
Genome-Wide Identification and Expression Analysis of BBX Transcription Factors in Iris germanica L.

Abstract
:1. Introduction
2. Results
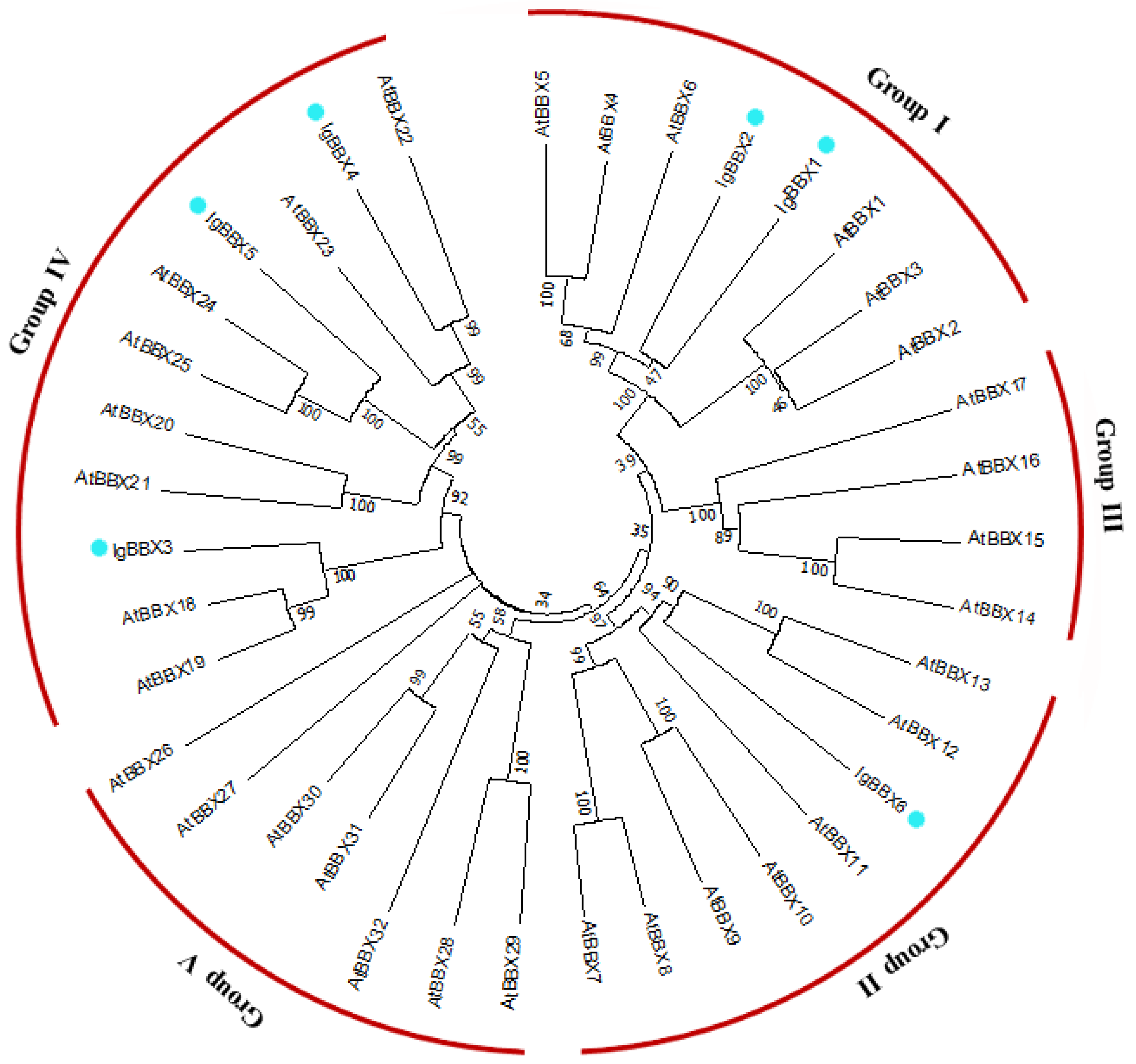
2.1. Identification and Phylogenetic Analysis of IgBBX Genes in I. germanica
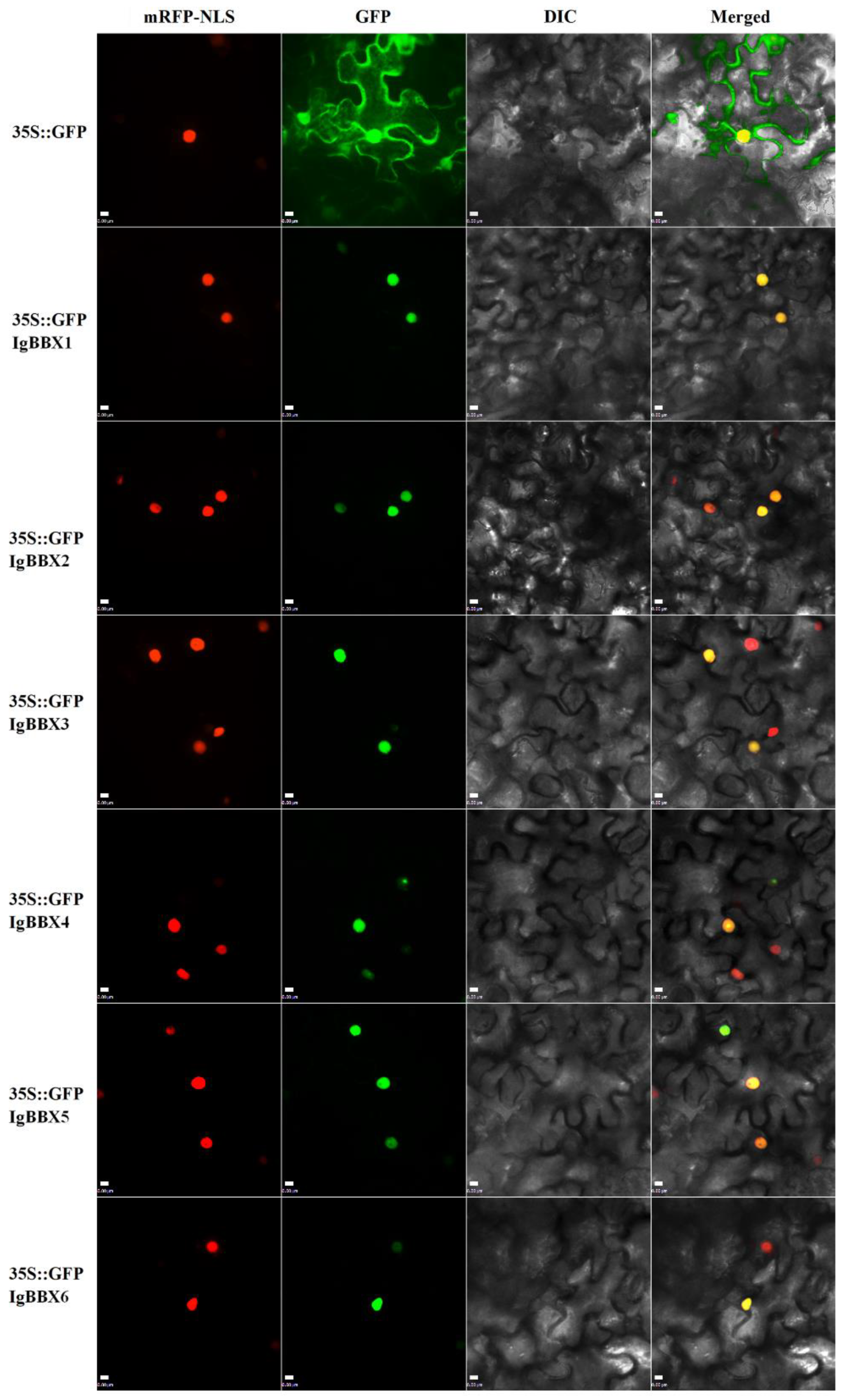
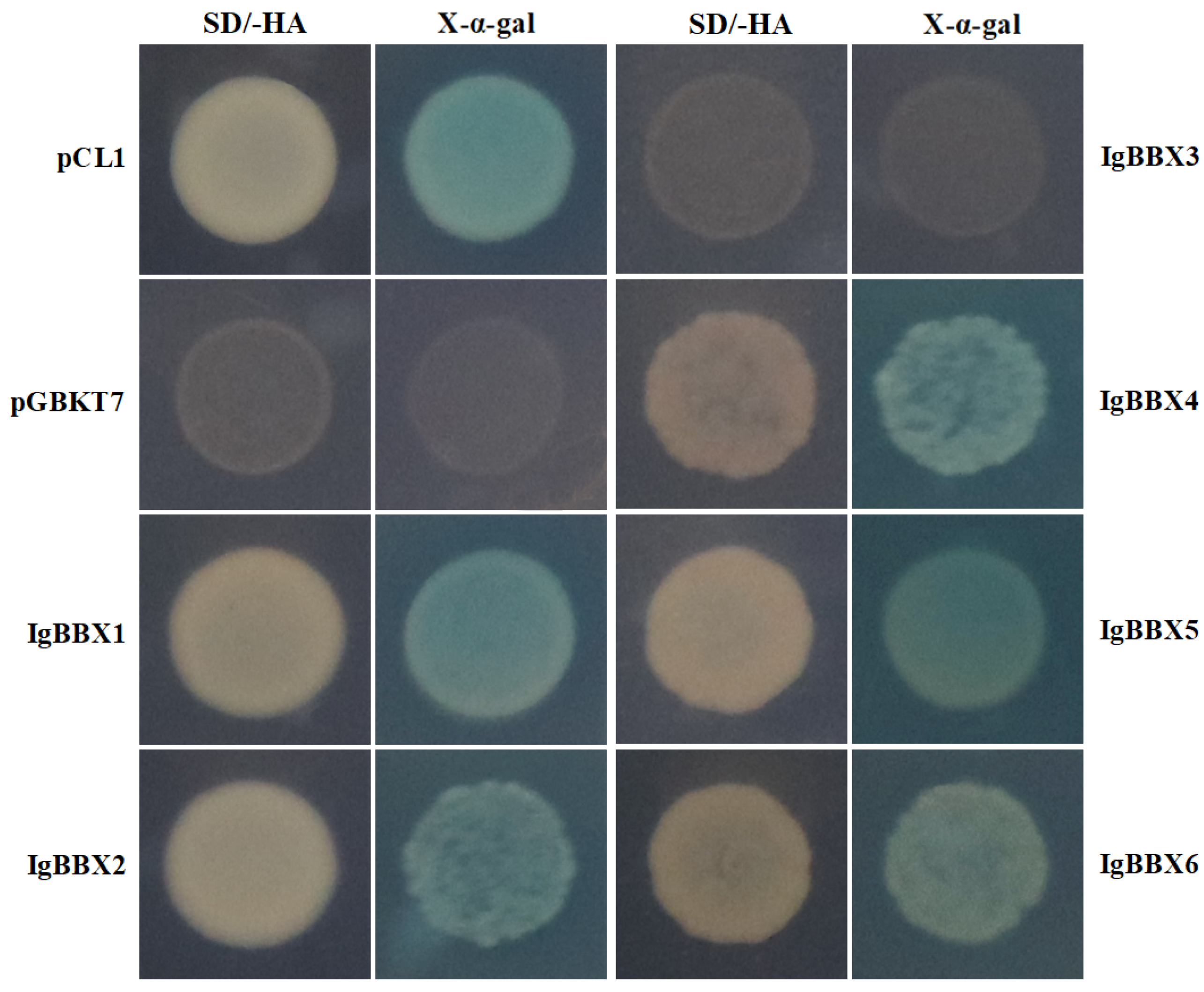
2.2. Subcellular Localization and Transcription Activation of IgBBX Genes
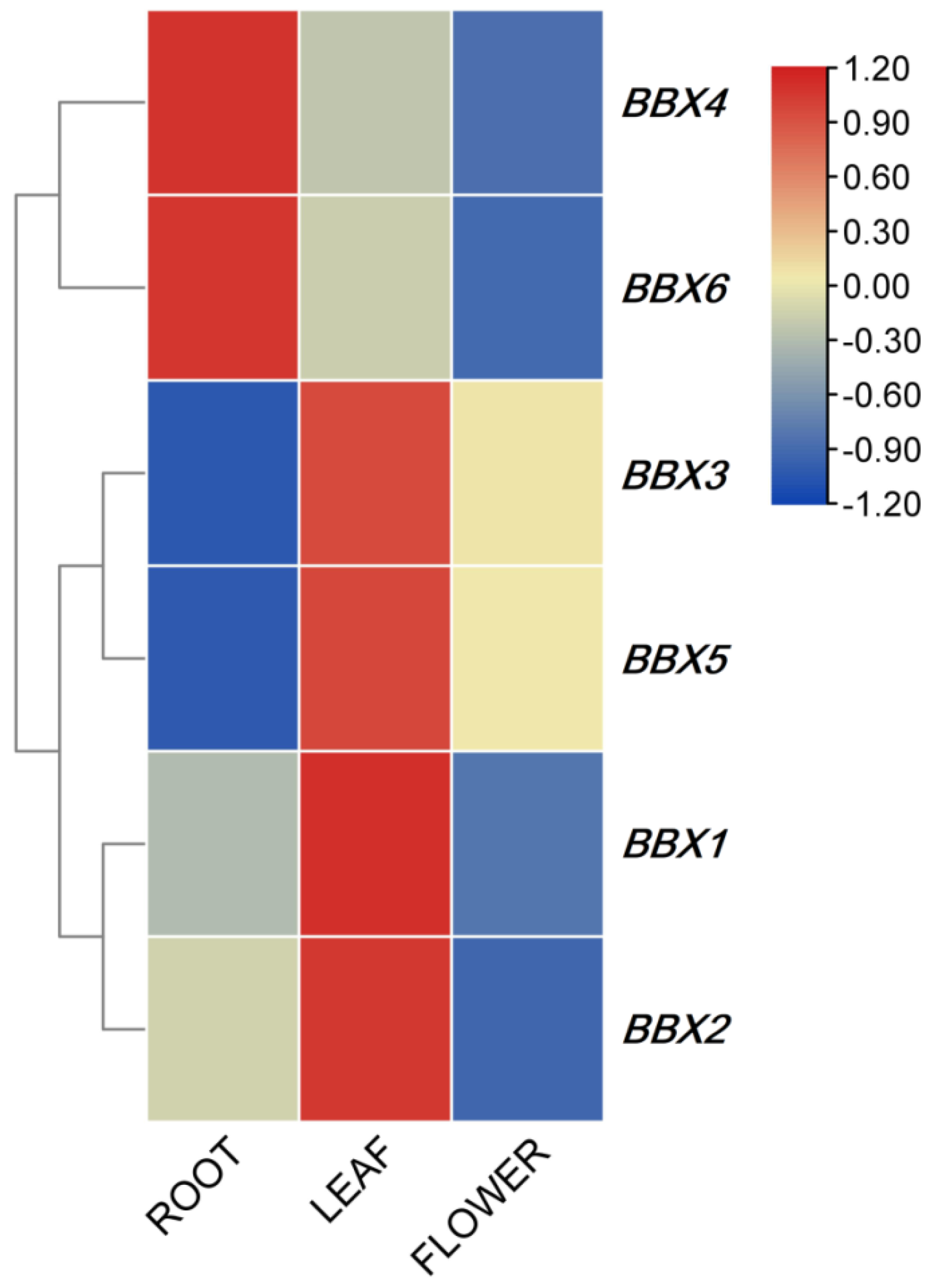
2.3. Expression Profile of IgBBX in Different Plant Tissues
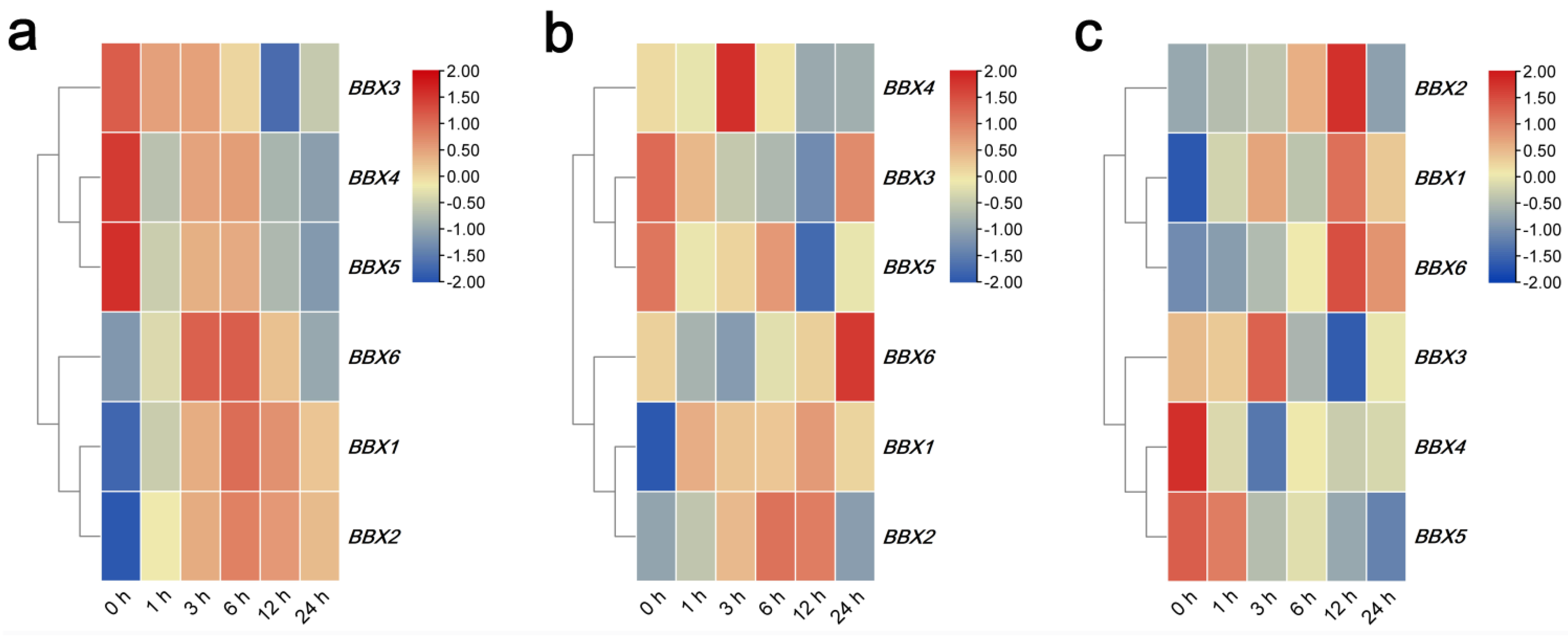
2.4. Expression Profile of IgBBX Genes after ABA, MeJA and SA Phytohormones Treatment
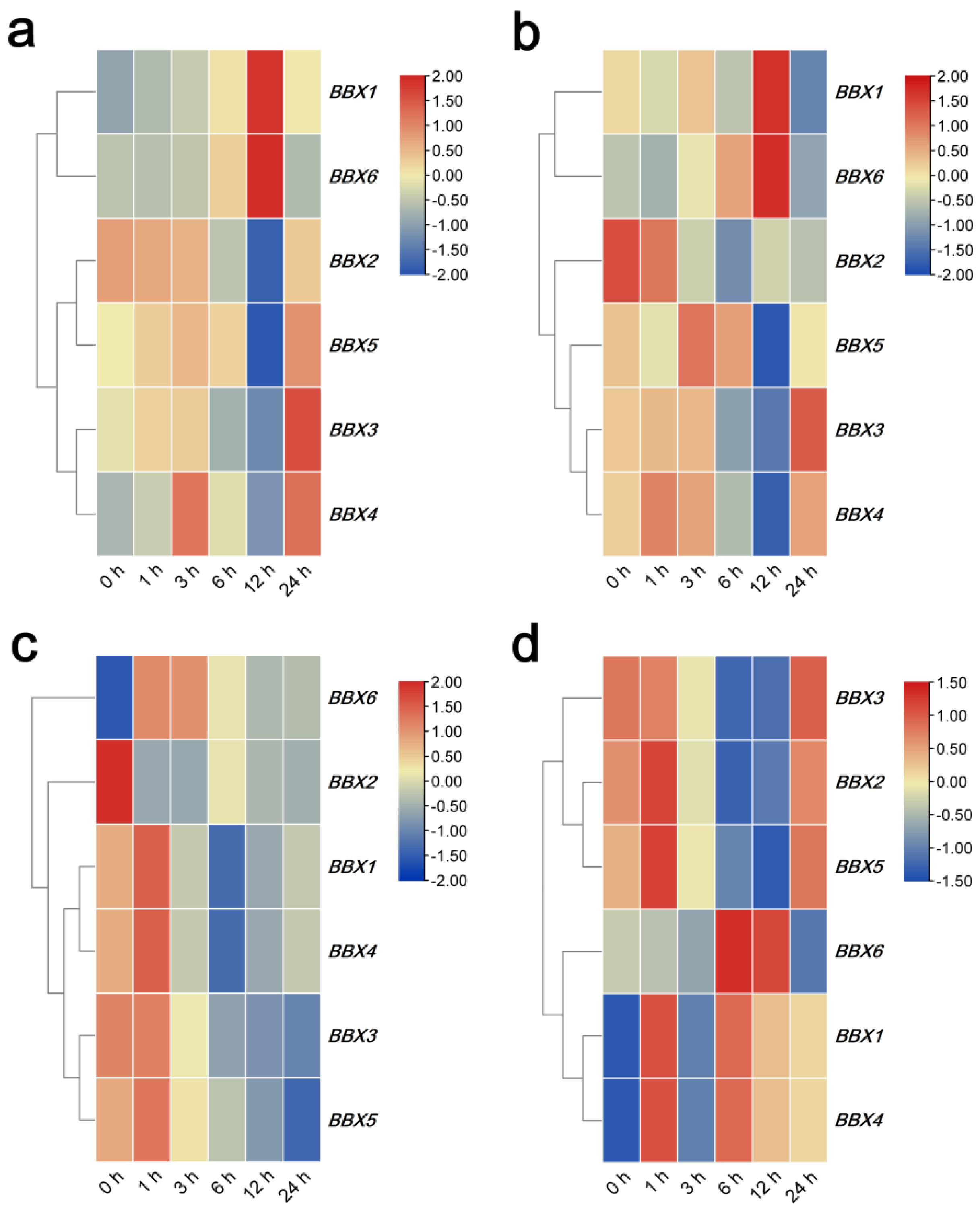
2.5. Transcription Profiling of IgBBX Genes under Abiotic Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatments
4.2. Isolation and Sequencing of Full-Length IgBBX cDNAs
4.3. Sequence Alignment and Phylogenetic Tree Construction
4.4. Subcellular Localization of IgBBX Genes
4.5. Transactivation Activity Assay of IgBBX Genes
4.6. Transcription Analysis of the IgBBX Genes
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–470. [Google Scholar] [CrossRef]
- Ping, Q.; Cheng, P.L.; Huang, F.; Ren, L.P.; Cheng, H.; Guan, Z.Y.; Fang, W.M.; Chen, S.M.; Chen, F.D.; Jiang, J.F. The heterologous expression in Arabidopsis thaliana of a chrysanthemum gene encoding the BBX family transcription factor CmBBX13 delays flowering. Plant Physiol. Bioch. 2019, 144, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.Y.; Zhang, Y.; Deng, Y.; Chen, G.J.; Yu, Y.X.; Wei, Q. Genomic identifcation and expression analysis of the BBX transcription factor gene family in Petunia hybrida. Mol. Biol. Rep. 2020, 47, 6027–6041. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, X.; Weng, X.; Wang, L.; Xie, W. The rice B-Box zinc finger gene family: Genomic identification, characterization, expression profiling and diurnal analysis. PLoS ONE 2012, 7, e48242. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jiao, C.; Jin, Q.; Lin, Y.; Cai, Y. B-BOX genes: Genome-wide identification, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri Rehd). BMC Plant Biol. 2017, 17, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, R.; Dai, Y.Q.; Chen, X.S.; Wang, X.Y. Genome-wide identification and expression analysis of the B-box gene family in the Apple (Malus domestica Borkh.) genome. Mol. Genet. Genom. 2017, 293, 303–315. [Google Scholar] [CrossRef]
- Chaurasia, A.K.; Patil, H.B.; Azeez, A.; Subramaniam, V.R.; Krishna, B.; Sane, A.P.; Sane, P.V. Molecular characterization of CONSTANS-Like (COL) genes in banana (Musa acuminata L. AAA Group, cv. Grand Nain). Physiol. Mol. Biol Plant 2016, 22, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Min, J.H.; Chung, J.S.; Lee, K.H.; Kim, C.S. The CONSTANS-like 4 transcription factor, AtCOL4, positively regulates abiotic stress tolerance through an abscisic acid-dependent manner in Arabidopsis. J. Integr. Plant Biol. 2015, 57, 313–324. [Google Scholar] [CrossRef]
- An, H.; Roussot, C.; Suárez-López, P.; Corbesier, L.; Vincent, C.; Piñeiro, M.; Hepworth, S.; Mouradov, A.; Justin, S.; Turnbull, C.; et al. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development 2004, 131, 3615–3626. [Google Scholar] [CrossRef] [Green Version]
- Suárez-López, P.; Wheatley, K.; Robson, F.; Onounch, H.; Valverde, F.; Coupland, G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 2001, 416, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Carvallo, M.; Hamilton, E.E.; Preuss, S.; Kay, S.A. Arabidopsis B-BOX32 interacts with CONSTANS-LIKE3 to regulate flowering. Proc. Natl. Acad. Sci. USA 2017, 114, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassidim, M.; Harir, Y.; Yakir, E.; Kron, I.; Green, R.M. Over-expression of CONSTANSLIKE 5 can induce flowering in short-day grown Arabidopsis. Planta 2009, 230, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.F.; Wang, Z.Y. Overexpression of COL9, a CONSTANSLIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J. 2005, 43, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q.; Guthrie, C.; Sarmast, M.K.; Dehesh, K. BBX19 interacts with CONSTANS to repress FLOWERING LOCUS T transcription, defining a flowering time checkpoint in Arabidopsis. Plant Cell 2014, 26, 3589–3602. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Sun, J.; Wang, D.; Bai, S.; Clarke, A.K.; Holm, M. The B-box family gene STO (BBX24) in Arabidopsis thaliana regulates flowering time in different pathways. PLoS ONE 2014, 9, e87544. [Google Scholar] [CrossRef]
- Park, H.Y.; Lee, S.Y.; Seok, H.Y.; Kim, S.H.; Sung, Z.R.; Moon, Y.H. EMF1 interacts with EIP1, EIP6 or EIP9 involved in the regulation of flowering time in Arabidopsis. Plant Cell Physiol. 2011, 52, 1376–1388. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Tu, X.; Zhang, J.; Chen, X.; Rao, L. Heat stress-induced BBX18 negatively regulates the thermotolerance in Arabidopsis. Mol. Biol. Rep. 2013, 40, 2679–2688. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Takano, T. Salt tolerance-related protein STO binds to a Myb transcription factor homologue and confers salt tolerance in Arabidopsis. J. Exp. Bot. 2003, 54, 2231–2237. [Google Scholar] [CrossRef]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z.; Cao, S.; Liu, Y. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol. Genet. Genom. 2012, 287, 495–513. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, C.; Xu, Y.; Wei, Q.; Imtiaz, M.; Lan, H.; Gao, S.; Cheng, L.; Wang, M.; Fei, Z. A zinc finger protein regulates flowering time and abiotic stress tolerance in chrysanthemum by modulating gibberellin biosynthesis. Plant Cell 2014, 26, 2038. [Google Scholar] [CrossRef] [Green Version]
- Kielbowicz-Matuk, A.; Rey, P.; Rorat, T. Interplay between circadian rhythm, time of the day and osmotic stress constraints in the regulation of the expression of a Solanum Double B-box gene. Ann. Bot. 2014, 113, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.Y.; Sun, Y.; Cao, D.M.; Bai, M.Y.; Luo, X.M.; Yang, H.J.; Wei, C.Q.; Zhu, S.W.; Sun, Y.; Chong, K.; et al. BZS1, a B-box protein, promotes photomorphogenesis downstream of both brassinosteroid and light signaling pathways. Mol. Plant 2012, 5, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Zhang, Y.X.; Liu, Q.Q.; Liu, L.Q.; Huang, S.Z.; Yuan, H.Y. Reference gene selection for qRT-PCR normalization in Iris germanica L. Phyton Int. J. Exp. Bot. 2020, 90, 277–290. [Google Scholar]
- Bai, B.; Zhao, J.; Li, Y.P.; Zhang, F.; Zhou, J.J.; Chen, F.; Xie, X.Z. OsBBX14 delays heading date by repressing florigen gene expression under long and short-day conditions in rice. Plant Sci. 2016, 247, 25–34. [Google Scholar] [CrossRef]
- Xu, Y.J.; Zhao, X.; Aiwaili, P.; Mu, X.Y.; Zhao, M.; Zhao, J.; Cheng, L.N.; Ma, C.; Gao, J.P.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Bai, S.L.; Tao, R.Y.; Tang, Y.X.; Yin, L.; Ma, Y.J.; Ni, J.B.; Yan, X.H.; Yang, Q.S.; Wu, Z.Y.; Zeng, Y.L.; et al. BBX16, a B-box protein, positively regulates light-induced anthocyanin accumulation by activating MYB10 in red pear. Plant Biotechnol. J. 2019, 17, 1985–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Z.N.; Wang, X.; Li, Y.; Yu, H.Y.; Li, J.H.; Lu, Y.G.; Li, H.X.; Ouyang, B. Genomic organization, phylogenetic and expression analysis of the B-BOX gene family in Tomato. Front. Plant Sci. 2016, 7, 1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, R.; Kronmiller, B.; Maszle, D.R.; Coupland, G.; Holm, M.; Mizuno, T.; Wu, S.H. The Arabidopsis B-box zinc finger family. Plant Cell 2009, 21, 3416–3420. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Sun, J.J.; Jiang, A.Q.; Bai, M.J.; Fan, C.G.; Liu, J.Y.; Ning, G.G.; Wang, C.Q. Alternate expressions of CONSTANS-LIKE 4 in short days and CONSTANS in long days facilitate day-neutral response in Rosa chinensis. J. Exp. Bot. 2020, 71, 4057–4068. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Huang, F.; Liu, Y.N.; Cheng, P.L.; Guan, Z.Y.; Fang, W.M.; Chen, S.M.; Chen, F.D.; Jiang, J.F. Constitutive expression of the chrysanthemum CmBBX29 delays flowering time in transgenic Arabidopsis. Can. J. Plant Sci. 2019, 100, 86–94. [Google Scholar] [CrossRef]
- Sheng, P.K.; Wu, F.Q.; Tan, J.J.; Zhang, H.; Ma, W.W.; Chen, L.P.; Wang, J.C.; Wang, J.; Zhu, S.S.; Guo, X.P.; et al. A CONSTANS-like transcriptional activator, OsCOL13, functions as a negative regulator of flowering downstream of OsphyB and upstream of Ehd1 in rice. Plant Mol. Biol. 2016, 92, 209–222. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Liu, X.; Li, R.; Dai, Y.Q.; Yuan, L.; Sun, Q.H.; Zhang, S.Z.; Wang, X.Y. A B-box zinc finger protein, MdBBX10, enhanced salt and drought stresses tolerance in Arabidopsis. Plant Mol. Biol. 2019, 99, 437–447. [Google Scholar] [CrossRef]
- Zou, Z.; Wang, R.; Wang, R.; Yang, S.; Yang, Y. Genome-wide identification, phylogenetic analysis, and expression profiling of the BBX family genes in pear. J. Hortic. Sci. Biotechnol. 2018, 93, 37–50. [Google Scholar] [CrossRef]
- Liu, Y.N.; Chen, H.; Ping, Q.; Zhang, Z.X.; Guan, Z.Y.; Fang, W.M.; Chen, S.M.; Chen, F.D.; Jiang, J.F.; Zhang, F. The heterologous expression of CmBBX22 delays leaf senescence and improves drought tolerance in Arabidopsis. Plant Cell Rep. 2019, 38, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.R.; Wang, P.P.; Chen, J.Q.; Li, C.J.; Wang, Y.Z.; Yuan, Y.B.; Fang, J.G.; Leng, X.P. Genome-wide identification and analysis of B-BOX gene family in grapevine reveal its potential functions in berry development. BMC Plant Biol. 2020, 20, 72. [Google Scholar] [CrossRef]
- Gao, C.Y.; Li, P.L.; Song, A.P.; Wang, H.B.; Wang, Y.J.; Ren, L.P.; Qi, X.Y.; Chen, F.D.; Jiang, J.F.; Chen, S.M. Isolation and characterization of six AP2/ERF transcription factor genes in Chrysanthemum nankingense. Int. J. Mol. Sci. 2015, 16, 2052–2065. [Google Scholar] [CrossRef] [Green Version]
- Song, A.P.; Gao, T.W.; Li, P.L.; Chen, S.M.; Guan, Z.Y.; Wu, D.; Xin, J.J.; Fang, Q.Q.; Zhao, K.K.; Chen, F.D. Transcriptome-Wide identification and expression profiling of the DOF transcription factor gene family in Chrysanthemum morifolium. Front. Plant Sci. 2016, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Ricachenevsky, F.K.; Sperotto, R.A.; Menguer, P.K.; Fett, J.P. Identification of Fe-excess-induced genes in rice shoots reveals a WRKY transcription factor responsive to Fe, drought and senescence. Mol. Biol. Rep. 2010, 37, 3735–3745. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.W.; Zhang, W.W.; Song, A.P.; An, C.; Xin, J.J.; Jiang, J.F.; Guan, Z.Y.; Chen, F.D.; Chen, S.M. Phylogenetic and transcriptional analysis of chrysanthemum GRAS transcription factors. Biol. Plant. 2018, 62, 711–720. [Google Scholar] [CrossRef]
- Alaey, M.; Babalar, M.; Naderi, R.; Kafi, M. Effect of pre- and post-harvest salicylic acid treatment on physio-chemical attributes in relation to vase-life of rose cut flowers. Postharvest Biol. Technol. 2011, 61, 91–94. [Google Scholar] [CrossRef]
- Gu, C.S.; Xu, S.; Wang, Z.Q.; Liu, L.Q.; Zhang, Y.X.; Deng, Y.M.; Huang, S.Z. De novo sequencing, assembly, and analysis of Iris lactea var. chinensis roots’ transcriptome in response to salt stress. Plant Physiol. Biochem. 2018, 125, 1–12. [Google Scholar] [CrossRef]
- Gu, C.S.; Liu, L.Q.; Deng, Y.M.; Zhang, Y.X.; Wang, Z.Q.; Yuan, H.Y.; Huang, S.Z. De novo characterization of the Iris lactea var. chinensis transcriptome and an analysis of genes under cadmium or lead exposure. Ecotoxicol. Environ. Saf. 2017, 144, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.Y.; He, K.; Liu, D.; Bai, S.N.; Gu, X.C.; Wei, L.P.; Luo, J.C. DATF: A database of Arabidopsis transcription factors. Bioinformatics 2005, 21, 2568–2569. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.J.; Sheng, L.P.; Zhang, H.R.; Du, X.P.; An, C.; Xia, X.L.; Chen, F.D.; Jiang, J.F.; Chen, S.M. CmMYB19 over-expression improves aphid tolerance in chrysanthemum by promoting lignin synthesis. Int. J. Mol. Sci. 2017, 18, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- De Hoon, M.J.; Imoto, S.; Nolan, J.; Miyano, S. Open source clustering software. Bioinformatics 2004, 20, 1453–1454. [Google Scholar] [CrossRef] [Green Version]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | GenBank Accession No. | cDNA Length (bp) | Amino Acids Length (aa) | AtBBX Orthologs | Locus Name | E-Value |
|---|---|---|---|---|---|---|
| IgBBX1 | MW357644 | 1176 | 312 | AtBBX6 | AT5G57660 | 2 × 10−52 |
| IgBBX2 | MW357645 | 987 | 328 | AtBBX4 | AT2G24790 | 2 × 10−76 |
| IgBBX3 | MW357646 | 749 | 197 | AtBBX19 | AT4G38960 | 1 × 10−74 |
| IgBBX4 | MW357647 | 919 | 192 | AtBBX22 | AT1G78600 | 3 × 10−76 |
| IgBBX5 | MW357648 | 894 | 246 | AtBBX24 | AT1G06040 | 2 × 10−86 |
| IgBBX6 | MW357649 | 1478 | 475 | AtBBX12 | AT2G33500 | 1 × 10−63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, Y.; Liu, Q.; Zhang, T.; Chong, X.; Yuan, H. Genome-Wide Identification and Expression Analysis of BBX Transcription Factors in Iris germanica L. Int. J. Mol. Sci. 2021, 22, 8793. https://doi.org/10.3390/ijms22168793
Wang Y, Zhang Y, Liu Q, Zhang T, Chong X, Yuan H. Genome-Wide Identification and Expression Analysis of BBX Transcription Factors in Iris germanica L. International Journal of Molecular Sciences. 2021; 22(16):8793. https://doi.org/10.3390/ijms22168793
Chicago/Turabian StyleWang, Yinjie, Yongxia Zhang, Qingquan Liu, Ting Zhang, Xinran Chong, and Haiyan Yuan. 2021. "Genome-Wide Identification and Expression Analysis of BBX Transcription Factors in Iris germanica L." International Journal of Molecular Sciences 22, no. 16: 8793. https://doi.org/10.3390/ijms22168793