
Role of Thylakoid Protein Phosphorylation in Energy-Dependent Quenching of Chlorophyll Fluorescence in Rice Plants


Abstract
:1. Introduction
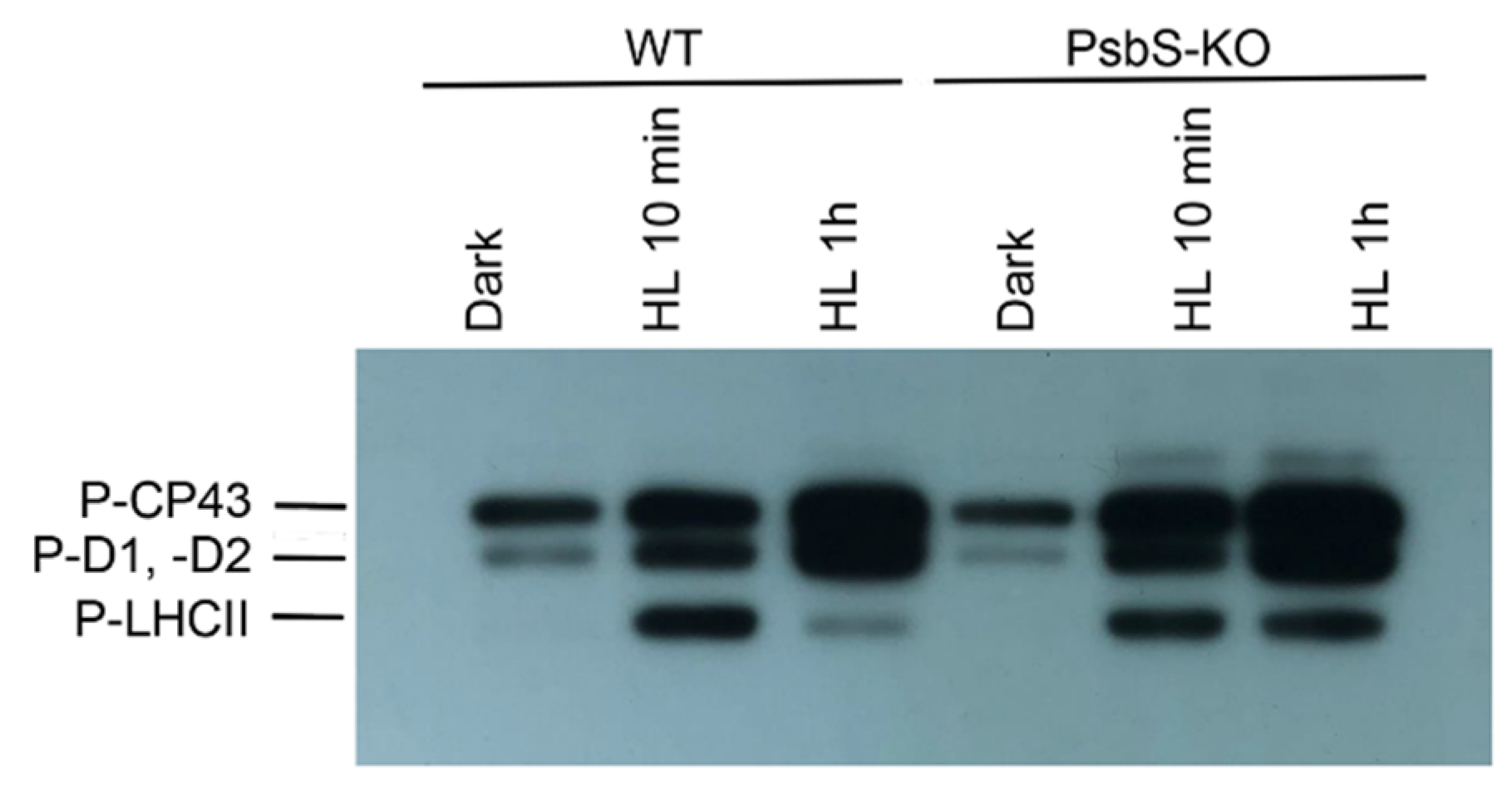
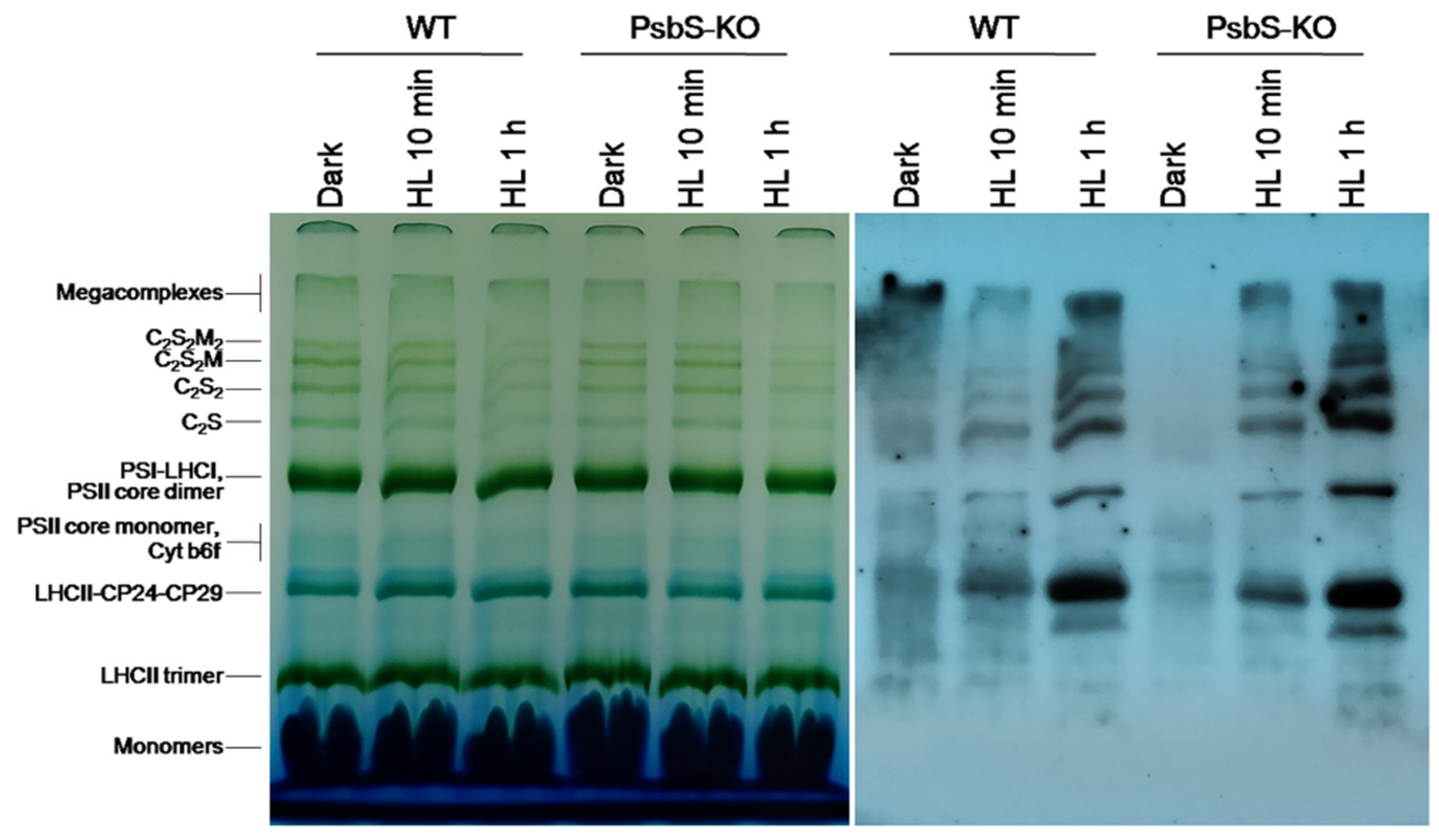
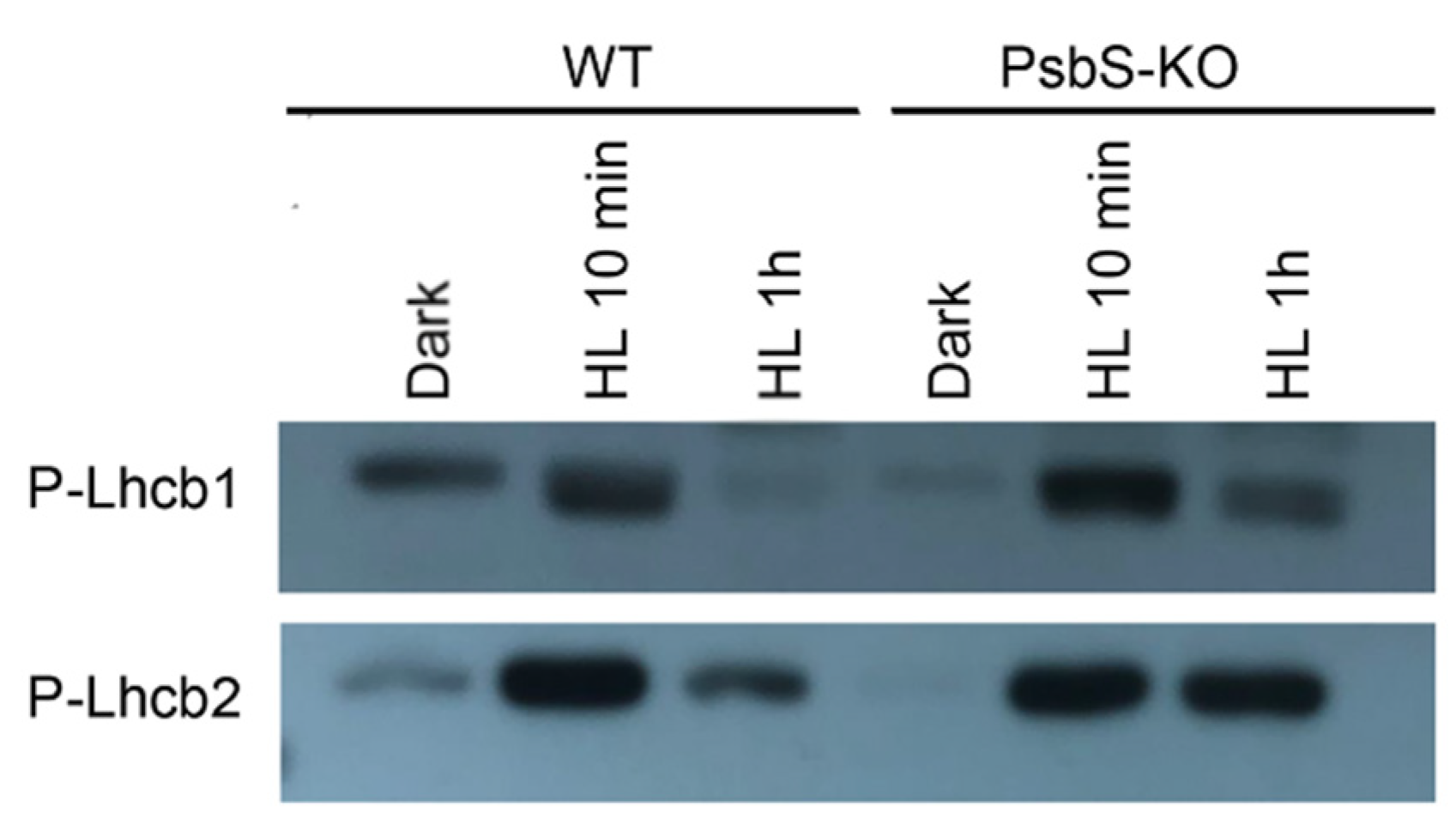
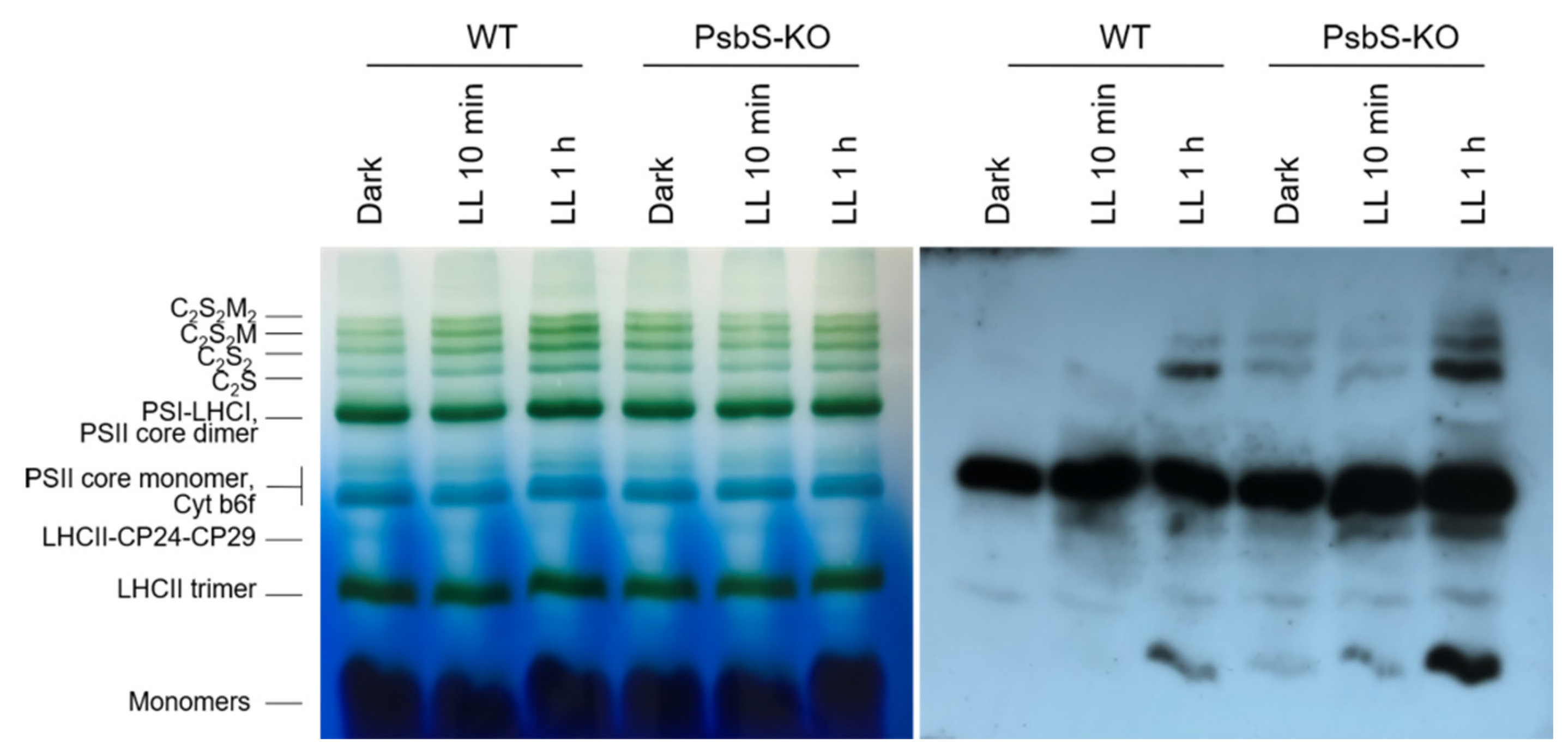
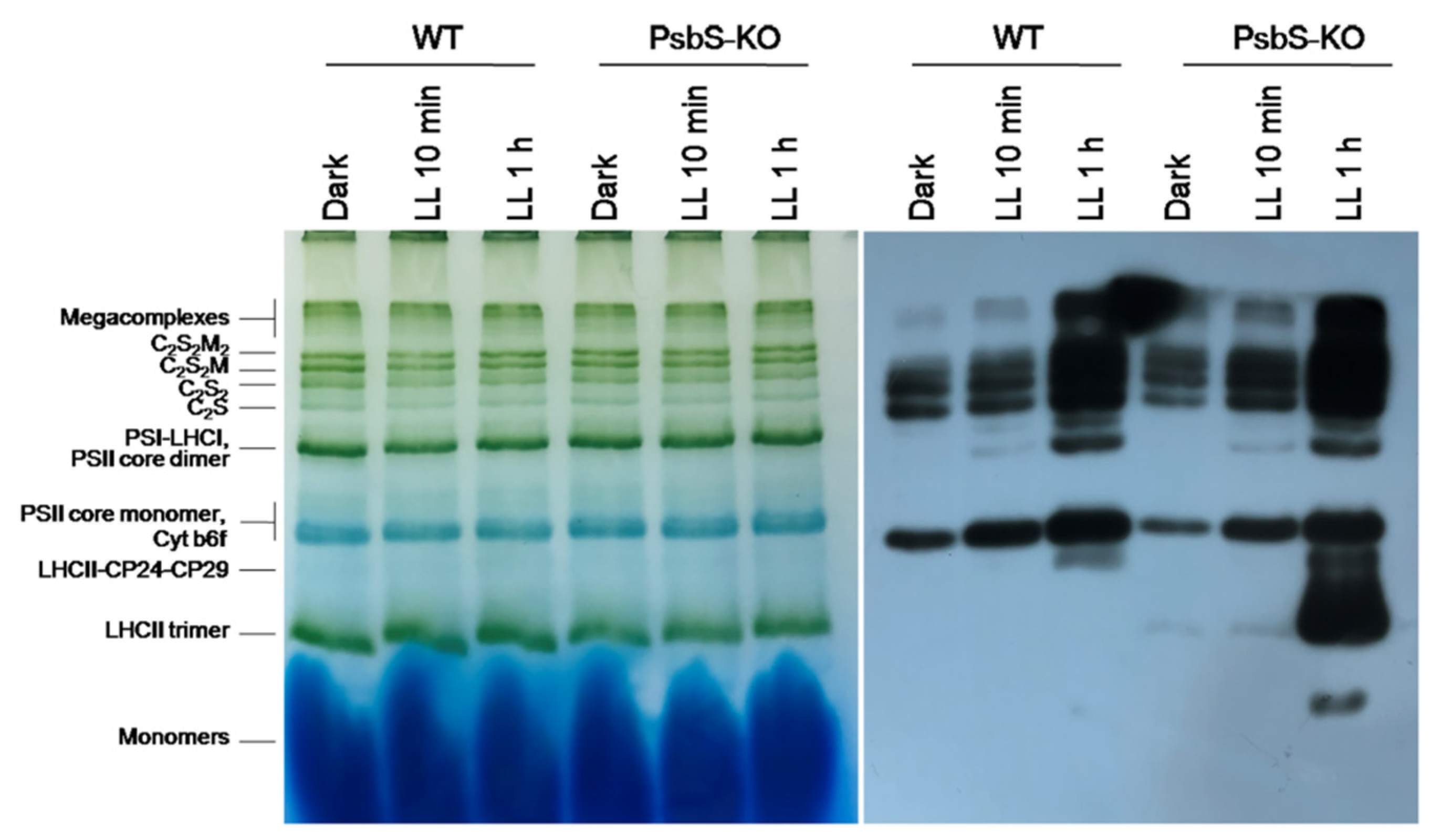
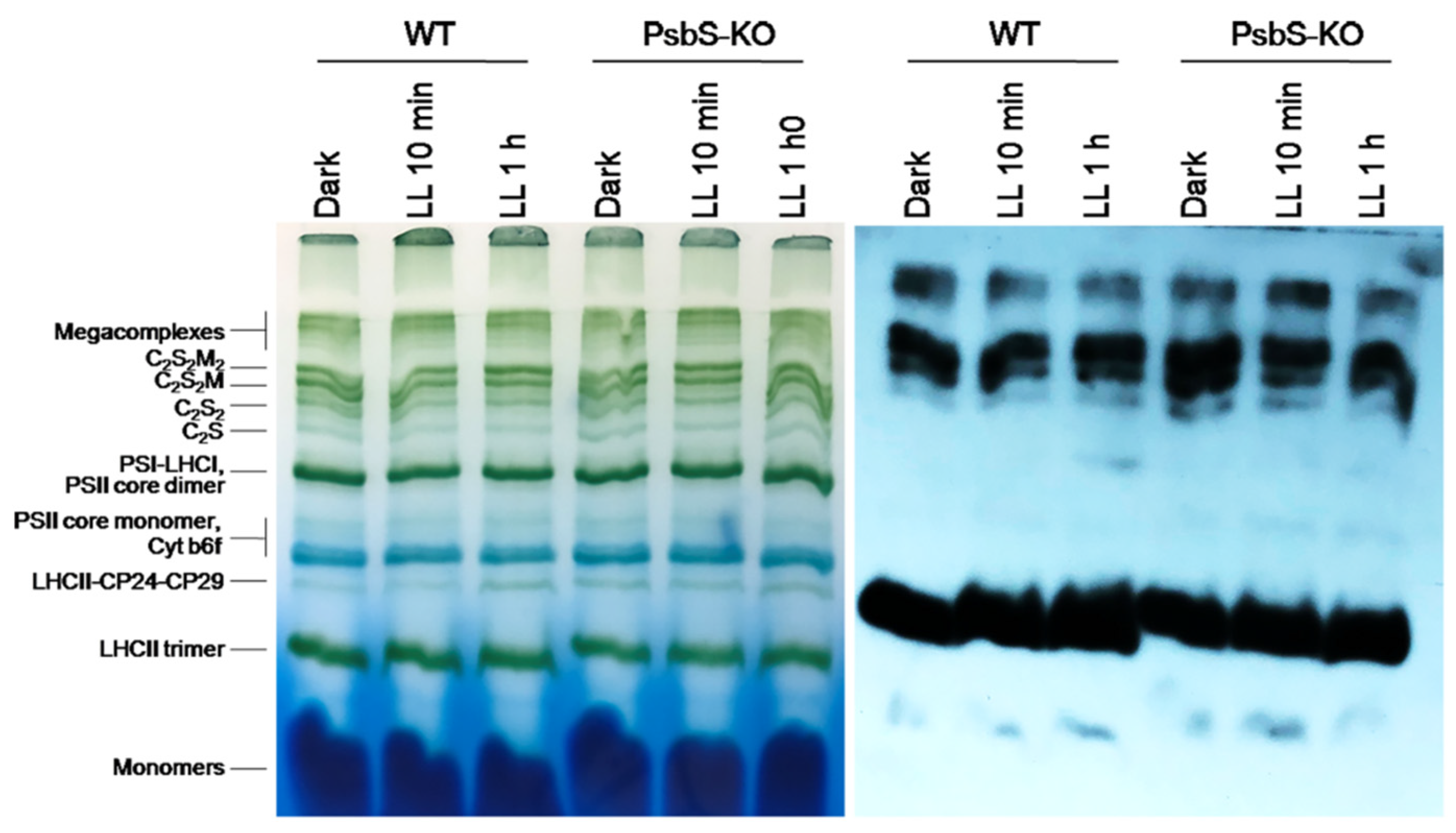
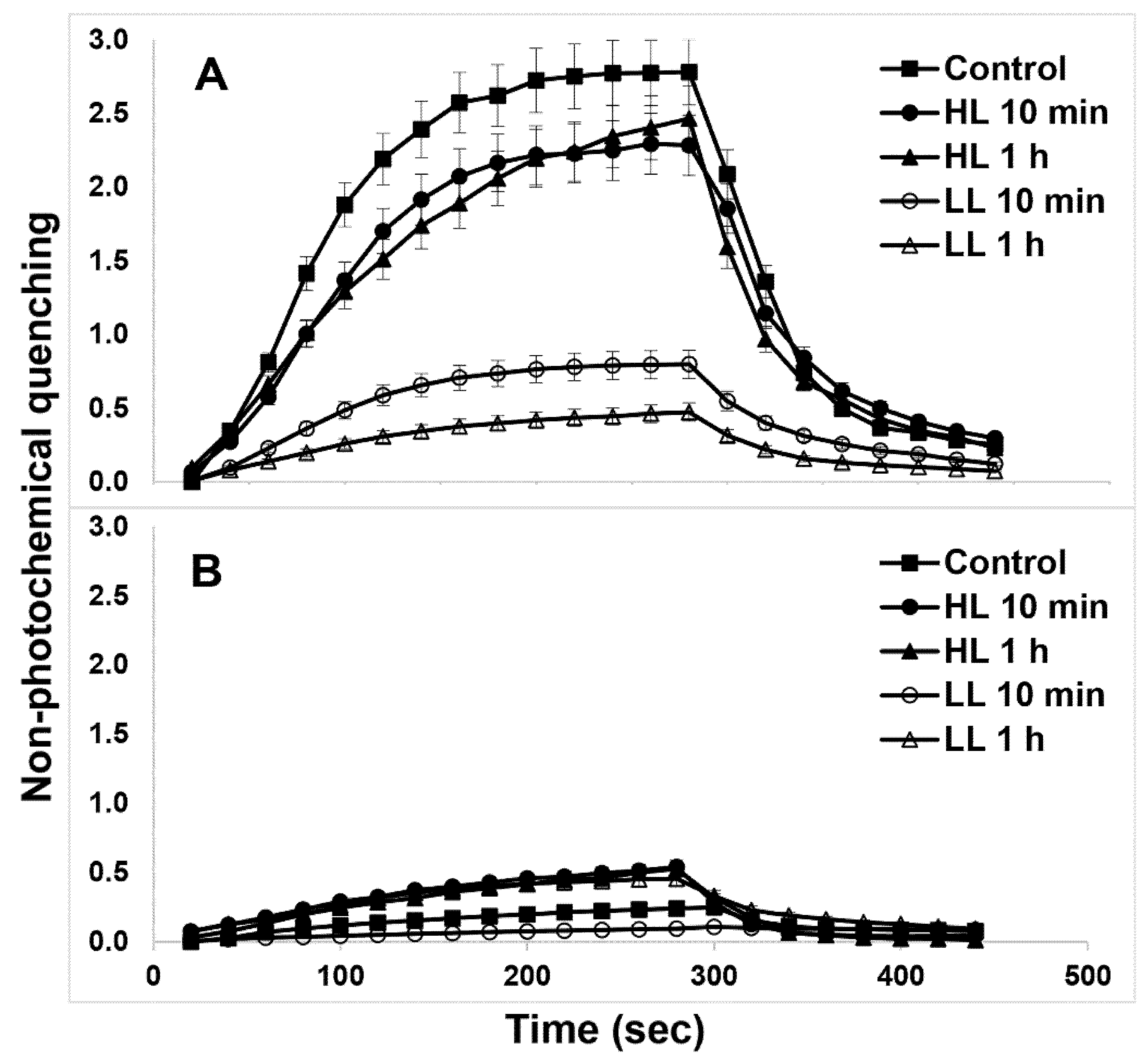
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Light Treatment
4.2. Isolation of Thylakoid Membranes and Chlorophyll Determination
4.3. Blue-Native Polyacrylamide Gel Electrophoresis
4.4. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis and Immunoblotting Analysis
4.5. Chlorophyll Fluorescence and Electron Transport Measurements
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Demmig-Adams, B.; Adams, W.W. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef]
- Crosatti, C.; Rizza, F.; Badeck, F.W.; Mazzucotelli, E.; Cattivelli, L. Harden the chloroplast to protect the plant. Physiol. Plant. 2013, 147, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zulfugarov, I.S.; Ham, O.K.; Mishra, S.R.; Kim, J.Y.; Nath, K.; Koo, H.Y.; Kim, H.S.; Moon, Y.H.; An, G.; Lee, C.H. Dependence of reaction center-type energy-dependent quenching on photosystem II antenna size. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 773–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruban, A.V. Nonphotochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malnoë, A. Photoinhibition or photoprotection of photosynthesis? Update on the (newly termed) sustained quenching component qH. Environ. Exp. Bot. 2018, 154, 123–133. [Google Scholar] [CrossRef]
- Zulfugarov, I.S.; Wu, G.; Tovuu, A.; Lee, C.H. Effect of oxygen on the non-photochemical quenching of vascular plants and potential oxygen deficiency in the stroma of PsbS-knock-out rice. Plant Sci. 2019, 286, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-P.; Björkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A Pigment-Binding Protein Essential for Regulation of Photosynthetic Light Harvesting. Nature 2000, 403, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Li, X.P.; Niyogi, K.K. Update on Photosynthesis Non-Photochemical Quenching. A Response to Excess Light Energy 1. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulfugarov, I.S.; Tovuu, A.; Eu, Y.J.; Dogsom, B.; Poudyal, R.S.; Nath, K.; Hall, M.; Banerjee, M.; Yoon, U.C.; Moon, Y.H.; et al. Production of superoxide from Photosystem II in a rice (Oryza sativa L.) mutant lacking PsbS. BMC Plant Biol. 2014, 14, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Hoang, M.H.; Jeon, Y.A.; Wu, G.; Lee, C.H. Differential down-regulation of zeaxanthin epoxidation in two rice (Oryza sativa L.) cultivars with different chilling sensitivities. J. Plant Biol. 2017, 60, 413–422. [Google Scholar] [CrossRef]
- Hoang, M.H.; Kim, H.S.; Zulfugarov, I.S.; Lee, C.H. Down-Regulation of Zeaxanthin Epoxidation in Vascular Plant Leaves Under Normal and Photooxidative Stress Conditions. J. Plant Biol. 2020, 63, 331–336. [Google Scholar] [CrossRef]
- Nilkens, M.; Kress, E.; Lambrev, P.; Miloslavina, Y.; Müller, M.; Holzwarth, A.R.; Jahns, P. Identification of a slowly inducible zeaxanthin-dependent component of non-photochemical quenching of chlorophyll fluorescence generated under steady-state conditions in Arabidopsis. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 466–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellaflore, S.; Barneche, F.; Peltler, G.; Rochalx, J.D. State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature 2005, 433, 892–895. [Google Scholar] [CrossRef]
- Heinrich Krause, G.; Somersalo, S.; Zumbusch, E.; Weyers, B.; Laasch, H. On the Mechanism of Photoinhibition in Chloroplasts. Relationship Between Changes in Fluorescence and Activity of Photosystem II. J. Plant Physiol. 1990, 136, 472–479. [Google Scholar] [CrossRef]
- Amstutz, C.L.; Fristedt, R.; Schultink, A.; Merchant, S.S.; Niyogi, K.K.; Malnoë, A. An atypical short-chain dehydrogenase–reductase functions in the relaxation of photoprotective qH in Arabidopsis. Nat. Plants 2020, 6, 154–166. [Google Scholar] [CrossRef]
- Kasahara, M.; Kagawa, T.; Olkawa, K.; Suetsugu, N.; Miyao, M.; Wada, M. Chlomplast avoidance movement reduces photodamage in plants. Nature 2002, 420, 829–832. [Google Scholar] [CrossRef]
- Cazzaniga, S.; Dall’Osto, L.; Kong, S.G.; Wada, M.; Bassi, R. Interaction between avoidance of photon absorption, excess energy dissipation and zeaxanthin synthesis against photooxidative stress in Arabidopsis. Plant J. 2013, 76, 568–579. [Google Scholar] [CrossRef]
- Albanese, P.; Manfredi, M.; Re, A.; Marengo, E.; Saracco, G.; Pagliano, C. Thylakoid proteome modulation in pea plants grown at different irradiances: Quantitative proteomic profiling in a non-model organism aided by transcriptomic data integration. Plant J. 2018, 96, 786–800. [Google Scholar] [CrossRef]
- Aliyeva, D.R.; Aydinli, L.M.; Pashayeva, A.N.; Zulfugarov, I.S.; Huseynova, I.M. Photosynthetic machinery and antioxidant status of wheat genotypes under drought stress followed by rewatering. Photosynthetica 2020, 58, 1217–1225. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A. Molecular design of the photosystem II light-harvesting antenna: Photosynthesis and photoprotection. J. Exp. Bot. 2005, 56, 365–373. [Google Scholar] [CrossRef]
- Kiss, A.Z.; Ruban, A.V.; Horton, P. The PsbS protein controls the organization of the photosystem II antenna in higher plant thylakoid membranes. J. Biol. Chem. 2008, 283, 3972–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betterle, N.; Ballottari, M.; Zorzan, S.; de Bianchi, S.; Cazzaniga, S.; Dall’Osto, L.; Morosinotto, T.; Bassi, R. Light-induced dissociation of an antenna hetero-oligomer is needed for non-photochemical quenching induction. J. Biol. Chem. 2009, 284, 15255–15266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffarri, S.; Kouřil, R.; Kereïche, S.; Boekema, E.J.; Croce, R. Functional architecture of higher plant photosystem II supercomplexes. EMBO J. 2009, 28, 3052–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kereïche, S.; Kiss, A.Z.; Kouřil, R.; Boekema, E.J.; Horton, P. The PsbS protein controls the macro-organisation of photosystem II complexes in the grana membranes of higher plant chloroplasts. FEBS Lett. 2010, 584, 759–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulfugarov, I.S.; Tovuu, A.; Dogsom, B.; Lee, C.Y.; Lee, C.H. PsbS-specific zeaxanthin-independent changes in fluorescence emission spectrum as a signature of energy-dependent non-photochemical quenching in higher plants. Photochem. Photobiol. Sci. 2010, 9, 697–703. [Google Scholar] [CrossRef]
- Ruban, A.V.; Berera, R.; Ilioaia, C.; Van Stokkum, I.H.M.; Kennis, J.T.M.; Pascal, A.A.; Van Amerongen, H.; Robert, B.; Horton, P.; Van Grondelle, R. Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature 2007, 450, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.K.; Avenson, T.J.; Ballottari, M.; Cheng, Y.C.; Niyogi, K.K.; Bassi, R.; Fleming, G.R. Architecture of a charge-transfer state regulating light harvesting in a plant antenna protein. Science 2008, 320, 794–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzwarth, A.R.; Miloslavina, Y.; Nilkens, M.; Jahns, P. Identification of two quenching sites active in the regulation of photosynthetic light-harvesting studied by time-resolved fluorescence. Chem. Phys. Lett. 2009, 483, 262–267. [Google Scholar] [CrossRef]
- Gerotto, C.; Franchin, C.; Arrigoni, G.; Morosinotto, T. In vivo identification of photosystem II light harvesting complexes interacting with PHOTOSYSTEM II SUBUNIT S. Plant Physiol. 2015, 168, 1747–1772. [Google Scholar] [CrossRef] [Green Version]
- Correa-Galvis, V.; Poschmann, G.; Melzer, M.; Stühler, K.; Jahns, P. PsbS interactions involved in the activation of energy dissipation in Arabidopsis. Nat. Plants 2016, 2, 1–8. [Google Scholar] [CrossRef]
- Sacharz, J.; Giovagnetti, V.; Ungerer, P.; Mastroianni, G.; Ruban, A.V. The xanthophyll cycle affects reversible interactions between PsbS and light-harvesting complex II to control non-photochemical quenching. Nat. Plants 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Trotta, A.; Bajwa, A.A.; Mancini, I.; Paakkarinen, V.; Pribil, M.; Aro, E.M. The role of phosphorylation dynamics of curvature thylakoid 1B in plant thylakoid membranes. Plant Physiol. 2019, 181, 1615–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S. Non-Photochemical Quenching Mechanisms in Plants-Light Induced Reorganization of the Thylakoid Membrane. Ph.D. Thesis, University of Zurich, Zürich, Switzerland, 2014; pp. 1–170, Volume 61. [Google Scholar]
- Holzwarth, A.R.; Jahns, P. Non-Photochemical Quenching Mechanisms in Intact Organisms as Derived from Ultrafast-Fluorescence Kinetic Studies. In Non-Photochemical Quenching and Energy dissipation In Plants, Algae and Cyanobacteria; Springer: Dordrecht, The Netherlands, 2014; Volume 21, pp. 129–156. [Google Scholar] [CrossRef]
- Anderson, J.M. Lateral heterogeneity of plant thylakoid protein complexes: Early reminiscences. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3384–3388. [Google Scholar] [CrossRef] [Green Version]
- Pesaresi, P.; Pribil, M.; Wunder, T.; Leister, D. Dynamics of reversible protein phosphorylation in thylakoids of flowering plants: The roles of STN7, STN8 and TAP38. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochaix, J.D.; Lemeille, S.; Shapiguzov, A.; Samol, I.; Fucile, G.; Willig, A.; Goldschmidt-Clermont, M. Protein kinases and phosphatases involved in the acclimation of the photosynthetic apparatus to a changing light environment. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3466–3474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, W.S.; Kim, E.H.; Horton, P.; Anderson, J.M. Granal stacking of thylakoid membranes in higher plant chloroplasts: The physicochemical forces at work and the functional consequences that ensue. Photochem. Photobiol. Sci. 2005, 4, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Sakamoto, W. Phosphorylation of photosystem II core proteins prevents undesirable cleavage of D1 and contributes to the fine-tuned repair of photosystem II. Plant J. 2014, 79, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Nath, K.; Mishra, S.R.; Zulfugarov, I.S.; Lee, C.B.; An, G.; Lee, C.H. Characterization of a T-DNA Inserted STN8 Kinase Mutant of Oryza sativa L. In Photosynthesis. Energy from the Sun; Springer: Dordrecht, The Netherlands, 2008; pp. 1307–1311. [Google Scholar] [CrossRef]
- Grieco, M.; Suorsa, M.; Jajoo, A.; Tikkanen, M.; Aro, E.M. Light-harvesting II antenna trimers connect energetically the entire photosynthetic machinery-Including both photosystems II and i. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Suorsa, M.; Rantala, M.; Mamedov, F.; Lespinasse, M.; Trotta, A.; Grieco, M.; Vuorio, E.; Tikkanen, M.; Järvi, S.; Aro, E.M. Light acclimation involves dynamic re-organization of the pigment-protein megacomplexes in non-appressed thylakoid domains. Plant J. 2015, 84, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Yokono, M.; Takabayashi, A.; Akimoto, S.; Tanaka, A. A megacomplex composed of both photosystem reaction centres in higher plants. Nat. Commun. 2015, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tikkanen, M.; Aro, E.-M. Integrative regulatory network of plant thylakoid energy transduction. Trends Plant Sci. 2014, 19, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Fristedt, R.; Willig, A.; Granath, P.; Crèvecoeur, M.; Rochaix, J.D.; Vener, A.V. Phosphorylation of photosystem II controls functional macroscopic folding of photosynthetic membranes in Arabidopsis. Plant Cell 2009, 21, 3950–3964. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Horton, P.; Kim, E.H.; Chow, W.S. Towards elucidation of dynamic structural changes of plant thylakoid architecture. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3515–3524. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, M.; Aro, E.M. Thylakoid protein phosphorylation in dynamic regulation of photosystem II in higher plants. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 232–238. [Google Scholar] [CrossRef] [Green Version]
- Goral, T.K.; Johnson, M.P.; Brain, A.P.R.; Kirchhoff, H.; Ruban, A.V.; Mullineaux, C.W. Visualizing the mobility and distribution of chlorophyll proteins in higher plant thylakoid membranes: Effects of photoinhibition and protein phosphorylation. Plant J. 2010, 6, 948–959. [Google Scholar] [CrossRef] [Green Version]
- Baena-González, E.; Barbato, R.; Aro, E.M. Role of phosphorylation in the repair cycle and oligomeric structure of photosystem II. Planta 1999, 208, 196–204. [Google Scholar] [CrossRef]
- Tikkanen, M.; Nurmi, M.; Kangasjärvi, S.; Aro, E.M. Core protein phosphorylation facilitates the repair of photodamaged photosystem II at high light. Biochim. Biophys. Acta Bioenerg. 2008, 1777, 1432–1437. [Google Scholar] [CrossRef] [Green Version]
- Breitholtz, H.L.; Srivastava, R.; Tyystjärvi, E.; Rintamäki, E. LHC II protein phosphorylation in leaves of Arabidopsis thaliana mutants deficient in non-photochemical quenching. Photosynth. Res. 2005, 84, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Gollan, P.J.; Aro, E.M. Photosynthetic signalling during high light stress and recovery: Targets and dynamics. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Savitch, L.V.; Ivanov, A.G.; Krol, M.; Sprott, D.P.; Quist, G.; Huner, N.P.A. Regulation of energy partitioning and alternative electron transport pathways during cold acclimation of Lodgepole pine is oxygen dependent. Plant Cell Physiol. 2010, 51, 1555–1570. [Google Scholar] [CrossRef]
- Järvi, S.; Suorsa, M.; Paakkarinen, V.; Aro, E.M. Optimized native gel systems for separation of thylakoid protein complexes: Novel super- and mega-complexes. Biochem. J. 2011, 439, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Galka, P.; Santabarbara, S.; Khuong, T.T.H.; Degand, H.; Morsomme, P.; Jennings, R.C.; Boekema, E.J.; Caffarria, S. Functional analyses of the plant photosystem I-light-harvesting complex II supercomplex reveal that light-harvesting complex II loosely bound to photosystem ii is a very efficient antenna for photosystem I in state II. Plant Cell 2012, 24, 2963–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longoni, P.; Douchi, D.; Cariti, F.; Fucile, G.; Goldschmidt-Clermont, M. Phosphorylation of the light-harvesting complex II isoform Lhcb2 is central to state transitions. Plant Physiol. 2015, 169, 2874–2883. [Google Scholar] [CrossRef] [PubMed]
- Leoni, C.; Pietrzykowska, M.; Kiss, A.Z.; Suorsa, M.; Ceci, L.R.; Aro, E.-M.; Jansson, S. Very rapid phosphorylation kinetics suggest a unique role for Lhcb2 during state transitions in Arabidopsis. Plant J. 2013, 76, 236–246. [Google Scholar] [CrossRef]
- Murchie, E.H.; Niyogi, K.K. Manipulation of photoprotection to improve plant photosynthesis. Plant Physiol. 2011, 155, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulfugarov, I.S.; Tovuu, A.; Lee, C.H. Acceleration of cyclic electron flow in rice plants (Oryza sativa L.) deficient in the PsbS protein of Photosystem II. Plant Physiol. Biochem. 2014, 84, 233–239. [Google Scholar] [CrossRef]
- Zulfugarov, I.S.; Tovuu, A.; Kim, C.Y.; Xuan Vo, K.T.; Ko, S.Y.; Hall, M.; Seok, H.Y.; Kim, Y.K.; Skogstrom, O.; Moon, Y.H.; et al. Enhanced resistance of PsbS-deficient rice (Oryza sativa L.) to fungal and bacterial pathogens. J. Plant Biol. 2016, 59, 616–626. [Google Scholar] [CrossRef]
- Grebe, S.; Trotta, A.; Bajwa, A.A.; Mancini, I.; Bag, P.; Jansson, S.; Tikkanen, M.; Aro, E.M. Specific thylakoid protein phosphorylations are prerequisites for overwintering of Norway spruce (Picea abies) photosynthesis. Proc. Natl. Acad. Sci. USA 2020, 117, 17499–17509. [Google Scholar] [CrossRef] [PubMed]
- Rantala, M.; Tikkanen, M.; Aro, E.M. Proteomic characterization of hierarchical megacomplex formation in Arabidopsis thylakoid membrane. Plant J. 2017, 92, 951–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damkjær, J.T.; Kereïche, S.; Johnson, M.P.; Kovacs, L.; Kiss, A.Z.; Boekema, E.J.; Ruban, A.V.; Horton, P.; Jansson, S. The photosystem II light-harvesting protein Lhcb3 affects the macrostructure of photosystem II and the rate of state transitions in Arabidopsis. Plant Cell 2009, 21, 3245–3256. [Google Scholar] [CrossRef] [Green Version]
- Pietrzykowska, M.; Suorsa, M.; Semchonok, D.A.; Tikkanen, M.; Boekema, E.J.; Aro, E.M.; Jansson, S. The light-harvesting chlorophyll a/b binding proteins Lhcb1 and Lhcb2 play complementary roles during state transitions in Arabidopsis. Plant Cell 2014, 26, 3646–3660. [Google Scholar] [CrossRef] [Green Version]
- Betterle, N.; Ballottari, M.; Baginsky, S.; Bassi, R. High light-dependent phosphorylation of photosystem II inner antenna CP29 in monocots is STN7 independent and enhances nonphotochemical quenching. Plant Physiol. 2015, 167, 457–471. [Google Scholar] [CrossRef] [Green Version]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. BBA Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Yang, Y.N.; Safarova, R.B.; Park, S.Y.; Sakuraba, Y.; Oh, M.H.; Zulfugarov, I.S.; Lee, C.B.; Tanaka, A.; Paek, N.C.; Lee, C.H. Chlorophyll Degradation and Light-harvesting Complex II Aggregate Formation During Dark-induced Leaf Senescence in Arabidopsis Pheophytinase Mutants. J. Plant Biol. 2019, 62, 27–38. [Google Scholar] [CrossRef]
- Oh, M.H.; Safarova, R.B.; Eu, Y.J.; Zulfugarov, I.S.; Kim, J.H.; Hwang, H.J.; Lee, C.B.; Lee, C.H. Loss of peripheral polypeptides in the stromal side of photosystem i by light-chilling in cucumber leaves. Photochem. Photobiol. Sci. 2009, 8, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef]
- Tovuu, A.; Zulfugarov, I.S.; Wu, G.; Kang, I.S.; Kim, C.; Moon, B.Y.; An, G.; Lee, C.H. Rice mutants deficient in ω-3 fatty acid desaturase (FAD8) fail to acclimate to cold temperatures. Plant Physiol. Biochem. 2016, 109, 525–535. [Google Scholar] [CrossRef]
- Peng, L.; Shimizu, H.; Shikanai, T. The chloroplast NAD(P)H dehydrogenase complex interacts with photosystem I in Arabidopsis. J. Biol. Chem. 2008, 283, 34873–34879. [Google Scholar] [CrossRef] [Green Version]
- Sipka, G.; Magyar, M.; Mezzetti, A.; Akhtar, P.; Zhu, Q.; Xiao, Y.; Han, G.; Santabarbara, S.; Shen, J.R.; Lambrev, P.H.; et al. Light-adapted charge-separated state of photosystem II: Structural and functional dynamics of the closed reaction center. Plant Cell 2021, 33, 1286–1302. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | R | τ1 | A1 | τ2 | A2 | |
|---|---|---|---|---|---|---|
| Wild type | Control | 0.997 ± 0.001 | 25.2 ± 2.1 | 2.51 ± 0.11 | 1577 ± 145 | 0.64 ± 0.08 |
| 10 min HL | 0.988 ± 0.001 | 53.9 ± 6.4 | 2.19 ± 0.09 | 51,853 ± 682 | 0.39 ± 0.05 | |
| 1 h HL | 0.996 ± 0.001 | 33.1 ± 2.7 | 1.98 ± 0.12 | 326 ± 24 | 0.69 ± 0.09 | |
| 10 min LL | 0.997 ± 0.001 | 30.7 ± 3.2 | 0.47 ± 0.08 | 3344 ± 287 | 4.29 ± 0.21 | |
| 1 h LL | 0.997 ± 0.001 | 33.1 ± 4.1 | 0.35 ± 0.09 | 1149 ± 98 | 0.43 ± 0.11 | |
| PsbS-KO | Control | 0.998 ± 0.001 | 21.8 ± 2.2 | 0.16 ± 0.05 | 2345 ± 186 | 0.29 ± 0.04 |
| 10 min HL | 0.997 ± 0.001 | 24.8 ± 2.7 | 0.51 ± 0.07 | n.d. | 0.02 ± 0.00 | |
| 1 h HL | 0.994 ± 0.001 | 36.5 ± 4.4 | 0.60 ± 0.09 | 52.11 ± 0.19 | 0.002 ± 0.00 | |
| 10 min LL | 0.953 ± 0.001 | 37.5 ± 3.6 | 0.04 ± 0.03 | n.d. | 0.004 ± 0.00 | |
| 1 h LL | 0.996 ± 0.001 | 23.7 ± 2.5 | 0.06 ± 0.03 | 49.39 ± 0.21 | 0.31 ± 0.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pashayeva, A.; Wu, G.; Huseynova, I.; Lee, C.-H.; Zulfugarov, I.S. Role of Thylakoid Protein Phosphorylation in Energy-Dependent Quenching of Chlorophyll Fluorescence in Rice Plants. Int. J. Mol. Sci. 2021, 22, 7978. https://doi.org/10.3390/ijms22157978
Pashayeva A, Wu G, Huseynova I, Lee C-H, Zulfugarov IS. Role of Thylakoid Protein Phosphorylation in Energy-Dependent Quenching of Chlorophyll Fluorescence in Rice Plants. International Journal of Molecular Sciences. 2021; 22(15):7978. https://doi.org/10.3390/ijms22157978
Chicago/Turabian StylePashayeva, Aynura, Guangxi Wu, Irada Huseynova, Choon-Hwan Lee, and Ismayil S. Zulfugarov. 2021. "Role of Thylakoid Protein Phosphorylation in Energy-Dependent Quenching of Chlorophyll Fluorescence in Rice Plants" International Journal of Molecular Sciences 22, no. 15: 7978. https://doi.org/10.3390/ijms22157978