
Impaired Morphogenesis and Function of Rat Adrenal Zona Glomerulosa by Developmental Low-Dose Exposure to DDT Is Associated with Altered Oct4 Expression

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
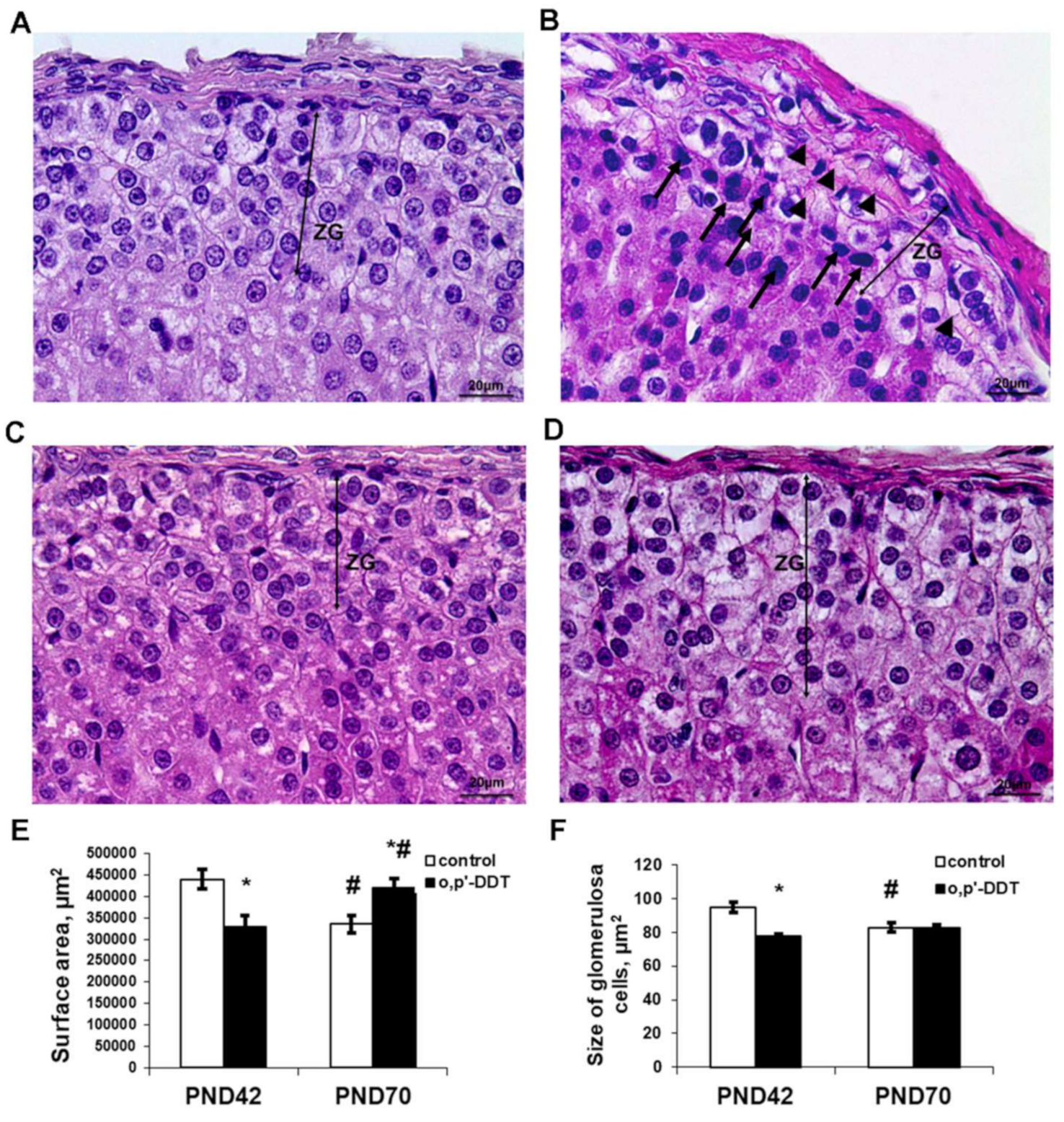
2.1. Histology of the Zona Glomerulosa of the Adrenal Cortex

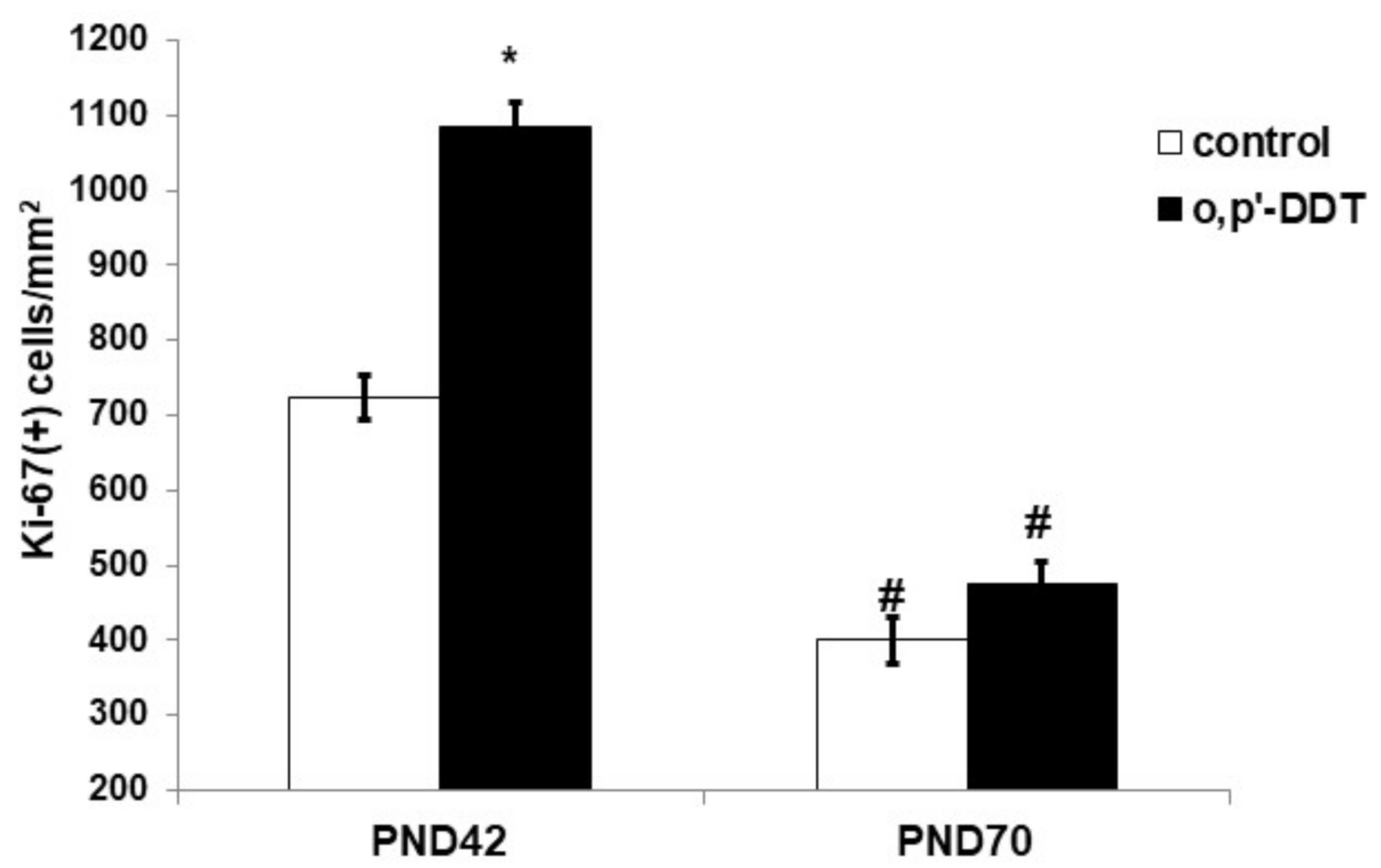
2.2. Proliferative Activity of Glomerulosa Cells
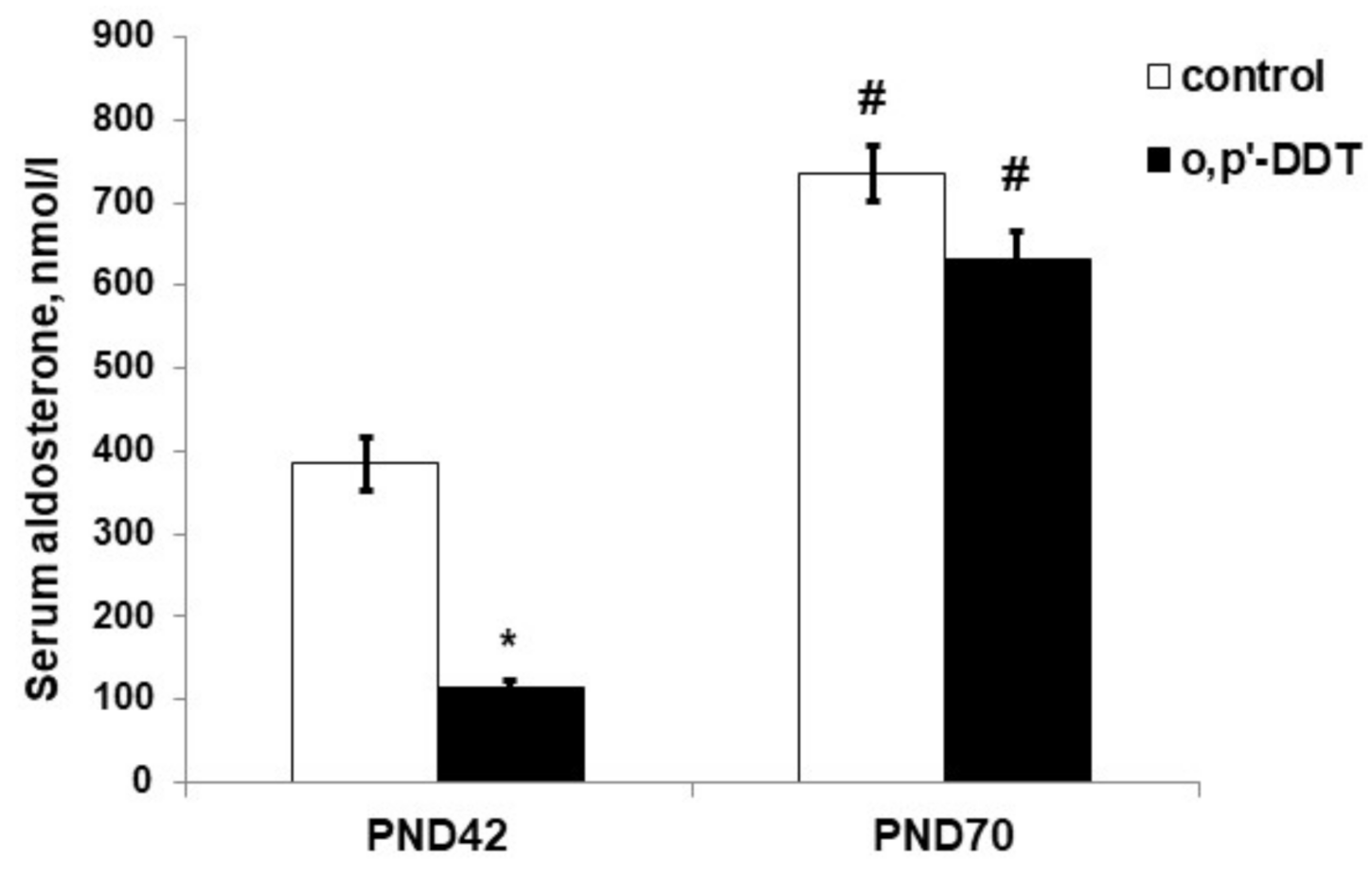
2.3. Aldosterone Production
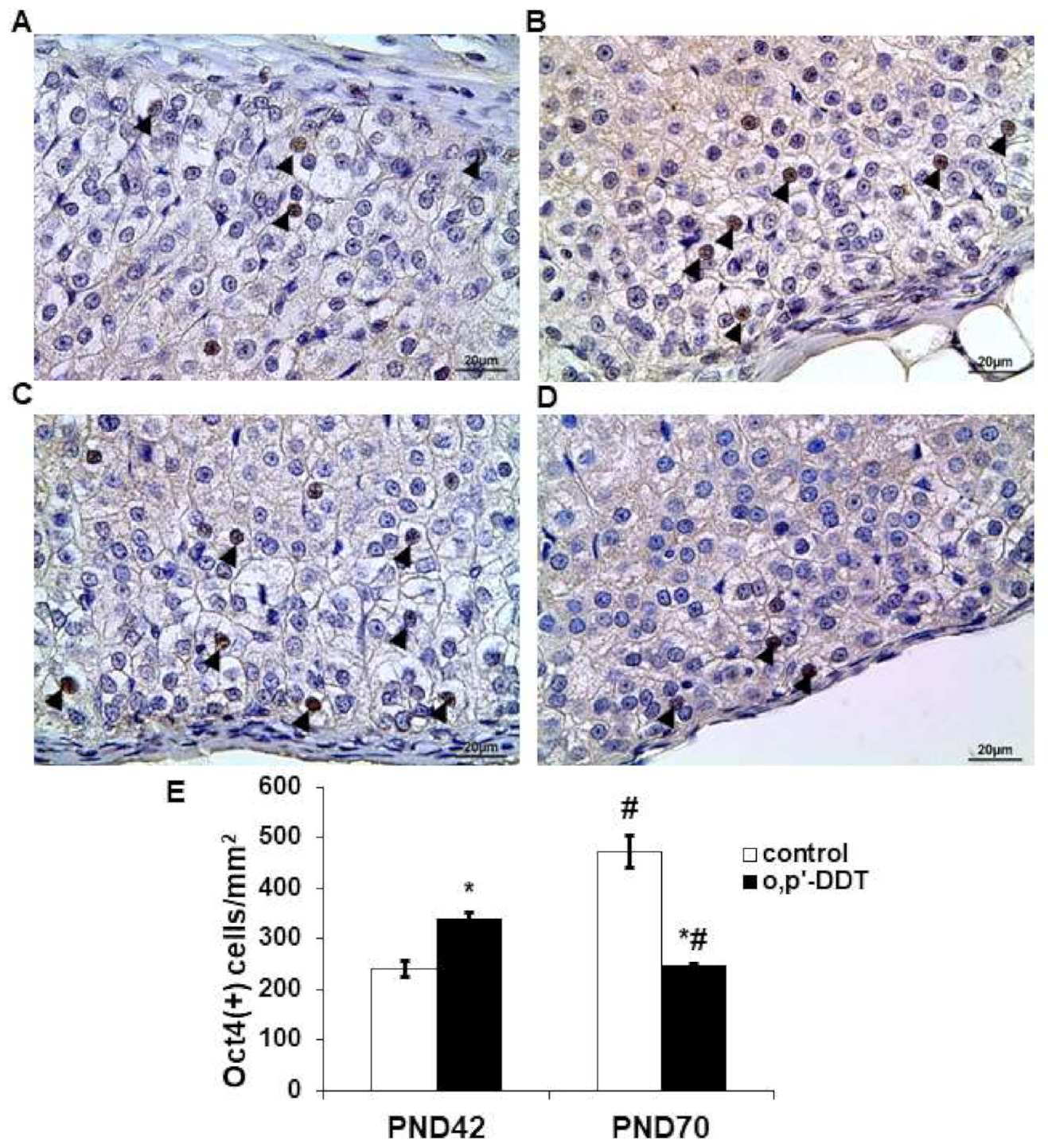
2.4. Expression of Transcriptional Factor Oct4

3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Adrenal Histology
4.4. Immunohistochemistry
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zoeller, R.T.; Brown, T.R.; Doan, L.L.; Gore, A.C.; Skakkebaek, N.E.; Soto, A.M.; Woodruff, T.J.; Vom Saal, F.S. Endocrine disrupting chemicals and public health protection: A statement of principles from The Endocrine Society. Endocrinology 2012, 153, 4097–4110. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef] [PubMed]
- Calsolaro, V.; Pasqualetti, G.; Niccolai, F.; Caraccio, N.; Monzani, F. Thyroid disrupting chemicals. Int. J. Mol. Sci. 2017, 18, 2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: Effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Street, M.E.; Angelini, S.; Bernasconi, S.; Burgio, E.; Cassio, A.; Catellani, C.; Cirillo, F.; Deodati, A.; Fabbrizi, E.; Fanos, V.; et al. Current knowledge on endocrine disrupting chemicals (EDCs) from animal biology to humans, from pregnancy to adulthood: Highlights from a national Italian meeting. Int. J. Mol. Sci. 2018, 19, 1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papalou, O.; Kandaraki, E.; Papadakis, G.; Diamanti-Kandarakis, E. Endocrine disrupting chemicals: An occult mediator of metabolic disease. Front. Endocrinol. 2019, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2, The Endocrine Society’s second scientific statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- La Merrill, M.A.; Vandenberg, L.N.; Smith, M.T.; Goodson, W.; Browne, P.; Patisaul, H.B.; Guyton, K.Z.; Kortenkamp, A.; Cogliano, V.J.; Woodruff, T.J.; et al. Consensus on the key characteristics of endocrine-disrupting chemicals as a basis for hazard identification. Nat. Rev. Endocrinol. 2020, 18, 45–57. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Pesticide Residues in Food—2018. Toxicological Evaluations; World Health Organization and Food and Agriculture Organization of the United Nations; WHO: Geneva, Switzerland, 2019; 780p. [Google Scholar]
- World Health Organization. Pesticide Residues in Food—2016 Evaluations. Part II—Toxicological; WHO: Geneva, Switzerland, 2016; 480p. [Google Scholar]
- Spear, R. Recognized and possible exposure to pesticide. In Handbook of Pesticide Toxicology; Hayes, W.J., Jr., Edwards, R.L., Jr., Eds.; Academic Press: New York, NY, USA, 1999; pp. 245–274. [Google Scholar]
- Jaga, K.; Dharmani, C. Global surveillance of DDT and DDE levels in human tissues. Int. J. Occup. Med. Environ. Health 2003, 16, 7–20. [Google Scholar] [CrossRef]
- Kelce, W.R.; Stone, C.R.; Laws, S.C.; Gray, L.E.; Kemppainen, J.A.; Wilson, E.M. Persistent DDT metabolite p,p’-DDE is a potent androgen receptor antagonist. Nature 1995, 375, 581–585. [Google Scholar] [CrossRef]
- Kwekel, J.C.; Forgacs, A.L.; Williams, K.J.; Zacharewski, T.R. o,p’-DDT-mediated uterotrophy and gene expression in immature C57BL/6 mice and Sprague-Dawley rats. Toxicol. Appl. Pharmacol. 2013, 273, 532–541. [Google Scholar] [CrossRef]
- Damgaard, I.N.; Skakkebaek, N.E.; Toppari, J.; Virtanen, H.E.; Shen, H.; Schramm, K.W.; Petersen, J.H.; Jensen, T.K.; Main, K.M. Persistent pesticides in human breast milk and cryptorchidism. Environ. Health Perspect. 2006, 114, 1133–1138. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, S.L.; Herring, A.H.; Sjodin, A.; Jones, R.; Needham, L.; Ma, C.; Ding, K.; Shaw, G.M. Hypospadias and halogenated organic pollutant levels in maternal mid-pregnancy serum samples. Chemosphere 2010, 80, 641–646. [Google Scholar] [CrossRef] [Green Version]
- Kalfa, N.; Philibert, P.; Baskin, L.S.; Sultan, C. Hypospadias: Interactions between environment and genetics. Mol. Cell. Endocrinol. 2011, 335, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Ozen, S.; Goksen, D.; Darcan, S. Agricultural pesticides and precocious puberty. Vitam. Horm. 2014, 94, 27–40. [Google Scholar] [CrossRef]
- Freire, C.; Koifman, R.J.; Sarcinelli, P.N.; Simoes Rosa, A.C.; Clapauch, R.; Koifman, S. Long-term exposure to organochlorine pesticides and thyroid status in adults in a heavily contaminated area in Brazil. Environ. Res. 2013, 127, 7–15. [Google Scholar] [CrossRef]
- Alsen, M.; Sinclair, C.; Cooke, P.; Ziadkhanpour, K.; Genden, E.; van Gerven, M. Endocrine Disrupting Chemicals and Thyroid Cancer: An Overview. Toxics 2021, 9, 14. [Google Scholar] [CrossRef]
- Martinez-Arguelles, D.; Papadopoulos, V. Mechanisms mediating environmental chemical-induced endocrine disruption in the adrenal gland. Front. Endocrinol. 2015, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Helsley, R.; Zhou, C. Epigenetic impact of endocrine disrupting chemicals on lipid homeostasis and atherosclerosis: A pregnane X receptor-centric view. Environ. Epigenet. 2017, 3, dvx017. [Google Scholar] [CrossRef]
- Alavian-Ghavanini, A.; Ruegg, J. Understanding epigenetic effects of endocrine disrupting chemicals: From mechanisms to novel test methods. Basic Clin. Pharmacol. Toxicol. 2018, 122, 38–45. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Tsomartova, D.A.; Obernikhin, S.S.; Nazimova, S.V. The role of the canonical Wnt-signaling pathway in morphogenesis and regeneration of the adrenal cortex in rats exposed to the endocrine disruptor dichlorodiphenyltrichloroethane during prenatal and postnatal development. Biol. Bull. 2019, 46, 74–81. [Google Scholar] [CrossRef]
- Mariniello, K.; Ruiz-Babot, G.; McGaugh, E.C.; Nicholson, J.G.; Gualtieri, A.; Gaston-Massuet, C.; Nostro, M.C.; Guasti, L. Stem cells, self-renewal, and lineage commitment in the endocrine system. Front. Endocrinol. 2019, 10, 772. [Google Scholar] [CrossRef]
- Olariu, V.; Lövkvist, C.; Sneppen, K. Nanog, Oct4 and Tet1 interplay in establishing pluripotency. Sci. Rep. 2016, 6, 25438. [Google Scholar] [CrossRef]
- Liu, X.; Huang, J.; Chen, T.; Wang, Y.; Xin, S.; Li, J.; Pei, G.; Kang, J. Yamanaka factors critically regulate the developmental signaling network in mouse embryonic stem cells. Cell Res. 2008, 18, 1177–1189. [Google Scholar] [CrossRef]
- Zangrossi, S.; Marabese, M.; Broggini, M.; Giordano, R.; D’Erasmo, M.; Montelatici, E.; Intini, D.; Neri, A.; Pesce, M.; Rebulla, P.; et al. Oct-4 expression in adult human differentiated cells challenges its role as a pure stem cell marker. Stem Cells 2007, 25, 1675–1680. [Google Scholar] [CrossRef]
- Alexander, R.; Cheng, L.; Grignon, D.; Indrees, M. Cytoplasmic OCT4 staining is a sensitive marker of neuroendocrine differentiation. Hum. Pathol. 2014, 45, 27–32. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Obernikhin, S.S.; Nazimova, S.V.; Yaglov, V.V. Role of transcription factor Oct4 in postnatal development and function of the adrenal cortex. Bull. Exp. Biol. Med. 2019, 167, 568–573. [Google Scholar] [CrossRef]
- Obernikhin, S.S.; Yaglova, N.V.; Nazimova, S.V.; Yaglov, V.V.; Timokhina, E.P. Transcriptional regulation of morphogenesis of rat adrenal zona glomerulosa during postnatal development. Clin. Exp. Morphol. 2019, 167, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, D.; Pinto, P.; Magalhaes, M.M.; Magalhaes, M.C. The development of the adrenal gland zona glomerulosa in the rat. A morphological, immunohistochemical and biochemical study. Mol. Cell. Endocrinol. 1998, 140, 163–168. [Google Scholar] [CrossRef]
- Lengner, C.; Camargo, F.; Hochedlinger, K.; Welstead, G.; Zaidi, S.; Okhale, S.; Scholer, H.; Tomilin, A.; Jaenisch, R. Oct4 expression is not required for mouse somatic stem cell self-renewal. Cell Stem Cell 2007, 1, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Yaglova, N.V.; Obernikhin, S.S.; Nazimova, S.V.; Yaglov, V.V. Developmental exposure to endocrine disrupter dichlorodiphenyltrichloroethane alters transcriptional regulation of postnatal morphogenesis of adrenal zona fasciculata. Saudi J. Biol. Sci. 2020, 27, 3655–3659. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mukhopadhyay, P.; Pisano, M.M.; Lu, X.; Huang, L.; Lu, Q.; Dean, D.C. Repression of Zeb1 and hypoxia cause sequential MET and induction of Aid, Oct4, and Dnmt1, leading to immortalization and multipotential reprogramming of fibroblasts in spheres. Stem Cells 2013, 31, 1350–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-Q. Networks of transcription factors for Oct4 expression in mice. DNA Cell Biol. 2017, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bruder, E.; Nagler, A.; Raff, H. Oxygen-dependence of ACTH-stimulated aldosterone and corticosterone synthesis in the rat adrenal cortex: Developmental aspects. J. Endocrinol. 2002, 172, 595–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorente, M.; Mirapeux, R.; Miguel, M.; Longmei, W.; Volk, D.; Cervos-Navarro, J. Chronic hypoxia induced ultrastructural changes in the rat adrenal zona glomerulosa. Histol. Histopathol. 2002, 17, 185–190. [Google Scholar] [CrossRef]
- Mitani, F. Functional zonation of the rat adrenal cortex: The development and maintenance. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2014, 90, 163–183. [Google Scholar] [CrossRef] [Green Version]
- Pignatti, E.; Leng, S.; Yuchi, Y.; Borges, K.; Guagliardo, N.; Shan, M.; Ruiz-Babot, G.; Katyiawasam, D.; Taketo, M.; Mial, J.; et al. Beta-catenin causes adrenal hyperplasia by blocking zonal transdifferentiation. Cell Rep. 2020, 31, 107524. [Google Scholar] [CrossRef]
- Pascual, A.; Aranda, A. Thyroid hormone receptors, cell growth and differentiation. Biochem. Biophys. Acta 2013, 1830, 3908–3916. [Google Scholar] [CrossRef]
- Blitz, E.; Matsuda, H.; Guenther, S.; Morikawa, T.; Kubota, Y.; Zada, D.; Lerer-Goldshtein, T.; Stainier, D.; Appelbaum, L. Thyroid hormones regulate goblet cell differentiation and Fgf19-Fgfr4 signaling. Endocrinology 2021, 162, bqab047. [Google Scholar] [CrossRef]
- Gaspar, F.W.; Harley, K.G.; Kogut, K.; Chevrier, J.; Mora, A.M.; Sjödin, A.; Ezkenasi, B. Prenatal DDT and DDE exposure and child IQ in the CHAMACOS cohort. Environ. Int. 2015, 85, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Mariano, J.A.; Torres-Sanchez, L.; Bassol-Mayagoitia, S.; Escamilla-Nunez, M.C.; Cebrian, M.E.; Villeda-Gutierrez, E.A.; Lopez-Rodriguez, G.; Felix-Arellano, E.E.; Blanco-Munoz, J. Effect of exposure to p,p-DDE during the first half of pregnancy in the maternal thyroid profile of female residents in a Mexican floriculture area. Environ. Res. 2017, 156, 597–604. [Google Scholar] [CrossRef]
- Leemans, M.; Couderq, S.; Demeneix, B.; Fini, J.-B. Pesticides with potential thyroid hormone-disrupting effects: A review of recent data. Front. Endocrinol. 2019, 10, 743. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, D.; Xiao, F.; Gouvtia, A.; Ferreira, J.; Vinson, G. Adrenarche in the rat. J. Endocrinol. 2006, 191, 301–308. [Google Scholar] [CrossRef]
- Yamazaki, H.; Takano, R.; Shimizu, M.; Murayama, N.; Kitajima, M.; Shono, M. Human blood concentrations of dichlorodiphenyltrichloroethane (DDT) extrapolated from metabolism in rats and humans and physiologically based pharmacokinetic modeling. J. Health Sci. 2010, 56, 566–575. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaglova, N.V.; Obernikhin, S.S.; Tsomartova, D.A.; Nazimova, S.V.; Yaglov, V.V.; Tsomartova, E.S.; Chereshneva, E.V.; Ivanova, M.Y.; Lomanovskaya, T.A. Impaired Morphogenesis and Function of Rat Adrenal Zona Glomerulosa by Developmental Low-Dose Exposure to DDT Is Associated with Altered Oct4 Expression. Int. J. Mol. Sci. 2021, 22, 6324. https://doi.org/10.3390/ijms22126324
Yaglova NV, Obernikhin SS, Tsomartova DA, Nazimova SV, Yaglov VV, Tsomartova ES, Chereshneva EV, Ivanova MY, Lomanovskaya TA. Impaired Morphogenesis and Function of Rat Adrenal Zona Glomerulosa by Developmental Low-Dose Exposure to DDT Is Associated with Altered Oct4 Expression. International Journal of Molecular Sciences. 2021; 22(12):6324. https://doi.org/10.3390/ijms22126324
Chicago/Turabian StyleYaglova, Nataliya V., Sergey S. Obernikhin, Dibakhan A. Tsomartova, Svetlana V. Nazimova, Valentin V. Yaglov, Elina S. Tsomartova, Elizaveta V. Chereshneva, Marina Y. Ivanova, and Tatiana A. Lomanovskaya. 2021. "Impaired Morphogenesis and Function of Rat Adrenal Zona Glomerulosa by Developmental Low-Dose Exposure to DDT Is Associated with Altered Oct4 Expression" International Journal of Molecular Sciences 22, no. 12: 6324. https://doi.org/10.3390/ijms22126324