
Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages

 , and
, and 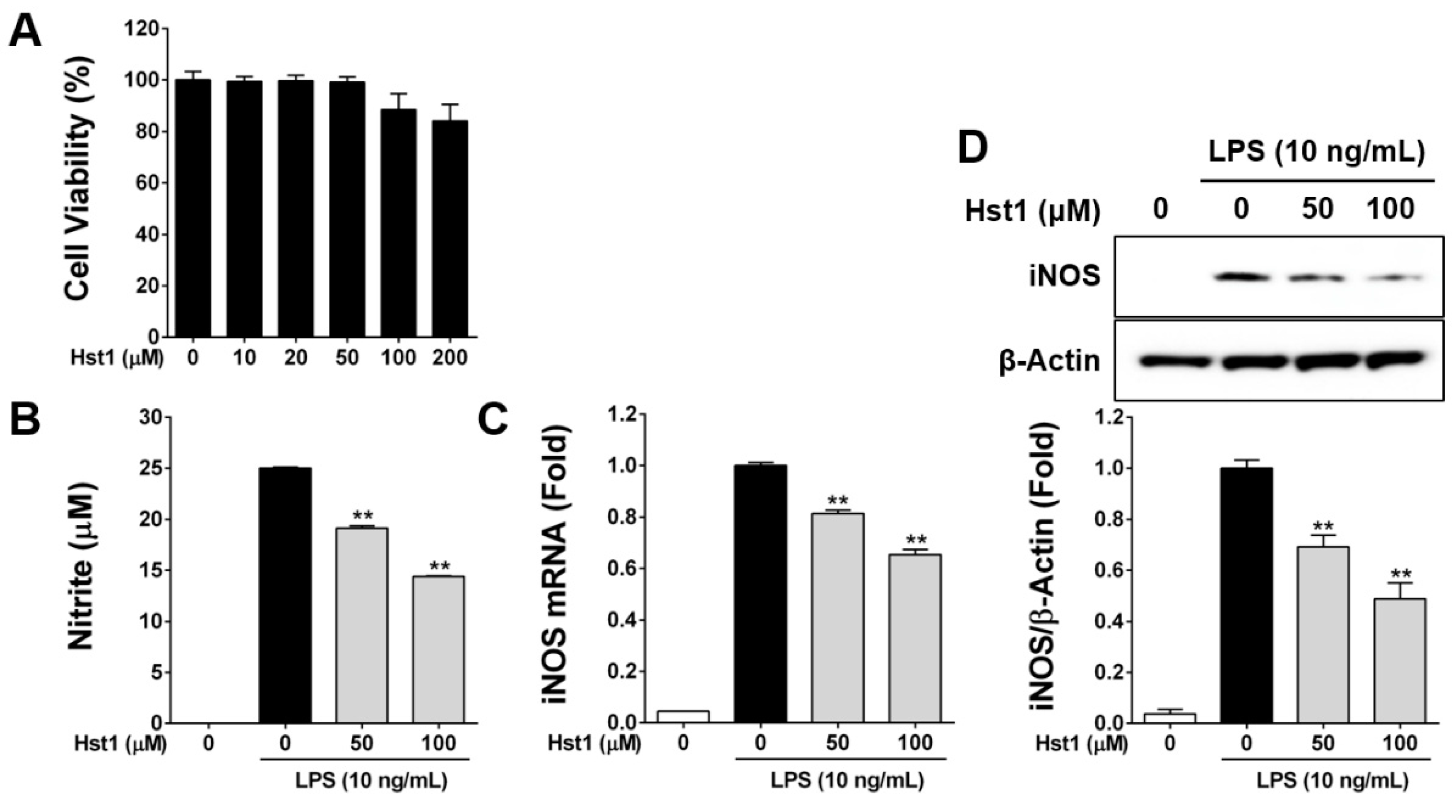{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Histatin-1 Reduces LPS-Induced NO Production
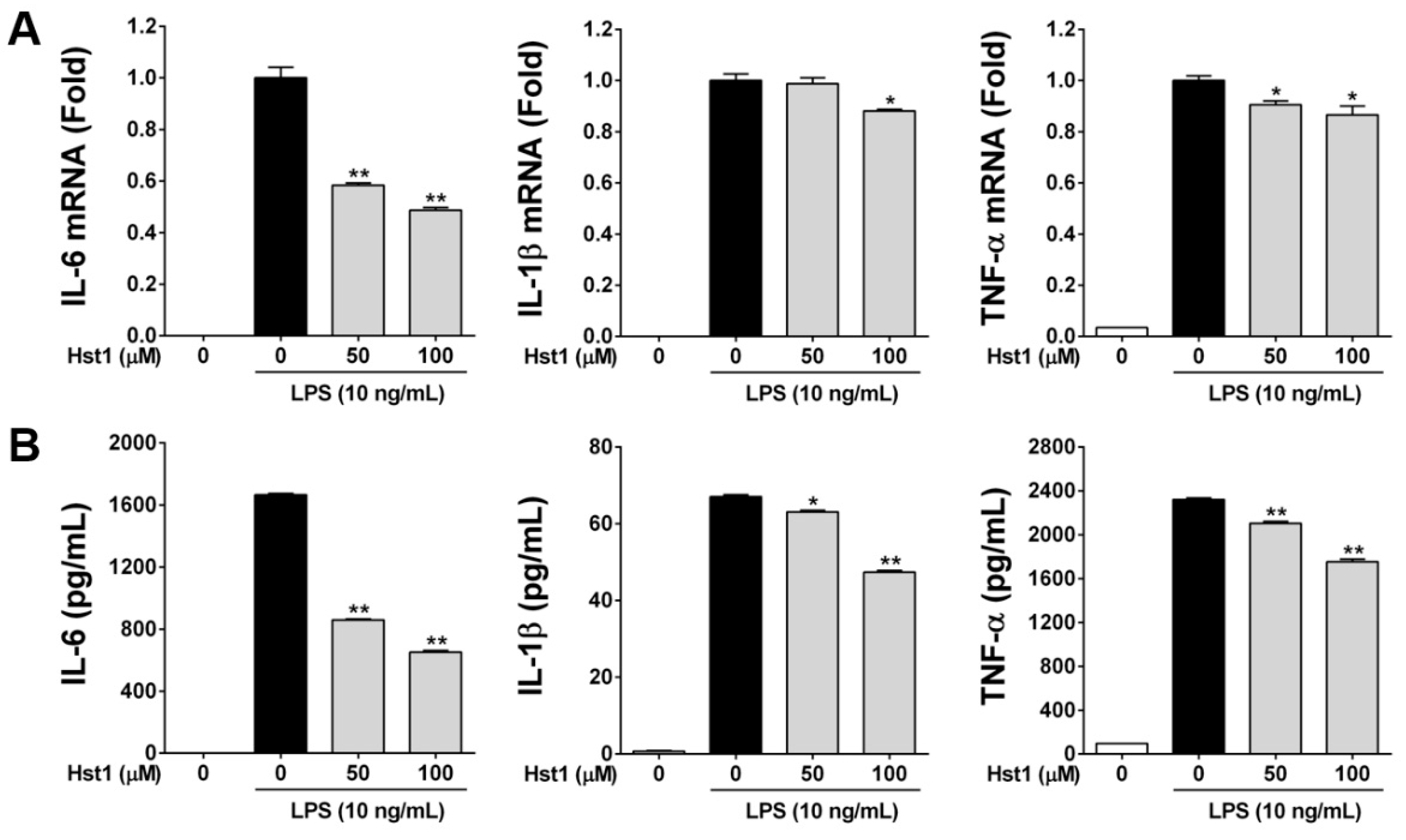
2.2. Histatin-1 Reduces LPS-Induced Inflammatory Cytokine Production
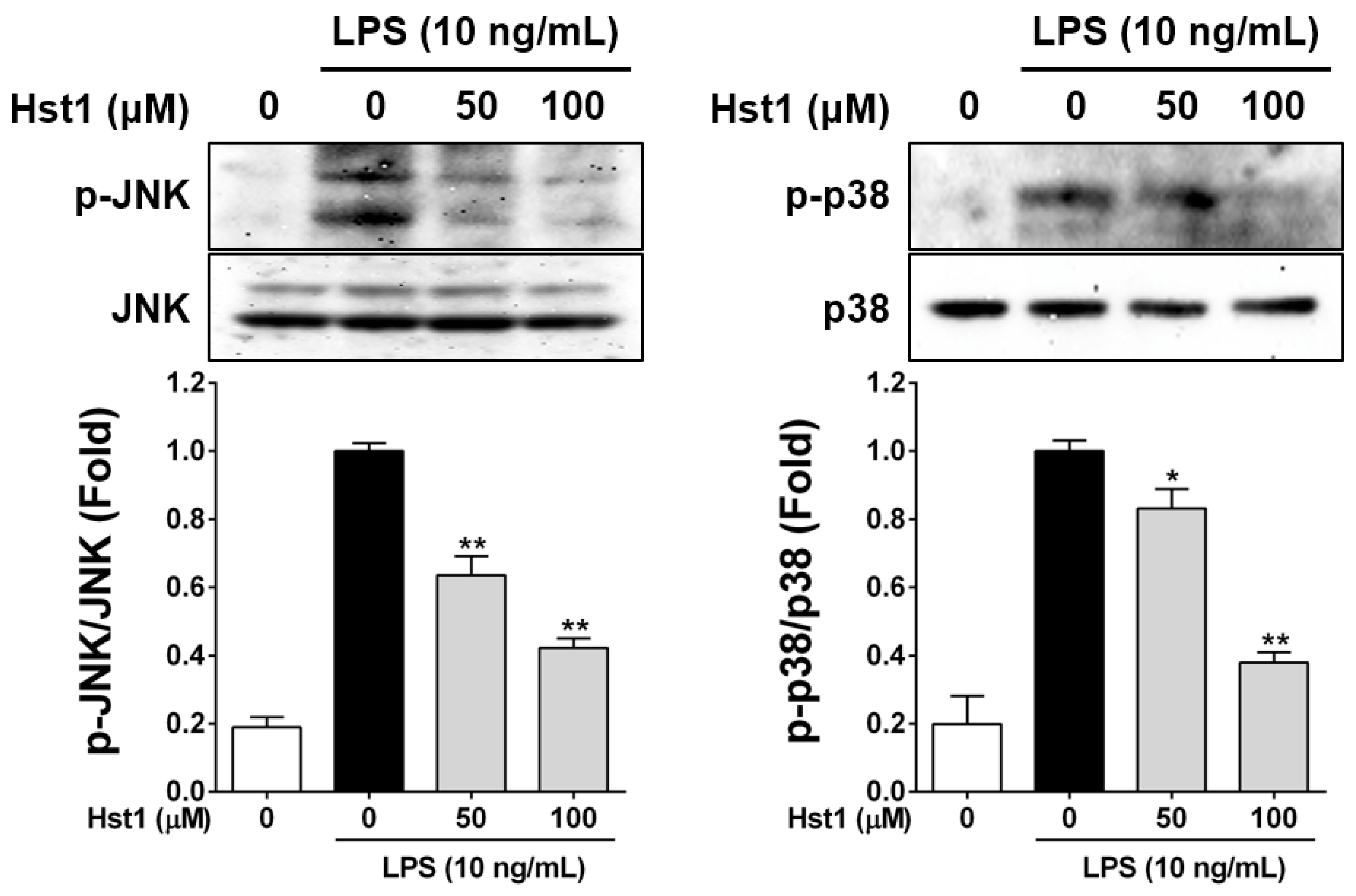
2.3. Histatin-1 Reduces LPS-Induced MAPK Activation
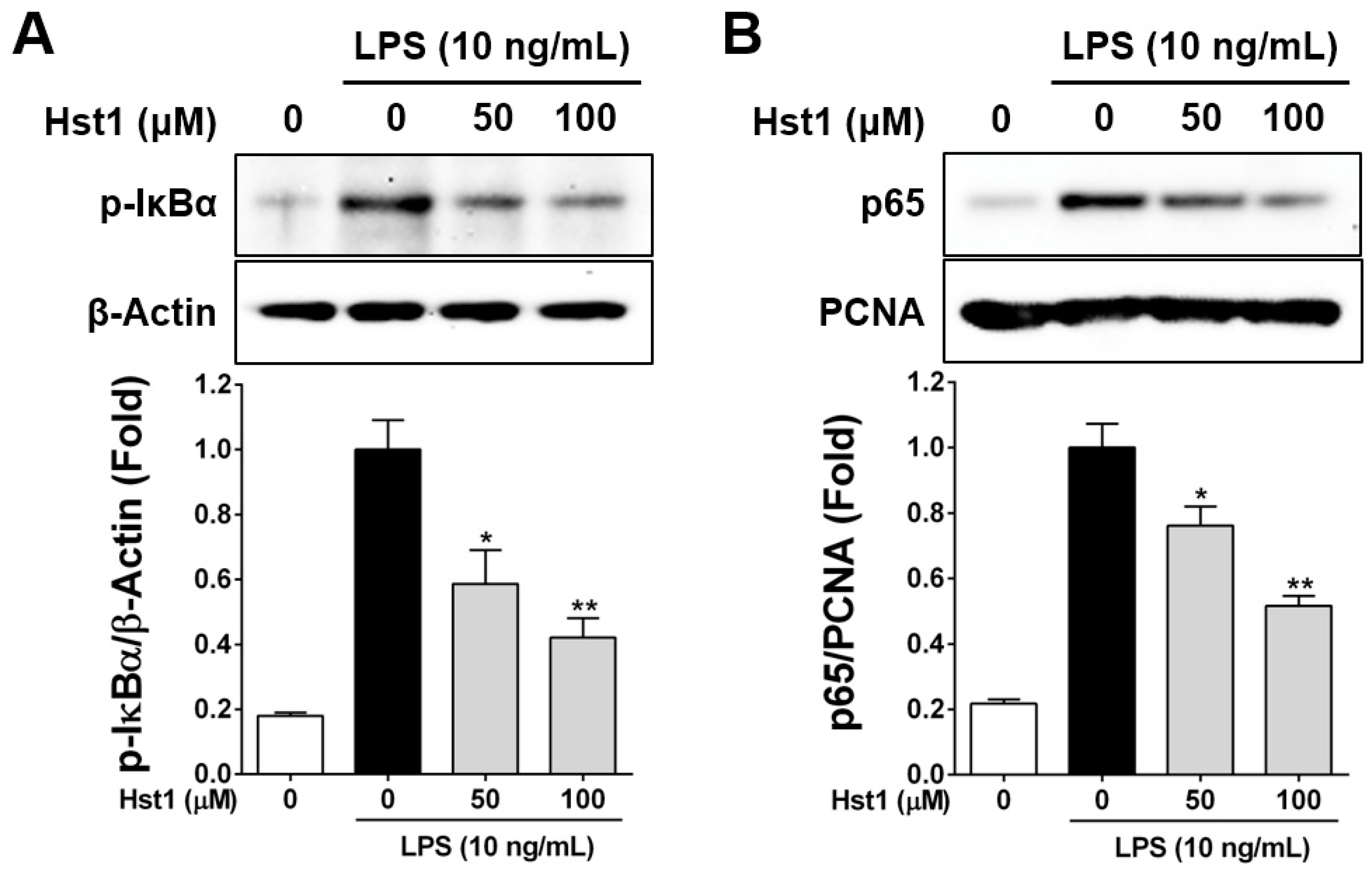
2.4. Histatin-1 Reduces LPS-Induced NF-kB Activation
3. Discussion
4. Materials and Methods
4.1. Materials and Cell Culture
4.2. Viability of RAW264.7 Macrophage Cells
4.3. Measurement of NO Production
4.4. Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.5. Western Blot Analysis
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Eshac, Y.; Redfern, R.L.; Aakalu, V.K. The Role of Endogenous Antimicrobial Peptides in Modulating Innate Immunity of the Ocular Surface in Dry Eye Diseases. Int. J. Mol. Sci. 2021, 22, 721. [Google Scholar] [CrossRef] [PubMed]
- Pussinen, P.J.; Vilkuna-Rautiainen, T.; Alfthan, G.; Palosuo, T.; Jauhiainen, M.; Sundvall, J.; Vesanen, M.; Mattila, K.; Asikainen, S. Severe periodontitis enhances macrophage activation via increased serum lipopolysaccharide. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2174–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, K.T.; Xiao, Y.; Pflugfelder, S.C.; de Paiva, C.S. Inflammatory response to lipopolysaccharide on the ocular surface in a murine dry eye model. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2443–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, I.-C.; Coursey, T.G.; Bian, F.; Barbosa, F.L.; de Paiva, C.S.; Pflugfelder, S.C. Macrophage phenotype in the ocular surface of experimental murine dry eye disease. Arch. Immunol. Ther. Exp. 2015, 63, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Chen, Y.-T.; Chen, F.; Gallup, M.; Vijmasi, T.; Bahrami, A.F.; Noble, L.B.; van Rooijen, N.; McNamara, N.A. Critical Involvement of Macrophage Infiltration in the Development of Sjögren’s Syndrome–Associated Dry Eye. Am. J. Pathol. 2012, 181, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, W.; Chauhan, S.K.; Dana, R. Dry eye disease: An immune-mediated ocular surface disorder. Arch. Ophthalmol. 2012, 130, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Garaicoa-Pazmino, C.; Fretwurst, T.; Squarize, C.H.; Berglundh, T.; Giannobile, W.V.; Larsson, L.; Castilho, R.M. Characterization of macrophage polarization in periodontal disease. J. Clin. Periodontol. 2019, 46, 830–839. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, Y.; Duan, D.; Wang, P.; Xin, Y.; Bai, L.; Liu, Y.; Xu, Y. Enhanced activity of macrophage M1/M2 phenotypes in periodontitis. Arch. Oral Biol. 2018, 96, 234–242. [Google Scholar] [CrossRef]
- Almubarak, A.; Tanagala, K.K.K.; Papapanou, P.N.; Lalla, E.; Momen-Heravi, F. Disruption of monocyte and macrophage homeostasis in periodontitis. Front. Immunol. 2020, 11, 330. [Google Scholar] [CrossRef]
- Redfern, R.L.; Barabino, S.; Baxter, J.; Lema, C.; McDermott, A.M. Dry eye modulates the expression of toll-like receptors on the ocular surface. Exp. Eye Res. 2015, 134, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Redfern, R.L.; McDermott, A.M. Toll-like receptors in ocular surface disease. Exp. Eye Res. 2010, 90, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Redfern, R.L.; Patel, N.; Hanlon, S.; Farley, W.; Gondo, M.; Pflugfelder, S.C.; McDermott, A.M. Toll-like receptor expression and activation in mice with experimental dry eye. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1554–1563. [Google Scholar] [CrossRef] [PubMed]
- Redfern, R.L.; Reins, R.Y.; McDermott, A.M. Toll-like receptor activation modulates antimicrobial peptide expression by ocular surface cells. Exp. Eye Res. 2011, 92, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Reins, R.Y.; Courson, J.; Lema, C.; Redfern, R.L. MyD88 contribution to ocular surface homeostasis. PLoS ONE 2017, 12, e0182153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, P.; Castro, M.; Reyes, M.; Torres, V. Histatins, wound healing, and cell migration. Oral Dis. 2018, 24, 1150–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.; Ali, M.; Pasha, Z.; Jaboori, A.J.; Jassim, S.H.; Jain, S.; Aakalu, V.K. Histatin-1 expression in human lacrimal epithelium. PLoS ONE 2016, 11, e0148018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.; Ali, M.; Shukla, D.; Jain, S.; Aakalu, V.K. Effects of histatin-1 peptide on human corneal epithelial cells. PLoS ONE 2017, 12, e0178030. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.; Son, K.-N.; Kalmodia, S.; Lee, B.-S.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Wound Healing Properties of Histatin-5 and Identification of a Functional Domain Required for Histatin-5-Induced Cell Migration. Mol. Ther. Methods Clin. Dev. 2020, 17, 709–716. [Google Scholar] [CrossRef]
- Van Dijk, I.A.; Veerman, E.C.; Reits, E.A.; Bolscher, J.G.; Stap, J. Salivary peptide histatin 1 mediated cell adhesion: A possible role in mesenchymal-epithelial transition and in pathologies. Biol. Chem. 2018, 399, 1409–1419. [Google Scholar] [CrossRef]
- Torres, P.; Díaz, J.; Arce, M.; Silva, P.; Mendoza, P.; Lois, P.; Molina-Berríos, A.; Owen, G.I.; Palma, V.; Torres, V.A. The salivary peptide histatin-1 promotes endothelial cell adhesion, migration, and angiogenesis. FASEB J. 2017, 31, 4946–4958. [Google Scholar] [CrossRef] [Green Version]
- Gusman, H.; Travis, J.; Helmerhorst, E.J.; Potempa, J.; Troxler, R.F.; Oppenheim, F.G. Salivary histatin 5 is an inhibitor of both host and bacterial enzymes implicated in periodontal disease. Infect. Immun. 2001, 69, 1402–1408. [Google Scholar] [CrossRef] [Green Version]
- Khurshid, Z.; Najeeb, S.; Mali, M.; Moin, S.F.; Raza, S.Q.; Zohaib, S.; Sefat, F.; Zafar, M.S. Histatin peptides: Pharmacological functions and their applications in dentistry. Saudi Pharm. J. 2017, 25, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Kalmodia, S.; Son, K.-N.; Cao, D.; Lee, B.-S.; Surenkhuu, B.; Shah, D.; Ali, M.; Balasubramaniam, A.; Jain, S.; Aakalu, V.K. Presence of histatin-1 in human tears and association with aqueous deficient dry eye diagnosis: A preliminary study. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, K. Anti-lipopolysaccharide activity of histatins, peptides from human saliva. Experientia 1993, 49, 1095–1097. [Google Scholar] [CrossRef]
- Chih, Y.-H.; Wang, S.-Y.; Yip, B.-S.; Cheng, K.-T.; Hsu, S.-Y.; Wu, C.-L.; Yu, H.-Y.; Cheng, J.-W. Dependence on size and shape of non-nature amino aciD.S. in the enhancement of lipopolysaccharide (LPS) neutralizing activities of antimicrobial peptides. J. Colloid Interface Sci. 2019, 533, 492–502. [Google Scholar] [CrossRef]
- Inomata, M.; Into, T.; Murakami, Y. Suppressive effect of the antimicrobial peptide LL-37 on expression of IL-6, IL-8 and CXCL10 induced by Porphyromonas gingivalis cells and extracts in human gingival fibroblasts. Eur. J. Oral Sci. 2010, 118, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yuan, W.; Liu, H.; Huang, S.; Bian, L.; Guo, R. Injectable supramolecular gelatin hydrogel loading of resveratrol and histatin-1 for burn wound therapy. Biomater. Sci. 2020, 8, 4810–4820. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.M.; Son, K.-N.; Shah, D.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages. Int. J. Mol. Sci. 2021, 22, 7856. https://doi.org/10.3390/ijms22157856
Lee SM, Son K-N, Shah D, Ali M, Balasubramaniam A, Shukla D, Aakalu VK. Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages. International Journal of Molecular Sciences. 2021; 22(15):7856. https://doi.org/10.3390/ijms22157856
Chicago/Turabian StyleLee, Sang Min, Kyung-No Son, Dhara Shah, Marwan Ali, Arun Balasubramaniam, Deepak Shukla, and Vinay Kumar Aakalu. 2021. "Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages" International Journal of Molecular Sciences 22, no. 15: 7856. https://doi.org/10.3390/ijms22157856