockade Enhances Macrophage-Mediated Bacterial Phagocytosis and Improves Polymicrobial Sepsis Survival in Mice. J. Immunol. 2011, 186, 2444–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.S.; Kurtz, C.C.; Wilson, J.M.; Burnette, B.R.; Wiznerowicz, E.B.; Ross, W.G.; Rieger, J.M.; Figler, R.A.; Linden, J.; Crowe, S.E.; et al. A2A Adenosine Receptor (AR) Activation Inhibits pro-Inflammatory Cytokine Production by Human CD4+ Helper T Cells and Regulates Helicobacter-Induced Gastritis and Bacterial Persistence. Mucosal Immunol. 2009, 2, 232–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillitzer, R.; Goebeler, M. Chemokines in Cutaneous Wound Healing. J. Leukoc. Biol. 2001, 69, 513–521. [Google Scholar]
- Gordon, J.R.; Li, F.; Zhang, X.; Wang, W.; Zhao, X.; Nayyar, A. The Combined CXCR1/CXCR2 Antagonist CXCL8(3-74)K11R/G31P Blocks Neutrophil Infiltration, Pyrexia, and Pulmonary Vascular Pathology in Endotoxemic Animals. J. Leukoc. Biol. 2005, 78, 1265–1272. [Google Scholar] [CrossRef]
- Sônego, F.; Alves-Filho, J.C.; Cunha, F.Q. Targeting Neutrophils in Sepsis. Expert Rev. Clin. Immunol. 2014, 10, 1019–1028. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Target | Primer Sequence (Forward) | Primer Sequence (Reverse) |
|---|---|---|
| GAPDH | 5-CTGTGTCTTTCCGCTGTTTTC-3 | 5-TGTGCTGTGCTTATGGTCTCA-3 |
| IL1β | 5-AAAAATGCCTCGTGCTGTCT-3 | 5-TCGTTGCTTGTCTCTCCTTG-3 |
| TNFα | 5-AACTCCCAGAAAAGCAAGCA-3 | 5-CGAGCAGGAATGAGAAGAGG-3 |
| iNOS | 5-CTTGGAGCGAGTTGTGGATT-3 | 5-GGTGGGAGGGGTAGTGATG-3 |
| IL-6 | 5-CTGCTCTGGTCTTCTGGAGT-3 | 5-GGTCTTGGTCCTTAGCCACT-3 |
| Caspase3 | 5-CCATGTGTGAACTTGGTTGG-3 | 5-TCAACAATTTGAGGCTGCTG-3 |
| CCL2 | 5-GCTGCTACTCATTCACTGGC-3 | 5-GGTGCTGAAGTCCTTAGGGT-3 |
| IL10 | 5-CATCCGGGGTGACAATAA-3 | 5-TGTCCAGCTGGTCCTTCT-3 |
| A3R | 5-TTTACGGTCGGGAGTTCAAG-3 | 5-AGGGTTCATCATGGAGTTCG-3 |
| A2aR | 5-CCTCTTCTTCGCCTGTTTTG-3 | 5-GTTCCCGTCTTTCTGACTGC-3 |
| TIMP1 | 5-GGTTCCCTGGCATAATCTGA-3 | 5-GTCATCGAGACCCCAAGGTA-3 |
| TIMP2 | 5-CAAGTTCTTTGCCTGCATCA-3 | 5-GTTTCCAGGAAGGGATGTCA-3 |
| IFNγ | 5-GAACTGGCAAAAGGACGGTA-3 | 5-GGATCTGTGGGTTGTTCACC-3 |
| COX2 | 5-CTGAGGGGTTACCACTTCCA-3 | 5-TGAGCAAGTCCGTGTTCAAG-3 |
| MMP2 | 5-ACACTGGGACCTGTCACTCC-3 | 5-ACACGGCATCAATCTTTTCC-3 |
| MMP9 | 5-CACTGTAACTGGGGGCAACT-3 | 5-CACTTCTTGTCAGCGTCGAA-3 |
| Arg1 | 5-GGGAAGACACCAGAGGAGGT-3 | 5-TGATGCCCCAGATGACTTTT-3 |
| IL12 | 5-CATCTGCTGCTCCACAAGAA-3 | 5-GAGACTCAGGGGAACTGCTG-3 |
| IL4 | 5-TCCTTACGGCAACAAGGAAC-3 | 5-GTGAGTTCAGACCGCTGACA-3 |
| MMP9 | 5-CACTGTAACTGGGGGCAACT-3 | 5-CACTTCTTGTCAGCGTCGAA-3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bringué, J.; Guillamat-Prats, R.; Martinez, M.L.; Torrents, E.; Camprubí-Rimblas, M.; Blanch, L.; Artigas, A. Methotrexate Ameliorates Systemic Inflammation and Septic Associated-Lung Damage in a Cecal Ligation and Puncture Septic Rat Model. Int. J. Mol. Sci. 2021, 22, 9612. https://doi.org/10.3390/ijms22179612
Bringué J, Guillamat-Prats R, Martinez ML, Torrents E, Camprubí-Rimblas M, Blanch L, Artigas A. Methotrexate Ameliorates Systemic Inflammation and Septic Associated-Lung Damage in a Cecal Ligation and Puncture Septic Rat Model. International Journal of Molecular Sciences. 2021; 22(17):9612. https://doi.org/10.3390/ijms22179612
Chicago/Turabian StyleBringué, Josep, Raquel Guillamat-Prats, Maria Luisa Martinez, Eva Torrents, Marta Camprubí-Rimblas, Lluís Blanch, and Antonio Artigas. 2021. "Methotrexate Ameliorates Systemic Inflammation and Septic Associated-Lung Damage in a Cecal Ligation and Puncture Septic Rat Model" International Journal of Molecular Sciences 22, no. 17: 9612. https://doi.org/10.3390/ijms22179612