
Regulation of Proteolytic Activity to Improve the Recovery of Macrobrachium rosenbergii Nodavirus Capsid Protein

 , , and
, , and
Abstract
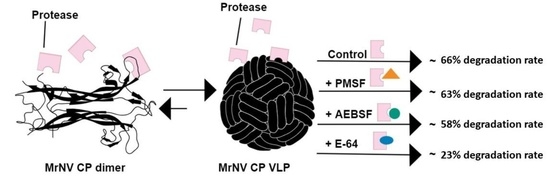
:
1. Introduction
2. Results
2.1. Edman Degradation Sequencing
2.2. Identification of Proteases That May Be Responsible for the Degradation of Recombinant MrNV CP
2.3. SDS-PAGE Densitometry Analysis with ImageJ
3. Discussion
4. Materials and Methods
4.1. Expression and Purification of MrNV CP
4.2. Identification of Proteolytic Cleavage Site by Edman Degradation Sequencing
4.3. Identification of Proteases That May Be Responsible for the Degradation of MrNV CP
4.4. Inhibitory Study of MrNV CP Using Protease Inhibitors
4.5. SDS-PAGE Densitometry Analysis by ImageJ
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arcier, J.M.; Herman, F.; Lightner, D.V.; Redman, R.M.; Mari, J.; Bonami, J.R. A viral disease associated with mortalities in hatchery-reared postlarvae of the giant freshwater prawn Macrobrachium rosenbergii. Dis. Aquat. Organ. 1999, 38, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Qian, D.; Shi, Z.; Zhang, S.; Cao, Z.; Liu, W.; Li, L.; Xie, Y.; Cambournac, I.; Bonami, J.R. Extra small virus-like particles (XSV) and nodavirus associated with whitish muscle disease in the giant freshwater prawn, Macrobrachium rosenbergii. J. Fish Dis. 2003, 26, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.S.; Yoganandhan, K.; Widada, J.S.; Bonami, J.R. Experimental transmission and tissue tropism of Macrobrachium rosenbergii nodavirus (MrNV) and its associated extra small virus (XSV). Dis. Aquat. Org. 2004, 62, 191–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoganandhan, K.; Leartvibhas, M.; Sriwongpuk, S.; Limsuwan, C. White tail disease of the giant freshwater prawn Macrobrachium rosenbergii in Thailand. Dis. Aquat. Org. 2006, 69, 255–258. [Google Scholar] [CrossRef] [Green Version]
- Saedi, T.A.; Moeini, H.; Tan, W.S.; Yusoff, K.; Daud, H.M.; Chu, K.B.; Tan, S.G.; Bhassu, S. Detection and phylogenetic profiling of nodavirus associated with white tail disease in Malaysian Macrobrachium rosenbergii de Man. Mol. Biol. Rep. 2012, 39, 5785–5790. [Google Scholar] [CrossRef]
- Goh, Z.H.; Tan, S.G.; Bhassu, S.; Tan, W.S. Virus-like particles of Macrobrachium rosenbergii nodavirus produced in bacteria. J. Virol. Methods 2011, 175, 74–79. [Google Scholar] [CrossRef]
- Ho, K.L.; Kueh, C.L.; Beh, P.L.; Tan, W.S.; Bhella, D. Cryo-electron microscopy structure of the Macrobrachium rosenbergii nodavirus capsid at 7 angstroms resolution. Sci. Rep. 2017, 7, 2083. [Google Scholar] [CrossRef] [Green Version]
- Jariyapong, P.; Chotwiwatthanakun, C.; Somrit, M.; Jitrapakdee, S.; Xing, L.; Cheng, H.R.; Weerachatyanukul, W. Encapsulation and delivery of plasmid DNA by virus-like nanoparticles engineered from Macrobrachium rosenbergii nodavirus. Virus Res. 2014, 179, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Thong, Q.X.; Biabanikhankahdani, R.; Ho, K.L.; Alitheen, N.B.; Tan, W.S. Thermally responsive virus-like particle for targeted delivery of cancer drug. Sci. Rep. 2019, 9, 3945. [Google Scholar] [CrossRef] [Green Version]
- Thong, Q.X.; Wong, C.L.; Ooi, M.K.; Kueh, C.L.; Ho, K.L.; Alitheen, N.B.; Tan, W.S. Peptide inhibitors of Macrobrachium rosenbergii nodavirus. J. Gen. Virol. 2018, 99, 1227–1238. [Google Scholar] [CrossRef]
- Yong, C.Y.; Yeap, S.K.; Goh, Z.H.; Ho, K.L.; Omar, A.R.; Tan, W.S. Induction of humoral and cell-mediated immune responses by hepatitis b virus epitope displayed on the virus-like particles of prawn nodavirus. Appl. Environ. Microbiol. 2015, 81, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, C.Y.; Yeap, S.K.; Ho, K.L.; Omar, A.R.; Tan, W.S. Potential recombinant vaccine against influenza A virus based on M2e displayed on nodaviral capsid nanoparticles. Int. J. Nanomed. 2015, 10, 2751–2763. [Google Scholar] [CrossRef] [Green Version]
- Ong, H.K.; Yong, C.Y.; Tan, W.S.; Yeap, S.K.; Omar, A.R.; Razak, M.A.; Ho, K.L. An influenza a vaccine based on the extracellular domain of matrix 2 protein protects BALB/c mice against H1N1 and H3N2. Vaccines 2019, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Ho, K.L.; Gabrielsen, M.; Beh, P.L.; Kueh, C.L.; Thong, Q.X.; Streetley, J.; Tan, W.S.; Bhella, D. Structure of the Macrobrachium rosenbergii nodavirus: A new genus within the Nodaviridae? PLoS Biol. 2018, 16, e3000038. [Google Scholar] [CrossRef] [Green Version]
- Kueh, C.L.; Yong, C.Y.; Masoomi, D.S.; Bhassu, S.; Tan, S.G.; Tan, W.S. Virus-like particle of Macrobrachium rosenbergii nodavirus produced in Spodoptera frugiperda (Sf9) cells is distinctive from that produced in Escherichia coli. Biotechnol. Prog. 2017, 33, 549–557. [Google Scholar] [CrossRef]
- Ryan, B.J. Avoiding proteolysis protein chromatography. Methods Mol. Biol. 2011, 681, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, B.J.; Henehan, G.T. Overview of approaches to preventing and avoiding proteolysis during expression and purification of proteins. Curr. Protoc. Protein Sci. 2013, 71, 5–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozkov, A.; Enfors, S.O. Analysis and control of proteolysis of recombinant proteins in Escherichia coli. Adv. Biochem. Eng. Biotechnol. 2004, 89, 163–195. [Google Scholar] [CrossRef]
- Gasteiger, E.H.C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. Proteom. Protoc. Handb. 2005, 571–607. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Goh, Z.H.; Azmina, N.; Mohd, S.; Tan, S.G.; Bhassu, S.; Tan, W.S. RNA-binding region of Macrobrachium rosenbergii nodavirus capsid protein. J. Gen. Virol. 2014, 9, 1919–1928. [Google Scholar] [CrossRef] [Green Version]
- Hanapi, U.F.; Yong, C.Y.; Goh, Z.H.; Alitheen, N.B.; Yeap, S.K.; Tan, W.S. Tracking the virus-like particles of Macrobrachium rosenbergii nodavirus in insect cells. PeerJ 2017, 5, e2947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurizi, M.R. Proteases and protein degradation in Escherichia coli. Experientia 1992, 48, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Nilsson, L.; Kurland, C.G. Gratuitous overexpression of genes in Escherichia coli leads to growth inhibition and ribosome destruction. J. Bacteriol. 1995, 177, 1497–1504. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.I.; Baek, S.H.; Hong, Y.M.; Kang, M.S.; Ha, D.B.; Goldberg, A.L.; Chung, C.H. Purification and characterization of protease Ci, a cytoplasmic metalloendoprotease in Escherichia coli. J. Biol. Chem. 1995, 270, 29799–29805. [Google Scholar] [CrossRef] [Green Version]
- Laskowska, E.; Kuczyńska-Wiśnik, D.; Skórko-Glonek, J.; Taylor, A. Degradation by proteases Lon, Clp and HtrA, of Escherichia coli proteins aggregated in vivo by heat shock; HtrA protease action in vivo and in vitro. Mol. Microbiol. 1996, 22, 555–571. [Google Scholar] [CrossRef]
- Tomoyasu, T.; Gamer, J.; Bukau, B.; Kanemori, M.; Mori, H.; Rutman, A.J.; Oppenheim, A.B.; Yura, T.; Yamanaka, K.; Nikki, H. Escherichia coli FtsH is a membrane-bound, ATP-dependent protease which degrades the heat-shock transcription factor sigma 32. EMBO J. 1995, 14, 2551–2560. [Google Scholar] [CrossRef]
- James, G.T. Inactivation of the protease inhibitor phenylmethylsulfonyl fluoride in buffers. Anal. Biochem. 1978, 86, 574–579. [Google Scholar] [CrossRef]
- Powers, J.C.; Asgian, J.L.; Ekici, Ö.D.; James, K.E. Irreversible inhibitors of serine, cysteine, and threonine proteases. Chem. Rev. 2002, 102, 4639–4750. [Google Scholar] [CrossRef]
- Matsumoto, K.; Mizoue, K.; Kitamura, K.; Tse, W.C.; Huber, C.P.; Ishida, T. Structural basis of inhibition of cysteine proteases by E-64 and its derivatives. Biopolymers 1999, 51, 99–107. [Google Scholar] [CrossRef]
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999, 11, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Gotiesmanl, S.; Maurizi, M.R. Regulation by Proteolysis: Energy-dependent proteases and their targets. Microbiol. Rev. 1992, 56, 592–621. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chong, F.C.; Tan, W.S.; Biak, D.R.A.; Ling, T.C.; Tey, B.T. Modulation of protease activity to enhance the recovery of recombinant nucleocapsid protein of Nipah virus. Process Biochem. 2010, 45, 133–137. [Google Scholar] [CrossRef]
- Sigma. Trans-Epoxysuccinyl-l-Leucylamido-(4-Guanidino)Butane Synonyms: E 64 1; Proteinase Inhibitor E 64 1; N-[N-(L-3-Transcarboxyirane-2-Carbonyl)-L-Leucyl]-Agmatine 2 [Internet]. Available online: https://www.sigmaaldrich.com/content/dam/sigmaaldrich/docs/Sigma/Product_Information_Sheet/e3132pis.pdf (accessed on 2 September 2019).
- Gallo-Oller, G.; Ordoñez, R.; Dotor, J. A new background subtraction method for Western blot densitometry band quantification through image analysis software. J. Immunol. Methods 2018, 457, 1–5. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Values |
|---|---|
| Linearity range (µg/well) | 0.5–3.0 |
| Regression equation | y = 240,299 [Equation (1)] |
| Correlation co-efficient | 0.9908 |
| Inhibitor | Molecular Weight (Da) | Stock Concentration | Working Concentration | Type of Protease Inhibition | References |
|---|---|---|---|---|---|
| PMSF | 174.2 | 200 mM in isopropanol | 2 mM | Serine | [6] |
| AEBSF | 239.5 | 208.6 mM in H2O | 1 mM | Serine and Cysteine | [34,35] |
| E-64 | 357.41 | 1 mM in H2O | 10 µM | Cysteine | [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvaraj, B.A.; Mariatulqabtiah, A.R.; Ho, K.L.; Ng, C.L.; Yong, C.Y.; Tan, W.S. Regulation of Proteolytic Activity to Improve the Recovery of Macrobrachium rosenbergii Nodavirus Capsid Protein. Int. J. Mol. Sci. 2021, 22, 8725. https://doi.org/10.3390/ijms22168725
Selvaraj BA, Mariatulqabtiah AR, Ho KL, Ng CL, Yong CY, Tan WS. Regulation of Proteolytic Activity to Improve the Recovery of Macrobrachium rosenbergii Nodavirus Capsid Protein. International Journal of Molecular Sciences. 2021; 22(16):8725. https://doi.org/10.3390/ijms22168725
Chicago/Turabian StyleSelvaraj, Bethilda Anne, Abdul Razak Mariatulqabtiah, Kok Lian Ho, Chyan Leong Ng, Chean Yeah Yong, and Wen Siang Tan. 2021. "Regulation of Proteolytic Activity to Improve the Recovery of Macrobrachium rosenbergii Nodavirus Capsid Protein" International Journal of Molecular Sciences 22, no. 16: 8725. https://doi.org/10.3390/ijms22168725