
Improved Transformation and Regeneration of Indica Rice: Disruption of SUB1A as a Test Case via CRISPR-Cas9

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of Callus Induction Medium for Mature Seeds
2.2. Optimization of Regeneration Medium for Mature Seeds
2.3. Regeneration Performance of Immature Embryo from Agrobacterium-Mediated Transformations Using the Optimized Shoot Induction Medium
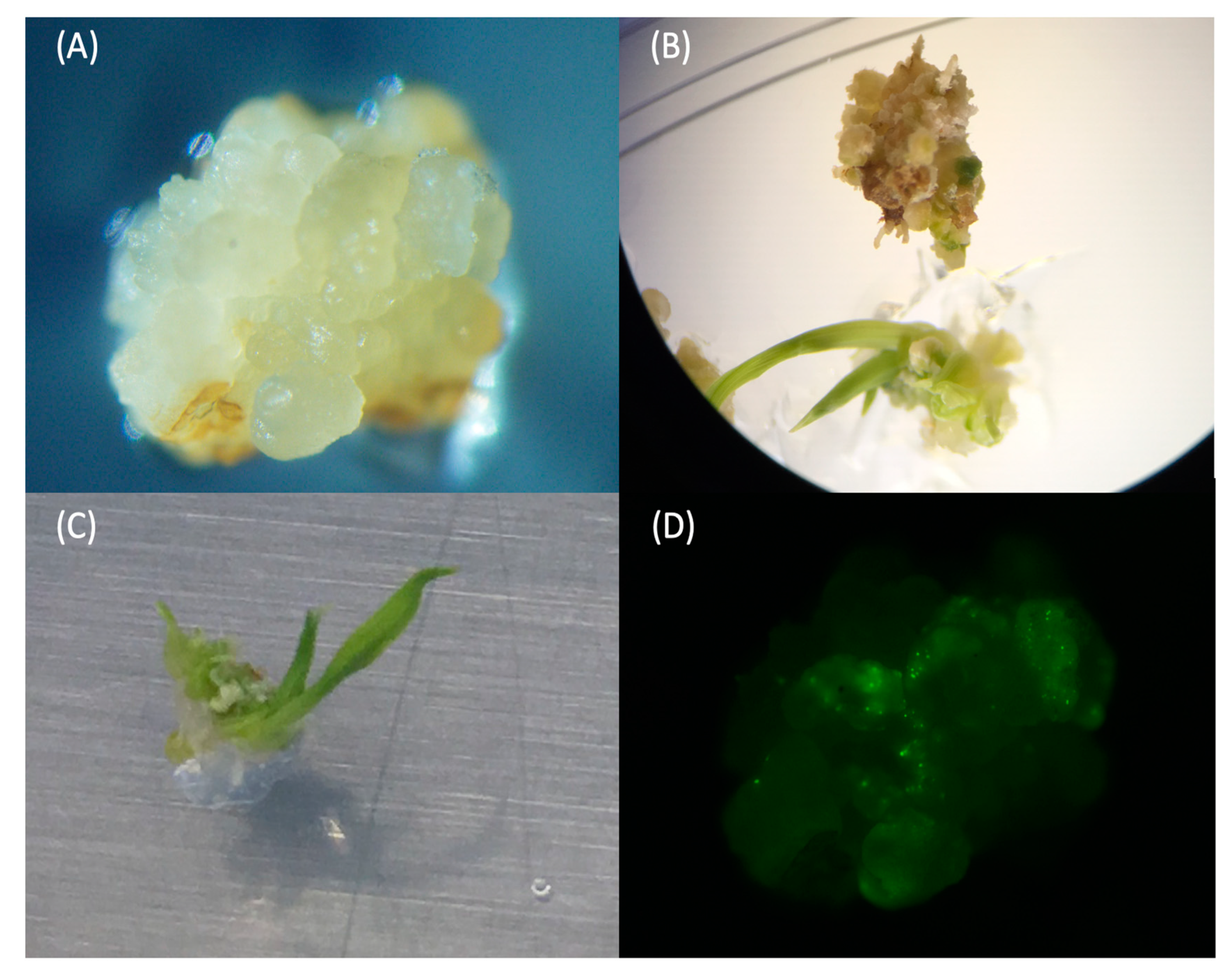
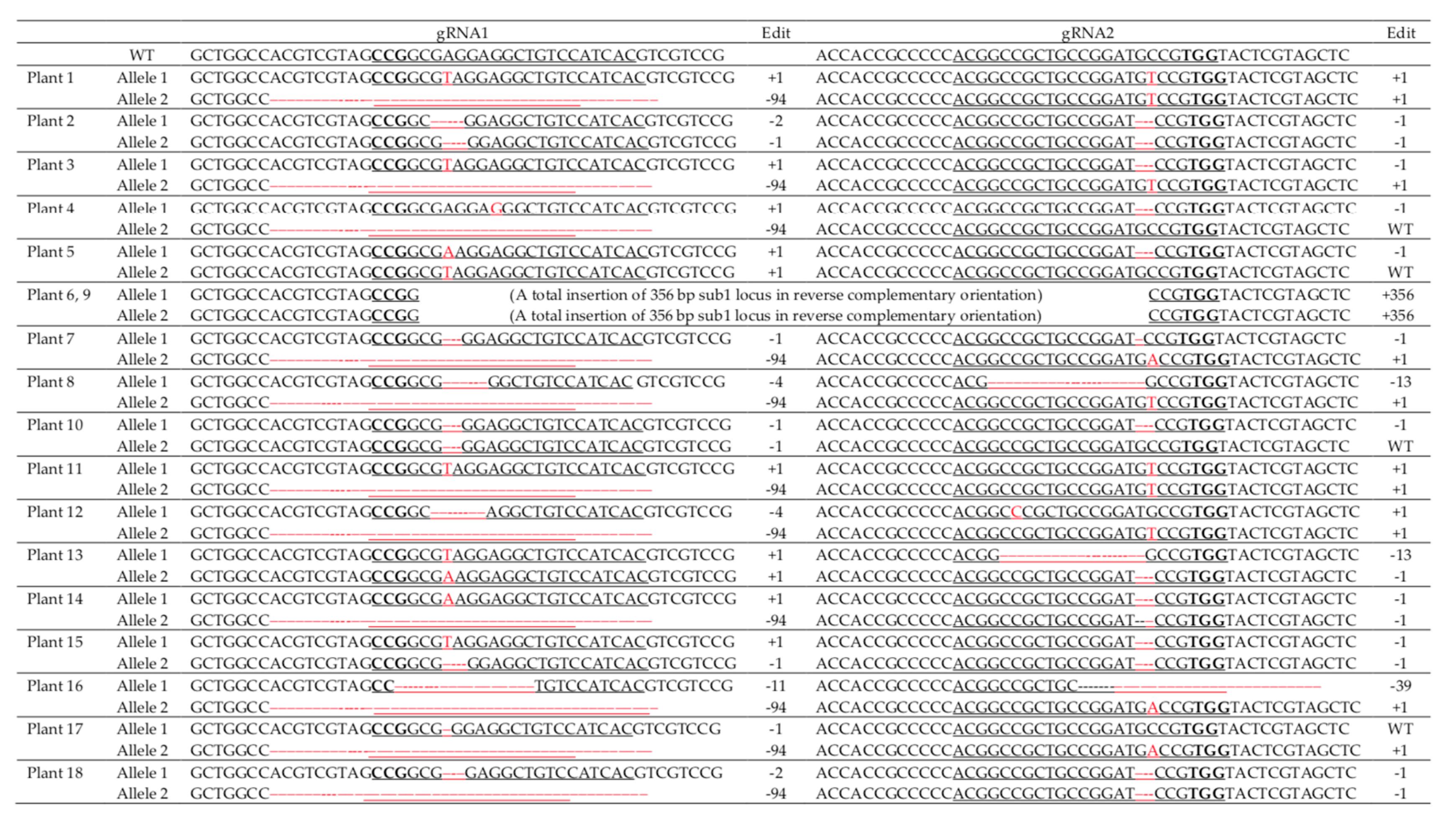
2.4. Validation of Transgenic Plants Derived from Immature Embryos
2.5. Optimization of Biolistic Bombardment Procedures
2.6. Validation of Transgenic Plants Derived from Mature Seeds
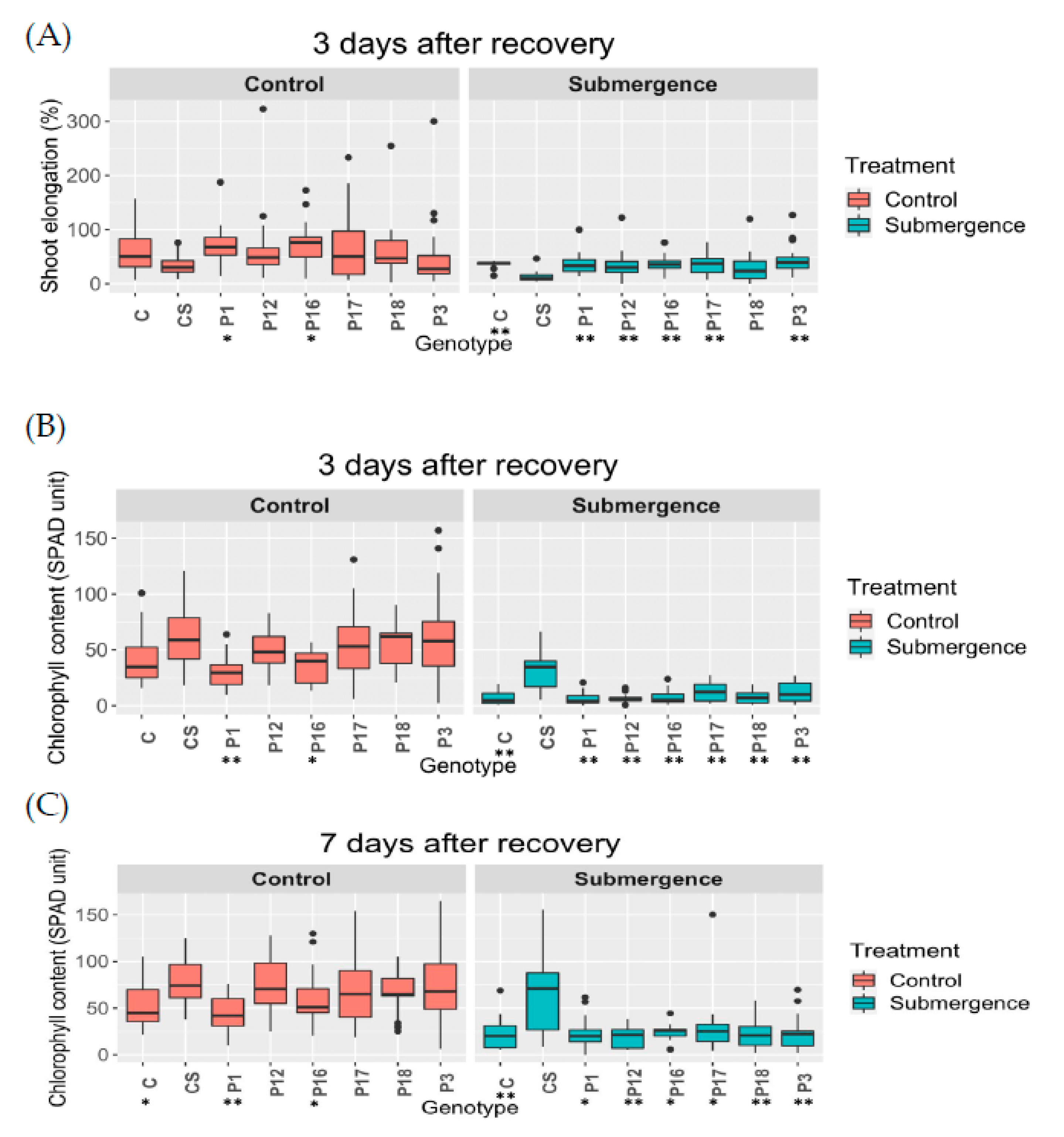
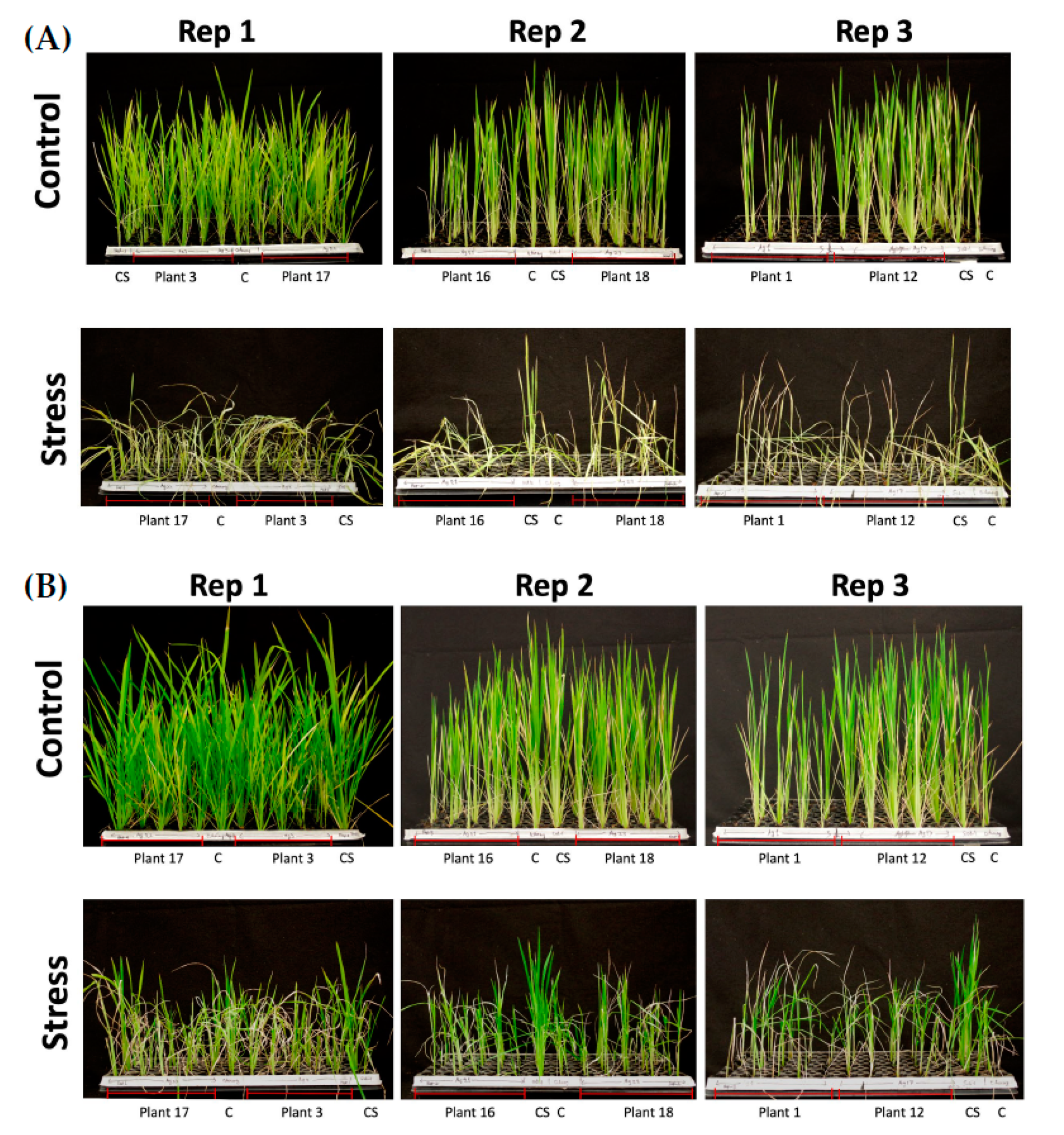
2.7. Phenotyping of T1 Transgenic Plants
3. Materials and Methods
3.1. Plant Materials
3.2. Construction of the CRISPR-Cas9 Vector
3.3. Immature Embryo Harvest, Tissue Culture, and Transformation
3.4. Callus Induction from Mature Seeds for Bombardment
3.5. Plasmid Preparation and Biolistic Bombardment
3.6. Callus Regeneration for Both Immature Embryos and Mature Seeds
3.7. Validation of Transgenic Plants
3.8. Calculation of Regeneration and Transformation Efficiency
3.9. Submergence Experiment and Phenotyping of T1 Plants
3.10. Phenotypic Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2:4-D 6BA CIM CRISPR-Cas9 | 2,4-Dichlorophenoxyacetic acid 6-Benzylaminopurine Callus induction medium Clustered regularly interspaced short palindromic repeats-Cas9 |
| gRNA | Guide RNA |
| HR | Homologous recombination |
| MS NAA NEHJ RIM SIM SUB1 | Murashige and Skoog 1-Naphthaleneacetic acid Non-homologous end-joining Root induction medium Shoot induction medium SUBMERGENCE 1 |
| WT | Wild type |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Formula (1) [19] | Formula (2) [22] | Formula (3) | Formula (4) | Formula (5) |
|---|---|---|---|---|---|
| 2,4-D (2,4-Dichlorophenoxyacetic acid) | 3 mg/L | 2.5 mg/L | 3 mg/L | 3 mg/L | 3 mg/L |
| 6BA (6-Benzylaminopurine) | 0.25 mg/L | 0.15 mg/L | 0.2 mg/L | 0.2 mg/L | 0.3 mg/L |
| Phytagel | 3 g/L | 4 g/L | 3 g/L | 4 g/L | 3 g/L |
| Agarose | - | 2 g/L | - | - | - |
| Components | Formula (1) [19] | Formula (2) | Formula (3) |
|---|---|---|---|
| Kinetin | 2 mg/L | 3 mg/L | 3 mg/L |
| NAA (1-Naphthaleneacetic acid) | 0.2 mg/L | 0.1 mg/L | 0.1 mg/L |
| Agarose | 8 g/L | 8 g/L | 7 g/L |
References
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous Editing of Three Homoeoalleles in Hexaploid Bread Wheat Confers Heritable Resistance to Powdery Mildew. Nat. Biotechnol. 2014, 32, 947. Available online: https://www.nature.com/articles/nbt.2969#supplementary-information (accessed on 29 June 2017). [CrossRef] [PubMed]
- Shan, Q.; Wang, Y.; Li, J.; Gao, C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat. Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doll, N.M.; Gilles, L.M.; Gérentes, M.-F.; Richard, C.; Just, J.; Fierlej, Y.; Borrelli, V.M.; Gendrot, G.; Ingram, G.C.; Rogowsky, P.M. Single and multiple gene knockouts by CRISPR–Cas9 in maize. Plant Cell Rep. 2019, 38, 487–501. [Google Scholar] [CrossRef]
- Qi, W.; Zhu, T.; Tian, Z.; Li, C.; Zhang, W.; Song, R. High-efficiency CRISPR/Cas9 multiplex gene editing using the glycine tRNA-processing system-based strategy in maize. BMC Biotechnol. 2016, 16, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, D.; Chen, M.; Liang, W.; Wei, J.; Qi, Y.; Yuan, Z. Development of japonica photo-sensitive genic male sterile rice lines by editing carbon starved anther using CRISPR/Cas9. J. Genet. Genom. 2016. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Guo, D.; Zhang, J.; Huang, Q.; Qin, G.; Zhang, X.; Wan, J.; Gu, H.; Qu, L.-J. Targeted mutagenesis in rice using CRISPR-Cas system. Cell Res. 2013, 23, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Biswal, A.K.; Dionora, J.; Perdigon, K.M.; Balahadia, C.P.; Mazumdar, S.; Chater, C.; Lin, H.-C.; Coe, R.A.; Kretzschmar, T. CRISPR-Cas9 and CRISPR-Cpf1 mediated targeting of a stomatal developmental gene EPFL9 in rice. Plant Cell Rep. 2017, 36, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Voytas, D.F.; Hsieh, T.-F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.W., 3rd; Qi, Y. CRISPR/Cas9 for plant genome editing: Accomplishments, problems and prospects. Plant Cell Rep. 2016, 35, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Gelvin, S.B. Agrobacterium-Mediated Plant Transformation: The Biology behind the “Gene-Jockeying” Tool. Microbiol. Mol. Biol. Rev. 2003, 67, 16. [Google Scholar] [CrossRef] [Green Version]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Aldemita, R.R.; Hodges, T.K. Agrobacterium tumefaciens-mediated transformation of japonica and indica rice varieties. Planta 1996, 199, 612–617. [Google Scholar] [CrossRef]
- Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed. Nat. Protoc. 2008, 3, 824. [Google Scholar] [CrossRef] [PubMed]
- Slamet-Loedin, I.H.; Chadha-Mohanty, P.; Torrizo, L. Agrobacterium-mediated transformation: Rice transformation. In Cereal Genomics; Henry, R., Furtado, A., Eds.; Methods in Molecular Biology (Methods and Protocols); Humana Press: Totowa, NJ, USA, 2014; Volume 1099, pp. 261–271. [Google Scholar] [CrossRef]
- Kumar, K.K.; Maruthasalam, S.; Loganathan, M.; Sudhakar, D.; Balasubramanian, P. An improvedAgrobacterium-mediated transformation protocol for recalcitrant elite indica rice cultivars. Plant Mol. Biol. Rep. 2005, 23, 67–73. [Google Scholar] [CrossRef]
- Lin, Y.J.; Zhang, Q. Optimising the tissue culture conditions for high efficiency transformation of indica rice. Plant Cell Rep. 2005, 23, 540–547. [Google Scholar] [CrossRef]
- Sahoo, K.K.; Tripathi, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. An improved protocol for efficient transformation and regeneration of diverse indica rice cultivars. Plant Methods 2011, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Hiei, Y.; Komari, T. Improved protocols for transformation of indica rice mediated by Agrobacterium tumefaciens. Plant Cell Tissue Organ Cult. 2006, 85, 271–283. [Google Scholar] [CrossRef]
- Zaidi, M.; Narayanan, M.; Sardana, R.; Taga, I.; Postel, S.; Johns, R.; McNulty, M.; Mottiar, Y.; Mao, J.; Loit, E. Optimizing tissue culture media for efficient transformation of different indica rice genotypes. Agron. Res. 2006, 4, 563–575. [Google Scholar]
- Sahoo, R.K.; Tuteja, N. Development of Agrobacterium-mediated transformation technology for mature seed-derived callus tissues of indica rice cultivar IR64. GM Crops Food 2012, 3, 123–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Septiningsih, E.M.; Hidayatun, N.; Sanchez, D.L.; Nugraha, Y.; Carandang, J.; Pamplona, A.M.; Collard, B.C.; Ismail, A.M.; Mackill, D.J. Accelerating the development of new submergence tolerant rice varieties: The case of Ciherang-Sub1 and PSB Rc18-Sub1. Euphytica 2015, 202, 259–268. [Google Scholar] [CrossRef]
- Toledo, A.M.U.; Ignacio, J.C.I.; Casal, C., Jr.; Gonzaga, Z.J.; Mendioro, M.S.; Septiningsih, E.M. Development of Improved Ciherang-Sub1 Having Tolerance to Anaerobic Germination Conditions. Plant Breed. Biotechnol. 2015, 32, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Gonzaga, Z.J.C.; Carandang, J.; Singh, A.; Collard, B.C.Y.; Thomson, M.J.; Septiningsih, E.M. Mapping QTLs for submergence tolerance in rice using a population fixed for SUB1A tolerant allele. Mol. Breed. 2017, 37, 47. [Google Scholar] [CrossRef]
- Singh, A.; Carandang, J.; Gonzaga, Z.J.C.; Collard, B.C.Y.; Ismail, A.M.; Septiningsih, E.M. Identification of QTLs for yield and agronomic traits in rice under stagnant flooding conditions. Rice 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pramudyawardani, E.F.; Aswidinnoor, H.; Purwoko, B.S.; Suwarno, W.B.; Islam, M.; Verdeprado, H.; Collard, B.C. Genetic analysis and QTL mapping for agronomic and yield-related traits in Ciherang-Sub1 rice backcross populations. Plant Breed. Biotechnol. 2018, 6, 177–192. [Google Scholar] [CrossRef]
- Septiningsih, E.M.; Mackill, D.J. Genetics and breeding of flooding tolerance in rice. In Rice Genomics, Genetics and Breeding; Sasaki, T., Ashikari, M., Eds.; Springer: Singapore, 2018; pp. 275–295. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor–like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Septiningsih, E.M.; Pamplona, A.M.; Sanchez, D.L.; Neeraja, C.N.; Vergara, G.V.; Heuer, S.; Ismail, A.M.; Mackill, D.J. Development of submergence-tolerant rice cultivars: The Sub1 locus and beyond. Ann. Bot. 2009, 103, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Voesenek, L.A.; Bailey-Serres, J. Flood adaptive traits and processes: An overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef]
- Mendoza, M.G.; Kaeppler, H.F. Auxin and sugar effects on callus induction and plant regeneration frequencies from mature embryos of wheat (Triticum aestivum L.). Cell. Dev. Biol. Plant 2002, 38, 39–45. [Google Scholar] [CrossRef]
- de Paiva Neto, V.B.; Otoni, W.C. Carbon sources and their osmotic potential in plant tissue culture: Does it matter? Sci. Hortic. 2003, 97, 193–202. [Google Scholar] [CrossRef]
- Orshinsky, B.R.; McGregor, L.J.; Johnson, G.I.E.; Hucl, P.; Kartha, K.K. Improved embryoid induction and green shoot regeneration from wheat anthers cultured in medium with maltose. Plant Cell Rep. 1990, 9, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Skoog, F.; Miller, C. Chemical regulation of growth and organ formation in plant tissues cultured. Symp. Soc. Exp. Biol. 1957, 11, 118–131. [Google Scholar] [PubMed]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell. Dev. Biol.-Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Skoog, F.; Tsui, C. Chemical Control of Growth and Bud Formation in Tobacco Stem Segments and Callus Cultured In Vitro. Am. J. Bot. 1948, 35, 782–787. [Google Scholar] [CrossRef]
- Deb, C.; Imchen, T. An Efficient In vitro Hardening Technique of Tissue Culture Raised Plants. Biotechnology 2010, 9, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Hauptmann, R.M.; Widholm, J.M.; Paxton, J.D. Benomyl: A broad spectrum fungicide for use in plant cell and protoplast culture. Plant Cell Rep. 1985, 4, 129–132. [Google Scholar] [CrossRef]
- Thielges, B.A.; Hoitink, H.A.J. Fungicides Aid Rooting of Eastern White Pine Cuttings. For. Sci. 1972, 18, 54–55. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Zhang, Y.; Liu, J.; Yin, K.; Qiu, J.L.; Gao, C. Genome editing of bread wheat using biolistic delivery of CRISPR/Cas9 in vitro transcripts or ribonucleoproteins. Nat. Protoc. 2018, 13, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bono, H.; Hino, K.; Ui-Tei, K.; Naito, Y. CRISPRdirect: Software for Designing CRISPR/Cas Guide RNA with Reduced Off-Target Sites. Bioinformatics 2014, 31, 1120–1123. [Google Scholar] [CrossRef]
- Biolistic® PDS-1000/He Particle Delivery System User Manual. Available online: http://www.bio-rad.com/webroot/web/pdf/lsr/literature/M1652249.pdf (accessed on 8 June 2021).
- Liang, Y.; Baring, M.; Wang, S.; Septiningsih, E.M. Mapping QTLs for Leafspot Resistance in Peanut Using SNP-Based Next-Generation Sequencing Markers. Plant Breed. Biotechnol. 2017, 5, 115–122. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Biswas, S.; Kim, B.; Bailey-Serres, J.; Septiningsih, E.M. Improved Transformation and Regeneration of Indica Rice: Disruption of SUB1A as a Test Case via CRISPR-Cas9. Int. J. Mol. Sci. 2021, 22, 6989. https://doi.org/10.3390/ijms22136989
Liang Y, Biswas S, Kim B, Bailey-Serres J, Septiningsih EM. Improved Transformation and Regeneration of Indica Rice: Disruption of SUB1A as a Test Case via CRISPR-Cas9. International Journal of Molecular Sciences. 2021; 22(13):6989. https://doi.org/10.3390/ijms22136989
Chicago/Turabian StyleLiang, Yuya, Sudip Biswas, Backki Kim, Julia Bailey-Serres, and Endang M. Septiningsih. 2021. "Improved Transformation and Regeneration of Indica Rice: Disruption of SUB1A as a Test Case via CRISPR-Cas9" International Journal of Molecular Sciences 22, no. 13: 6989. https://doi.org/10.3390/ijms22136989