
Blue Light Improves Photosynthetic Performance during Healing and Acclimatization of Grafted Watermelon Seedlings

 ,
, 
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Survival, Growth and Morphology of Grafted Seedlings
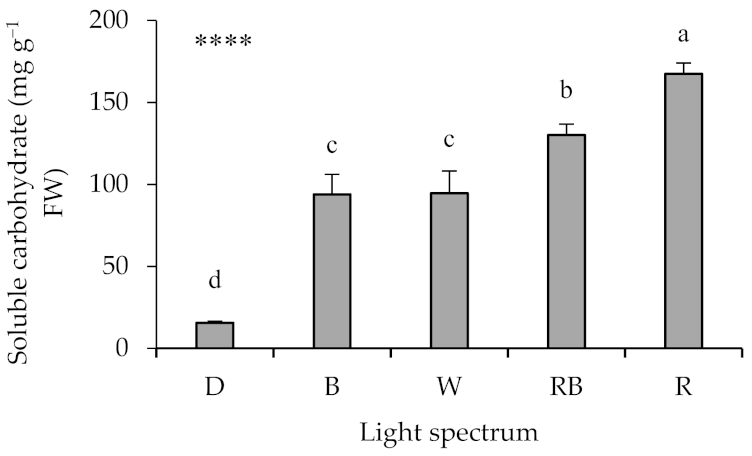
2.2. Leaf Soluble Carbohydrates
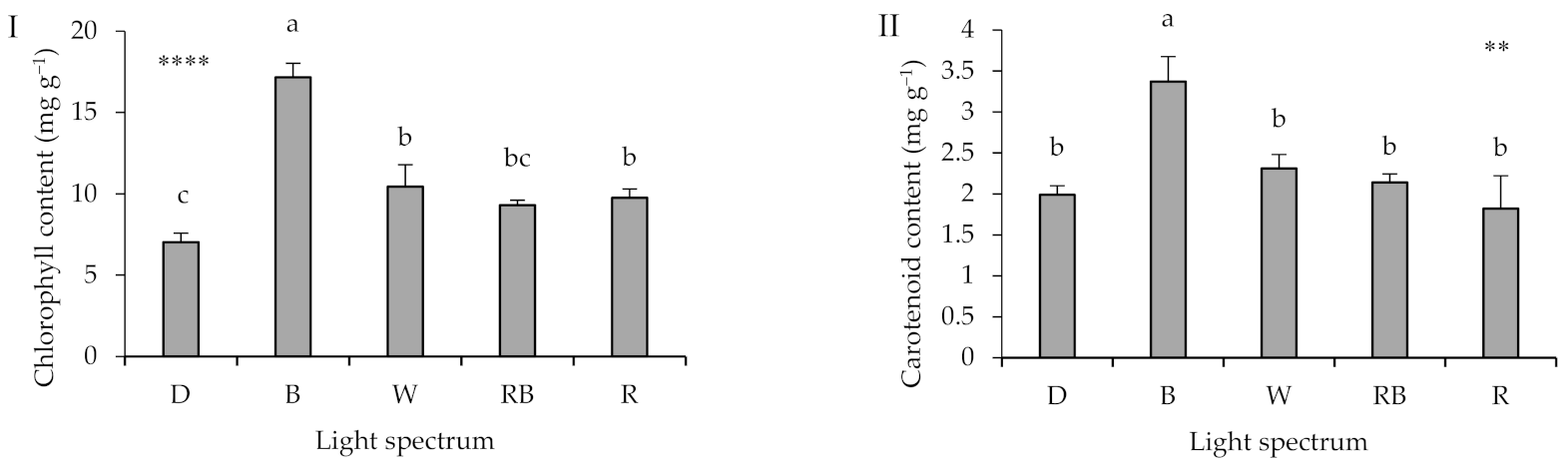
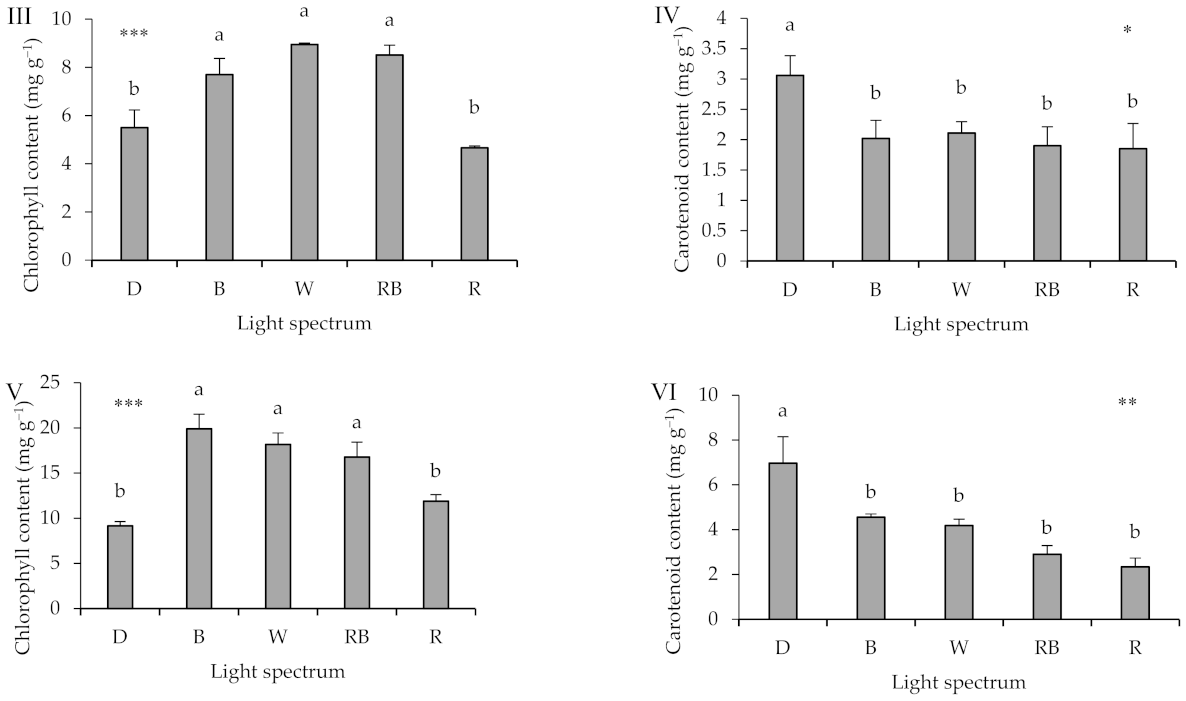
2.3. Leaf Photosynthetic Pigments
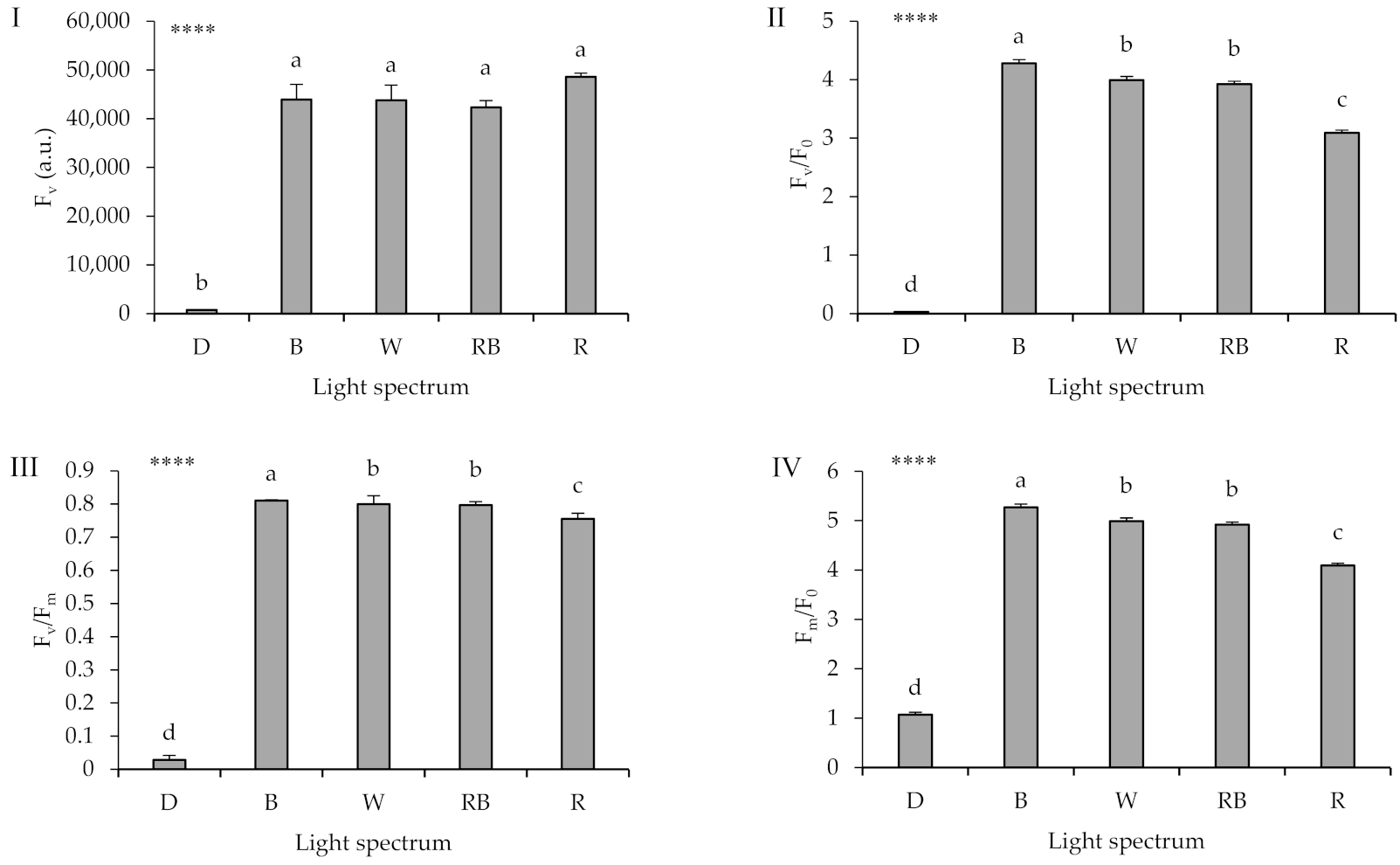
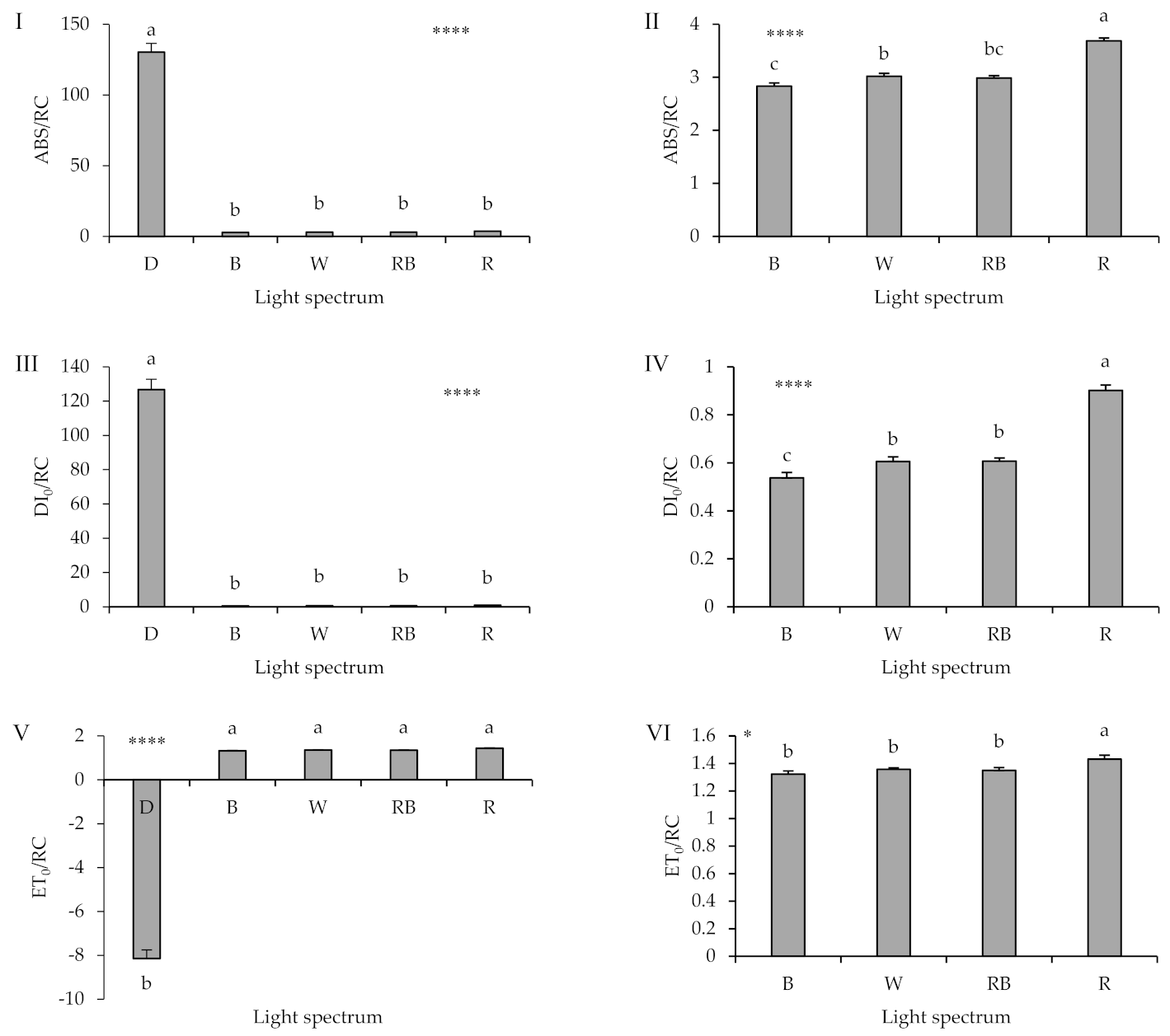
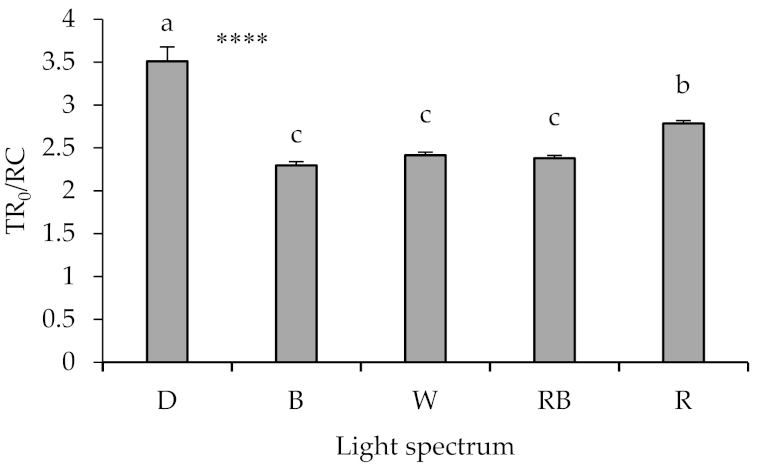
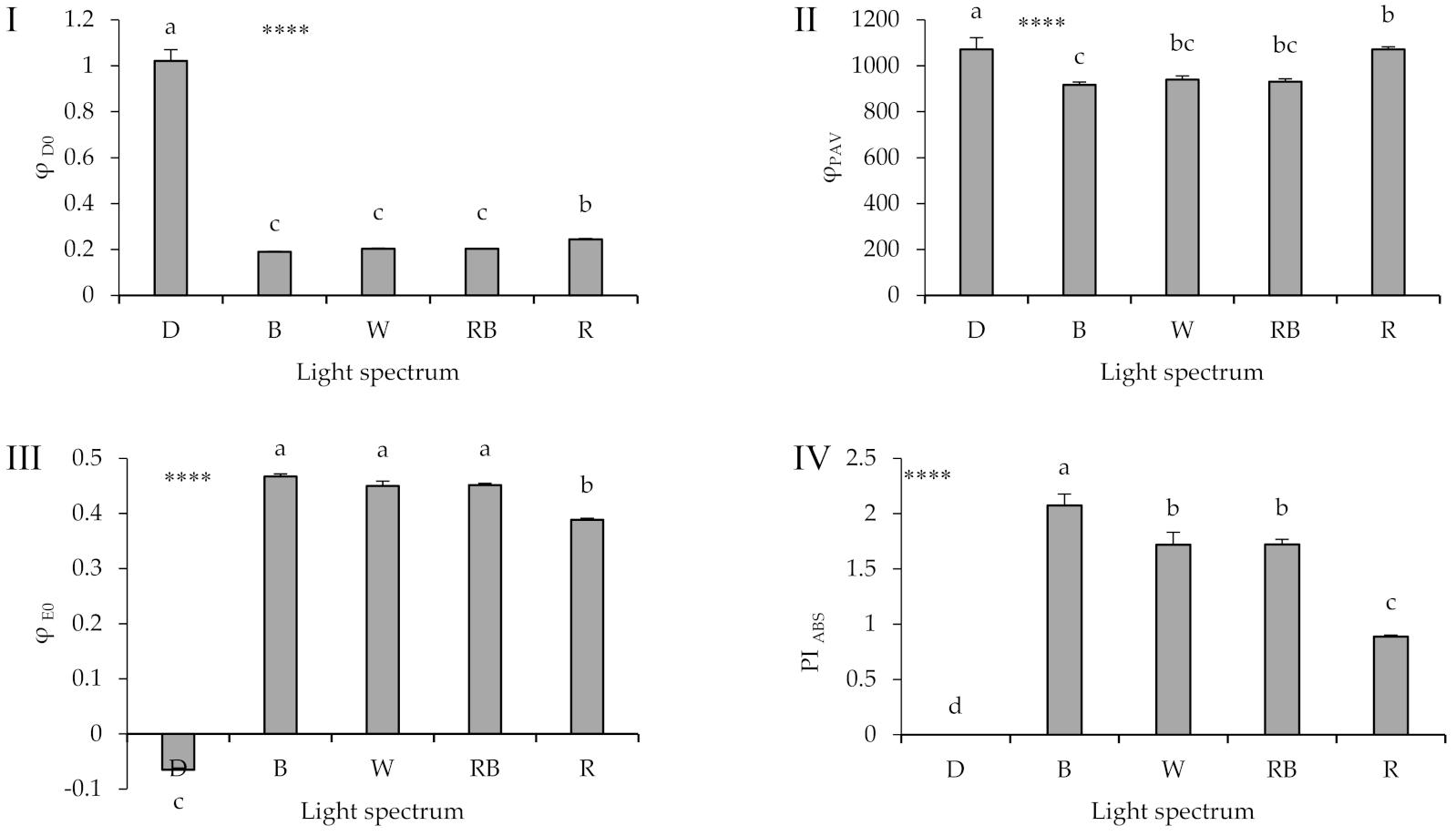
2.4. Chlorophyll Fluorescence Imaging
2.5. Polyphasic Chlorophyll Fluorescence Transient (OJIP) Evaluation
3. Discussion
3.1. Light during Healing and Acclimatization Improves Survival Ratio of the Grafted Seedlings
3.2. Morphology and Growth of the Grafted Seedlings Depends on the Light Spectrum during Healing and Acclimatization
3.3. Light Spectra during Healing and Acclimatization Affects Pigment Content in Different Parts of the Seedlings
3.4. Photosynthetic Functionality Is Affected by the Presence of Light and Its Spectrum during Healing and Acclimatization
3.5. High Proportion of B (i.e., B, W, RB) in the Overall Spectrum Up-Regulates Electron Transport, Leading to Improved Photosynthetic Functionality
4. Materials and Methods
4.1. Plant Material and Exposure Conditions
4.2. Survival Ratio, Plant Biomass and Leaf Morphology
4.3. Determination of Soluble Carbohydrates
4.4. Photosynthetic Pigments
4.5. Chlorophyll Fluorescence Imaging
4.6. Polyphasic Chlorophyll Fluorescence Transient (OJIP) Evaluation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basic Parameters | ||
|---|---|---|
| F0 | Minimum fluorescence, when all PSII reaction centers (RCs) are open (O-step of OJIP transient) | F50µs |
| FJ | Fluorescence intensity at the J-step (2 ms) of OJIP | F2 ms |
| FI | Fluorescence intensity at the I-step (30 ms) of OJIP | F30ms |
| Fluorescence Parameters | ||
| Fm | Maximum fluorescence, when all PSII RCs are closed (P-step of OJIP transient) | F1s = Fp |
| Fv | Variable fluorescence of the dark-adapted leaf | Fm − F0 |
| VJ | Relative variable fluorescence at time 2 ms (J-step) after start of actinic light pulse | (FJ − F0)/(Fm − F0) |
| VI | Relative variable fluorescence at time 30 ms (I-step) after start of actinic light pulse | (F30ms − F0)/(Fm − F0) |
| Fv/Fm | Maximal quantum yield of PSII photochemistry | 1 − (F0/Fm) = (Fm − F0)/Fm = φP0 = TR0/ABS |
| Quantum Yields and Efficiencies/Probabilities | ||
| φE0 | The quantum yield of electron transport | [1 − (F0/Fm)](1 − VJ) = ET0/ABS |
| φD0 | Quantum yield of energy dissipation | F0/Fm |
| φPAV | Average quantum yield for primary photochemistry | φP0 (1 − VJ) = φP0 (SM/tFM) |
| Specific Energy Fluxes (Per QA Reducing PSII RC) | ||
| ABS/RC | The specific energy fluxes per RC for energy absorption | M0 (1/VJ)(1/φP0) |
| TR0/RC | Trapped energy flux (leading to QA reduction) per RC | M0 (1/VJ) |
| ET0/RC | Electron transport flux (further than QA−) per RC | M0 (1/VJ)(1 − VJ) |
| DI0/RC | Dissipated energy flux | (ABS/RC) − (TR0 /RC) |
| Performance Indexes (Products of Terms Expressing Partial Potentials at Steps of Energy Bifurcations) | ||
| PIABS | Performance index for the photochemical activity | [(γRC/1 − γRC) (φP0 /1 − φP0) (ψE0 /1 − ψE0)] |
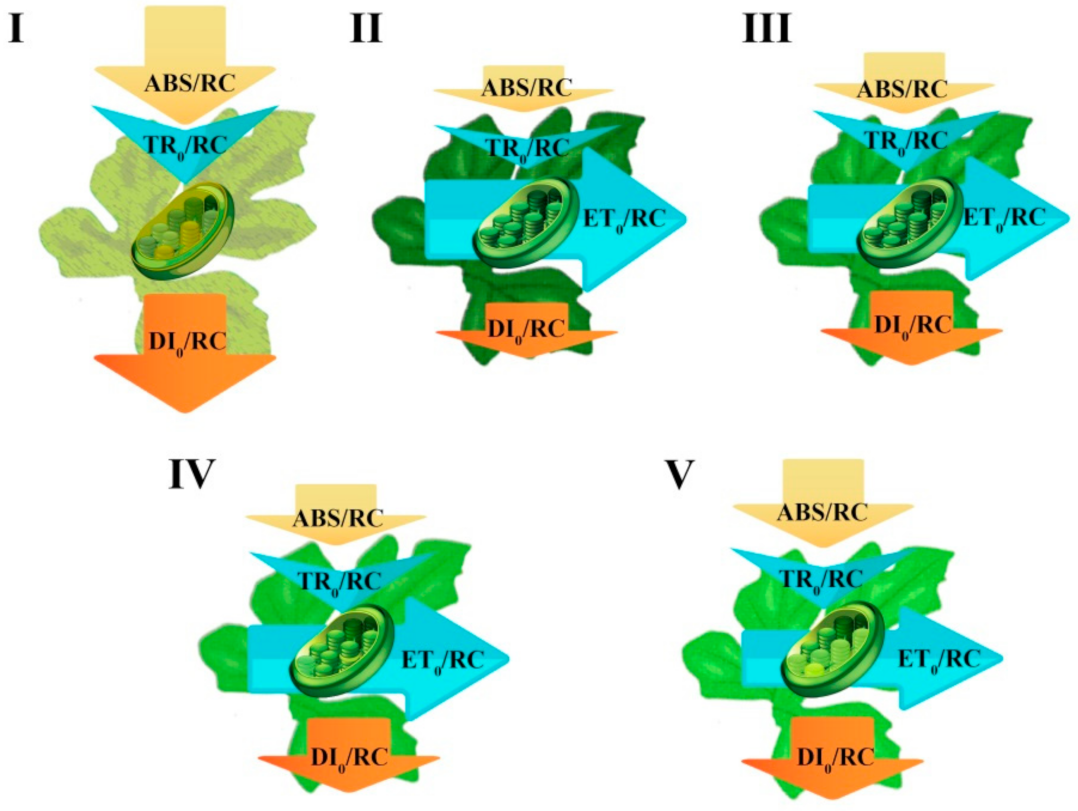
4.7. Energy Pipeline Model
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| B | blue |
| D | darkness |
| DQI | Dickson’s quality index |
| LMR | leaf mass ratio |
| O–J–I–P | polyphasic chlorophyll fluorescence induction curve |
| PPFD | photosynthetic photon flux density |
| PSII | Photosystem II |
| R | red |
| RB | red and blue |
| RC | reaction center |
| SLA | specific leaf area |
| W | white |
References
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Koukounaras, A.; Siomos, A.; Menexes, G.; Dangitsis, C.; Kintzonidis, D. Assessing quantitative criteria for characterization of quality categories for grafted watermelon seedlings. Horticulturae 2019, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Koukounaras, A.; Siomos, A.S.; Fotelli, M.N.; Kintzonidis, D. Bichromatic red and blue LEDs during healing enhance the vegetative growth and quality of grafted watermelon seedlings. Sci. Hortic. 2020, 261, 109000. [Google Scholar] [CrossRef]
- Johnson, S.J.; Miles, C.A. Effect of healing chamber design on the survival of grafted eggplant, tomato, and watermelon. Horttechnology 2011, 21, 752–758. [Google Scholar] [CrossRef] [Green Version]
- Vu, N.T.; Kim, Y.S.; Kang, H.M.; Kim, I.S. Influence of short-term irradiation during pre- and post-grafting period on the graft-take ratio and quality of tomato seedlings. Hortic. Environ. Biotechnol. 2014, 55, 27–35. [Google Scholar] [CrossRef]
- Jang, Y.; Mun, B.; Seo, T.; Lee, J.; Oh, S.; Chun, C. Effects of light quality and intensity on the carbon dioxide exchange rate, growth, and morphogenesis of grafted pepper transplants during healing and acclimatization. Korean J. Hortic. Sci. Technol. 2013, 31, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Colla, G.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Melnyk, C.W. Plant grafting: Insights into tissue regeneration. Regeneration 2017, 4, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, C.W.; Schuster, C.; Leyser, O.; Meyerowitz, E.M. A developmental framework for graft formation and vascular reconnection in arabidopsis thaliana. Curr. Biol. 2015, 25, 1306–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousef, A.F.; Ali, M.M.; Rizwan, H.M.; Ahmed, M.A.A.; Ali, W.M.; Kalaji, H.M.; Elsheery, N.; Wróbel, J.; Xu, Y.; Chen, F. Effects of light spectrum on morphophysiological traits of grafted tomato seedlings. PLoS ONE 2021, 16, e0250210. [Google Scholar] [CrossRef]
- Chen, X.L.; Wang, L.C.; Li, T.; Yang, Q.C.; Guo, W.Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.Y.; Wang, L.T.; Ma, J.H.; Ma, E.D.; Li, J.Y.; Gong, M. Effects of light quality on growth and development, photosynthetic characteristics and content of carbohydrates in tobacco (Nicotiana tabacum L.) plants. Photosynthetica 2017, 55, 467–477. [Google Scholar] [CrossRef]
- Dabirian, S.; Miles, C.A. Increasing survival of splice-grafted watermelon seedlings using a sucrose application. HortScience 2017, 52, 579–583. [Google Scholar] [CrossRef]
- Dabirian, S.; Miles, C.A. Antitranspirant application increases grafting success of watermelon. Horttechnology 2017, 27, 494–501. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Zagorchev, L.; Atanasova, A.; Albanova, I.; Traianova, A.; Mladenov, P.; Kouzmanova, M.; Goltsev, V.; Kalaji, H.; Teofanova, D. Functional Characterization of the Photosynthetic Machinery in Smicronix Galls on the Parasitic Plant Cuscuta campestris by JIP-Test. Cells 2021, 10, 1399. [Google Scholar] [CrossRef]
- Chen, Y.; Fanourakis, D.; Tsaniklidis, G.; Aliniaeifard, S.; Yang, Q.; Li, T. Low UVA intensity during cultivation improves the lettuce shelf-life, an effect that is not sustained at higher intensity. Postharvest Biol. Technol. 2020, 172, 111376. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and physiological properties of indoor cultivated lettuce in response to additional far-red light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Nobuoka, T.; Nishimoto, T.; Toi, K. Wind and light promote graft-take and growth of grafted tomato seedlings. J. Jpn. Soc. Hortic. Sci. 2005, 74, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Hong, Y.; Zhang, X. Effect of light quality on calluses induction and differentiation of Capsicum annuum. J. Hunan Agric. Univ. 2009, 35, 615–617. [Google Scholar]
- Afshari, R.T.; Angoshtari, R.; Kalantari, S. Effects of light and different plant growth regulators on induction of callus growth in rapeseed (’Brassica napus L.’) genotypes. Plant Omics J. 2011, 4, 60–67. [Google Scholar]
- Lee, K.M.; Lim, C.S.; Muneer, S.; Jeong, B.R. Functional vascular connections and light quality effects on tomato grafted unions. Sci. Hortic. 2016, 201, 306–317. [Google Scholar] [CrossRef]
- Zavala, J.A.; Ravetta, D.A. Allocation of photoassimilates to biomass, resin and carbohydrates in Grindelia chiloensis as affected by light intensity. Field Crop. Res. 2001, 69, 143–149. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef]
- Fan, J.; Yu, L.; Xu, C. A central role for triacylglycerol in membrane lipid breakdown, fatty acid β -oxidation, and plant survival under extended darkness. Plant Physiol. 2017, 174, 1517–1530. [Google Scholar] [CrossRef] [Green Version]
- Kunz, H.-H.; Scharnewski, M.; Feussner, K.; Feussner, I.; Flügge, U.-I.; Fulda, M.; Gierth, M. The ABC transporter PXA1 and peroxisomal β-oxidation are vital for metabolism in mature leaves of arabidopsis during extended darkness. Plant Cell 2009, 21, 2733–2749. [Google Scholar] [CrossRef] [Green Version]
- Usadel, B.; Bläsing, O.E.; Gibon, Y.; Poree, F.; Höhne, M.; Günter, M.; Trethewey, R.; Kamlage, B.; Poorter, H.; Stitt, M. Multilevel genomic analysis of the response of transcripts, enzyme activities and metabolites in Arabidopsis rosettes to a progressive decrease of temperature in the non-freezing range. Plant Cell Environ. 2008, 31, 518–547. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Monte, E.; Oka, Y.; Liu, T.; Carle, C.; Castillon, A.; Huq, E.; Quail, P.H. Multiple phytochrome-interacting bHLH transcription factors repress premature seedling photomorphogenesis in darkness. Curr. Biol. 2008, 18, 1815–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Johnson, R.E.; Kong, Y.; Zheng, Y. Elongation growth mediated by blue light varies with light intensities and plant species: A comparison with red light in arugula and mustard seedlings. Environ. Exp. Bot. 2020, 169, 103898. [Google Scholar] [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y. Pure blue light effects on growth and morphology are slightly changed by adding low-level UVA or far-red light: A comparison with red light in four microgreen species. Environ. Exp. Bot. 2019, 157, 58–68. [Google Scholar] [CrossRef]
- Seif, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.Z.; Shomali, A.; Fanourakis, D.; Li, T.; Woltering, E. Monochromatic red light during plant growth decreases the size and improves the functionality of stomata in chrysanthemum. Funct. Plant Biol. 2021, 48, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, N.; Ajima, C.; Yukawa, T.; Olsen, J.E. Antagonistic action of blue and red light on shoot elongation in petunia depends on gibberellin, but the effects on flowering are not generally linked to gibberellin. Environ. Exp. Bot. 2016, 121, 102–111. [Google Scholar] [CrossRef]
- Savvides, A.; Fanourakis, D.; van Ieperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2012, 63, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanourakis, D.; Aliniaeifard, S.; Sellin, A.; Giday, H.; Körner, O.; Rezaei Nejad, A.; Delis, C.; Bouranis, D.; Koubouris, G.; Kambourakis, E.; et al. Stomatal behavior following mid- or long-term exposure to high relative air humidity: A review. Plant Physiol. Biochem. 2020, 153, 92–105. [Google Scholar] [CrossRef]
- Fanourakis, D.; Bouranis, D.; Tsaniklidis, G.; Rezaei Nejad, A.; Ottosen, C.O.; Woltering, E.J. Genotypic and phenotypic differences in fresh weight partitioning of cut rose stems: Implications for water loss. Acta Physiol. Plant. 2020, 42, 48. [Google Scholar] [CrossRef]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Zare Mehrjerdi, M.; Li, T.; Lastochkina, O. Growth and photosynthetic performance of Calendula officinalis under monochromatic red light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Trouwborst, G.; Hogewoning, S.W.; van Kooten, O.; Harbinson, J.; van Ieperen, W. Plasticity of photosynthesis after the ‘red light syndrome’ in cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [Green Version]
- Haga, K.; Takano, M.; Neumann, R.; Iino, M. The rice COLEOPTILE PHOTOTROPISM1 gene encoding an ortholog of arabidopsis NPH3 is required for phototropism of coleoptiles and lateral translocation of auxin. Plant Cell 2005, 17, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Romero-Puertas, M.C. Leaf epinasty and auxin: A biochemical and molecular overview. Plant Sci. 2016, 253, 187–193. [Google Scholar] [CrossRef]
- Trouwborst, G.; Oosterkamp, J.; Hogewoning, S.W.; Harbinson, J.; van Ieperen, W. The responses of light interception, photosynthesis and fruit yield of cucumber to LED-lighting within the canopy. Physiol. Plant 2010, 138, 289–300. [Google Scholar] [CrossRef]
- Krapp, A.; Quick, W.P.; Stitt, M. Ribulose-1,5-bisphosphate carboxylase-oxygenase, other Calvin-cycle enzymes, and chlorophyll decrease when glucose is supplied to mature spinach leaves via the transpiration stream. Planta 1991, 186, 58–69. [Google Scholar] [CrossRef]
- Nafziger, E.D.; Koller, H.R. Influence of leaf starch concentration on CO2 assimilation in soybean. Plant Physiol. 1976, 57, 560–563. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Kurata, K. Effects of blue-light photon flux density on nitrogen and carbohydrate content and growth of spinach. Acta Hortic. 2008, 801, 1393–1398. [Google Scholar] [CrossRef]
- Terfa, M.T.; Solhaug, K.A.; Gislerød, H.R.; Olsen, J.E.; Torre, S. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa × hybrida but does not affect time to flower opening. Physiol. Plant 2013, 148, 146–159. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, A.; Zare Mehrjerdi, M.; Aliniaeifard, S.; Seif, M. Photosynthetic and growth responses of green and purple basil plants under different spectral compositions. Physiol. Mol. Biol. Plants 2019, 25, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Jiang, W.; Ding, M.; Lin, Y.; Huang, D. Light affects the chloroplast ultrastructure and post-storage photosynthetic performance of watermelon (Citrullus lanatus) plug seedlings. PLoS ONE 2014, 9, e111165. [Google Scholar] [CrossRef] [Green Version]
- Falqueto, A.R.; da Silva Júnior, R.A.; Gomes, M.T.G.; Martins, J.P.R.; Silva, D.M.; Partelli, F.L. Effects of drought stress on chlorophyll a fluorescence in two rubber tree clones. Sci. Hortic. 2017, 224, 238–243. [Google Scholar] [CrossRef]
- Gasulla, F.; Casano, L.; Guéra, A. Chlororespiration induces non–photochemical quenching of chlorophyll fluorescence during darkness in lichen chlorobionts. Physiol. Plant 2019, 166, 538–552. [Google Scholar] [CrossRef] [PubMed]
- Podmaniczki, A.; Nagy, V.; Vidal-Meireles, A.; Tóth, D.; Patai, R.; Kovács, L.; Tóth, S.Z. Ascorbate inactivates the oxygen-evolving complex in prolonged darkness. Physiol. Plant 2021, 171, 232–245. [Google Scholar] [CrossRef]
- Roosta, H.R.; Estaji, A.; Niknam, F. Effect of iron, zinc and manganese shortage-induced change on photosynthetic pigments, some osmoregulators and chlorophyll fluorescence parameters in lettuce. Photosynthetica 2018, 56, 606–615. [Google Scholar] [CrossRef]
- Xue, X.; Wang, Q.; Qu, Y.; Wu, H.; Dong, F.; Cao, H.; Wang, H.-L.; Xiao, J.; Shen, Y.; Wan, Y. Development of the photosynthetic apparatus of Cunninghamia lanceolata in light and darkness. New Phytol. 2017, 213, 300–313. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C. Spectral effects of artificial light on plant physiology and secondary metabolism: A Review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. HortScience 2001, 36, 380–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Van Labeke, M.C. Chrysanthemum morphology, photosynthetic efficiency and antioxidant capacity are differentially modified by light quality. J. Plant Physiol. 2017, 213, 66–74. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, T.; Fang, S.; Zhou, M.; Qin, J. Responses of morphology, gas exchange, photochemical activity of photosystem II, and antioxidant balance in Cyclocarya paliurus to light spectra. Front. Plant Sci. 2018, 9, 1704. [Google Scholar] [CrossRef] [Green Version]
- Pham, M.D.; Hwang, H.; Park, S.W.; Cui, M.; Lee, H.; Chun, C. Leaf chlorosis, epinasty, carbohydrate contents and growth of tomato show different responses to the red/blue wavelength ratio under continuous light. Plant Physiol. Biochem. 2019, 141, 477–486. [Google Scholar] [CrossRef]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell. Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Jeannette, E.; Reyss, A.; Grégory, N.; Gantet, P.; Prioul, J.L. Carbohydrate metabolism in a heat-girdled maize source leaf. Plant Cell Environ. 2000, 23, 61–69. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xin, G.; Liu, C.; Shi, Q.; Yang, F.; Wei, M. Effects of red and blue light on leaf anatomy, CO2 assimilation and the photosynthetic electron transport capacity of sweet pepper (Capsicum annuum L.) seedlings. BMC Plant Biol. 2020, 20, 318. [Google Scholar]
- Hassanvand, F.; Rezaei Nejad, A.; Fanourakis, D. Morphological and physiological components mediating the silicon-induced enhancement of geranium essential oil yield under saline conditions. Ind. Crop. Prod. 2019, 134, 19–25. [Google Scholar] [CrossRef]
- Żuk-Gołaszewska, K.; Upadhyaya, M.K.; Gołaszewski, J. The effect of UV-B radiation on plant growth and development. Plant Soil Environ. 2003, 49, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Fanourakis, D.; Briese, C.; Max, J.F.J.; Kleinen, S.; Putz, A.; Fiorani, F.; Ulbrich, A.; Schurr, U. Rapid determination of leaf area and plant height by using light curtain arrays in four species with contrasting shoot architecture. Plant Methods 2014, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Bergsträsser, S.; Fanourakis, D.; Schmittgen, S.; Cendrero-Mateo, M.P.; Jansen, M.; Scharr, H.; Rascher, U. HyperART: Non-invasive quantification of leaf traits using hyperspectral absorption-reflectance-transmittance imaging. Plant Methods 2015, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Kalhor, M.S.; Aliniaeifard, S.; Seif, M.; Asayesh, E.J.; Bernard, F.; Hassani, B.; Li, T. Enhanced salt tolerance and photosynthetic performance: Implication of ɤ-amino butyric acid application in salt-exposed lettuce (Lactuca sativa L.) plants. Plant Physiol. Biochem. 2018, 130, 157–172. [Google Scholar] [CrossRef]
- Sørensen, H.K.; Fanourakis, D.; Tsaniklidis, G.; Bouranis, D.; Rezaei Nejad, A.; Ottosen, C.O. Using artificial lighting based on electricity price without a negative impact on growth, visual quality or stomatal closing response in Passiflora. Sci. Hortic. 2020, 267, 109354. [Google Scholar] [CrossRef]
- Kalaji, M.H.; Goltsev, V.N.; Żuk-Gołaszewska, K.; Zivcak, M.; Brestic, M. Chlorophyll Fluorescence: Understanding Crop Performance—Basics and Applications; CRC Press LLC: Boca Raton, FL, USA, 2017; p. 222. ISBN 1498764509. [Google Scholar]
- Küpper, H.; Benedikty, Z.; Morina, F.; Andresen, E.; Mishra, A.; Trtílek, M. Analysis of OJIP chlorophyll fluorescence kinetics and QA reoxidation kinetics by direct fast imaging. Plant Physiol. 2019, 179, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi-Nezhad, S.M. How glycine betaine induces tolerance of cucumber plants to salinity stress. Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Gorbe, E.; Calatayud, A. Applications of chlorophyll fluorescence imaging technique in horticultural research: A review. Sci. Hortic. 2012, 138, 24–35. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Kromdijk, J. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10, ply052. [Google Scholar] [CrossRef] [PubMed]










| Light Regime | Survival Ratio (%) | Rootstock Length (cm) | Scion Length (cm) | Scion Stem Diameter (cm) | Leaf Number (plant−1) | Leaf Area (cm2 leaf−1) | Leaf Area (cm2 plant−1) | Plant Dry Weight (g) | Shoot Dry Weight (g) | Specific Leaf Area (SLA; cm2 g−1) | Leaf Mass Ratio (LMR; g g−1) | Root Length (cm) | Root Dry Weight (g) | Shoot to Root Ratio (g/g) | Dickson’s Quality Index (DQI) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D | 26.3 b | 7.51 | 2.45 b | 0.31 | 2.3 c | 5.01 d | 26.4 d | 0.174 b | 0.165 b | 247 c | 0.38 a,b | 4.14 b | 0.009 c | 19.17 a | 0.006 b |
| B | 100 a | 5.87 | 4.15 a | 0.33 | 5.0 b | 8.24 b | 65.9 b | 0.290 a | 0.265 a | 417 b | 0.35 c | 6.38 a | 0.025 a | 10.53 b | 0.015 a |
| W | 100 a | 6.05 | 4.10 a | 0.38 | 5.0 b | 10.11 a | 80.8 a | 0.251 a | 0.230 a | 540 a | 0.37 b | 6.33 a | 0.021 a | 11.02 b | 0.013 a |
| RB | 100 a | 6.27 | 2.42 b | 0.40 | 6.0 a | 8.15 b | 73.4 a,b | 0.282 a | 0.270 a | 411 b | 0.39 a | 4.24 b | 0.012 b | 23.17 a | 0.013 a |
| R | 100 a | 6.38 | 2.25 b | 0.38 | 5.3 b | 6.28 c | 52.2 c | 0.265 a | 0.256 a | 376 b | 0.34 c | 6.72 a | 0.024 a | 10.16 b | 0.016 a |
| p | <0.0001 | 0.0963 | 0.0075 | 0.23 | <0.0001 | <0.0001 | <0.0001 | 0.0003 | <0.0001 | <0.0001 | <0.0001 | 0.0065 | <0.0001 | <0.0001 | 0.0009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moosavi-Nezhad, M.; Salehi, R.; Aliniaeifard, S.; Tsaniklidis, G.; Woltering, E.J.; Fanourakis, D.; Żuk-Gołaszewska, K.; Kalaji, H.M. Blue Light Improves Photosynthetic Performance during Healing and Acclimatization of Grafted Watermelon Seedlings. Int. J. Mol. Sci. 2021, 22, 8043. https://doi.org/10.3390/ijms22158043
Moosavi-Nezhad M, Salehi R, Aliniaeifard S, Tsaniklidis G, Woltering EJ, Fanourakis D, Żuk-Gołaszewska K, Kalaji HM. Blue Light Improves Photosynthetic Performance during Healing and Acclimatization of Grafted Watermelon Seedlings. International Journal of Molecular Sciences. 2021; 22(15):8043. https://doi.org/10.3390/ijms22158043
Chicago/Turabian StyleMoosavi-Nezhad, Moein, Reza Salehi, Sasan Aliniaeifard, Georgios Tsaniklidis, Ernst J. Woltering, Dimitrios Fanourakis, Krystyna Żuk-Gołaszewska, and Hazem M. Kalaji. 2021. "Blue Light Improves Photosynthetic Performance during Healing and Acclimatization of Grafted Watermelon Seedlings" International Journal of Molecular Sciences 22, no. 15: 8043. https://doi.org/10.3390/ijms22158043