
Diabetes Affects the Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)-Like Immunoreactive Enteric Neurons in the Porcine Digestive Tract


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
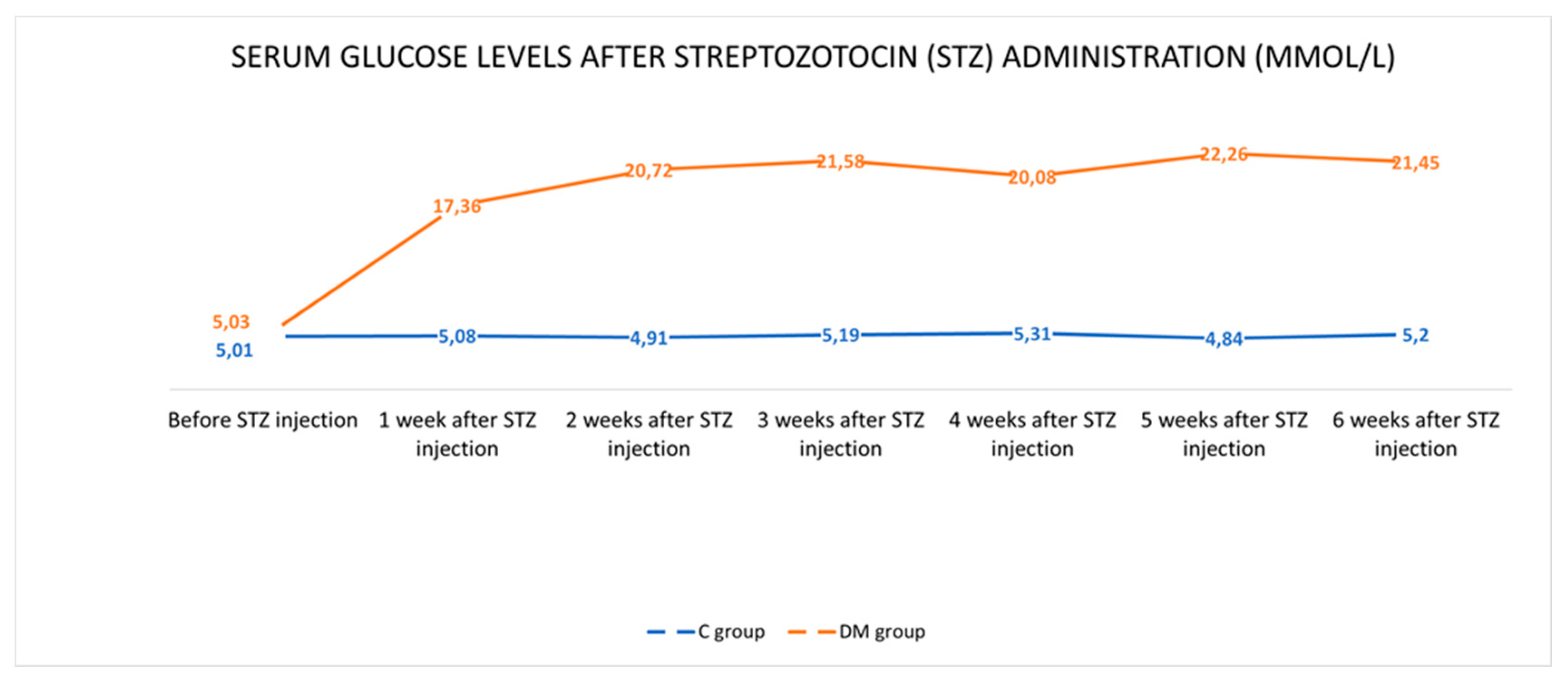
2.1. Control of Glycaemia
2.2. Immunofluorescence Technique
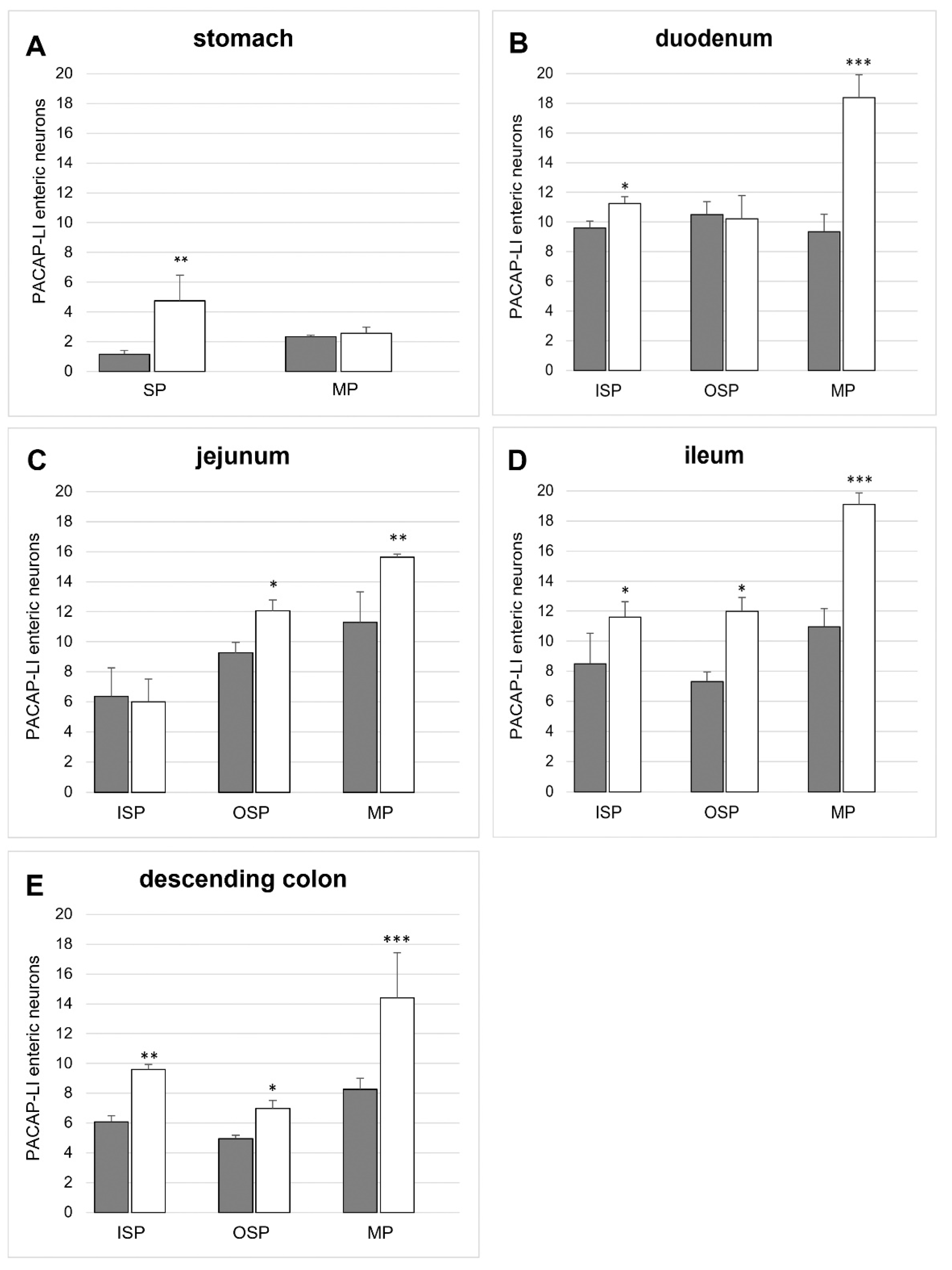
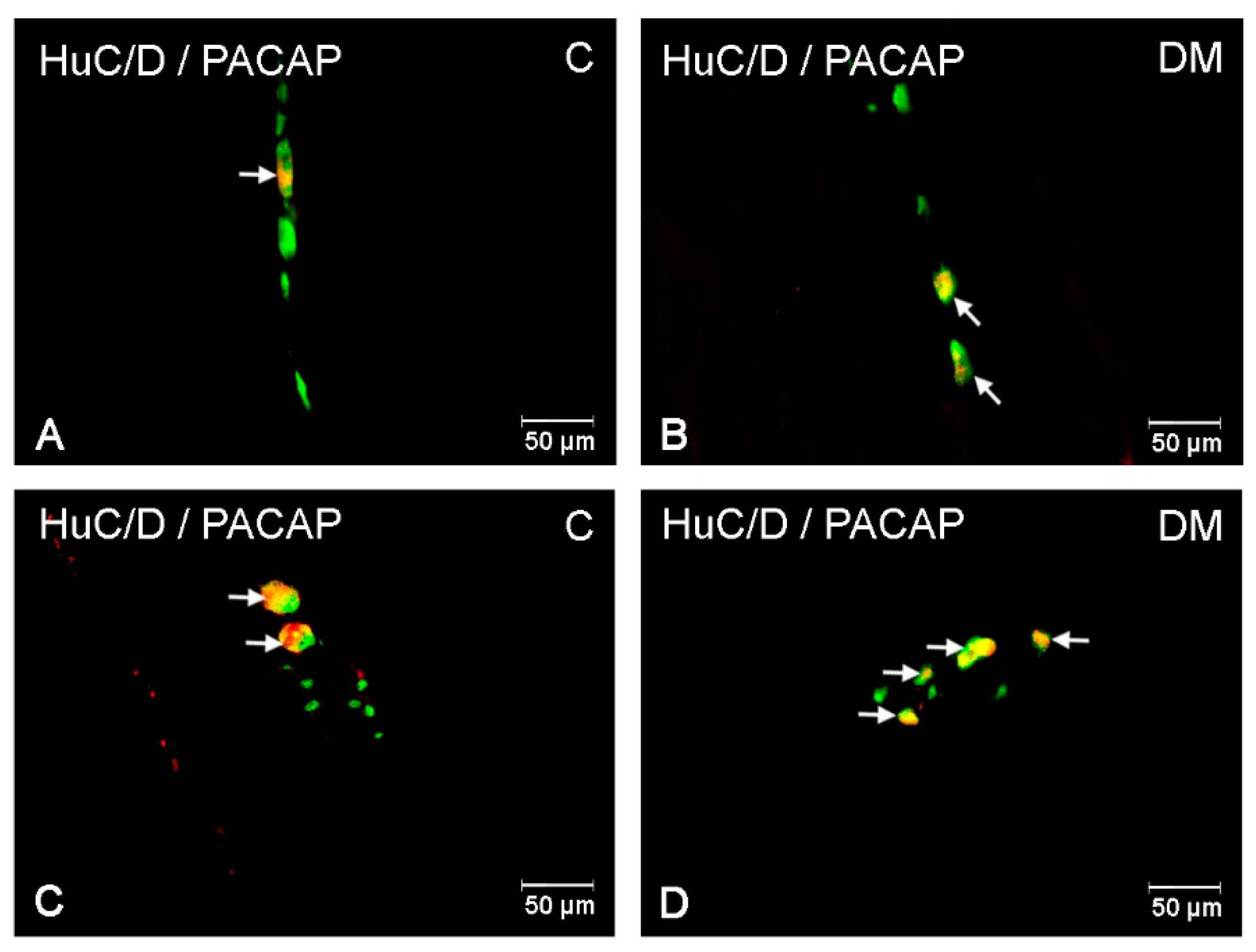
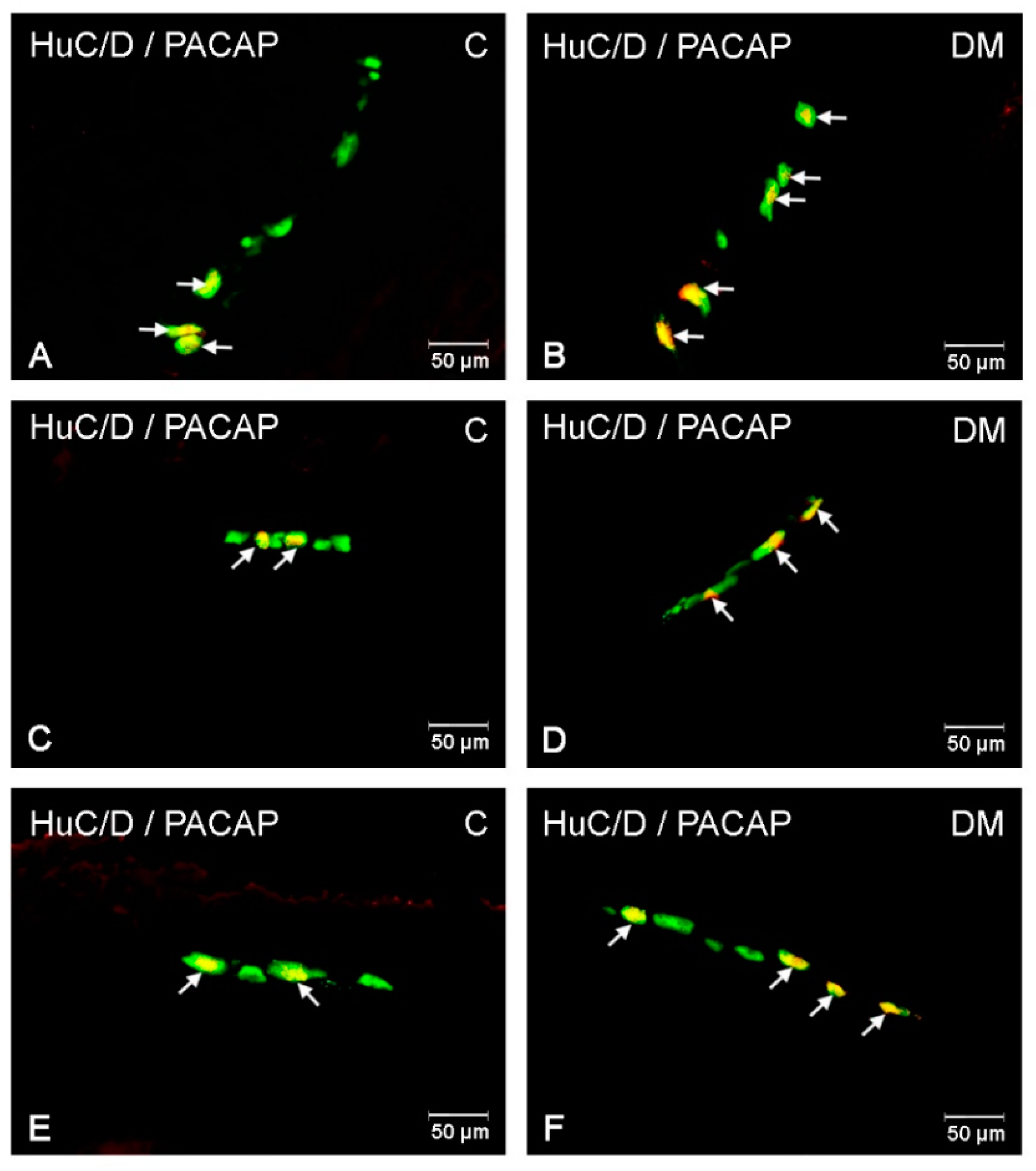
2.2.1. PACAP-LI Neurons in the Porcine Corpus of the Stomach
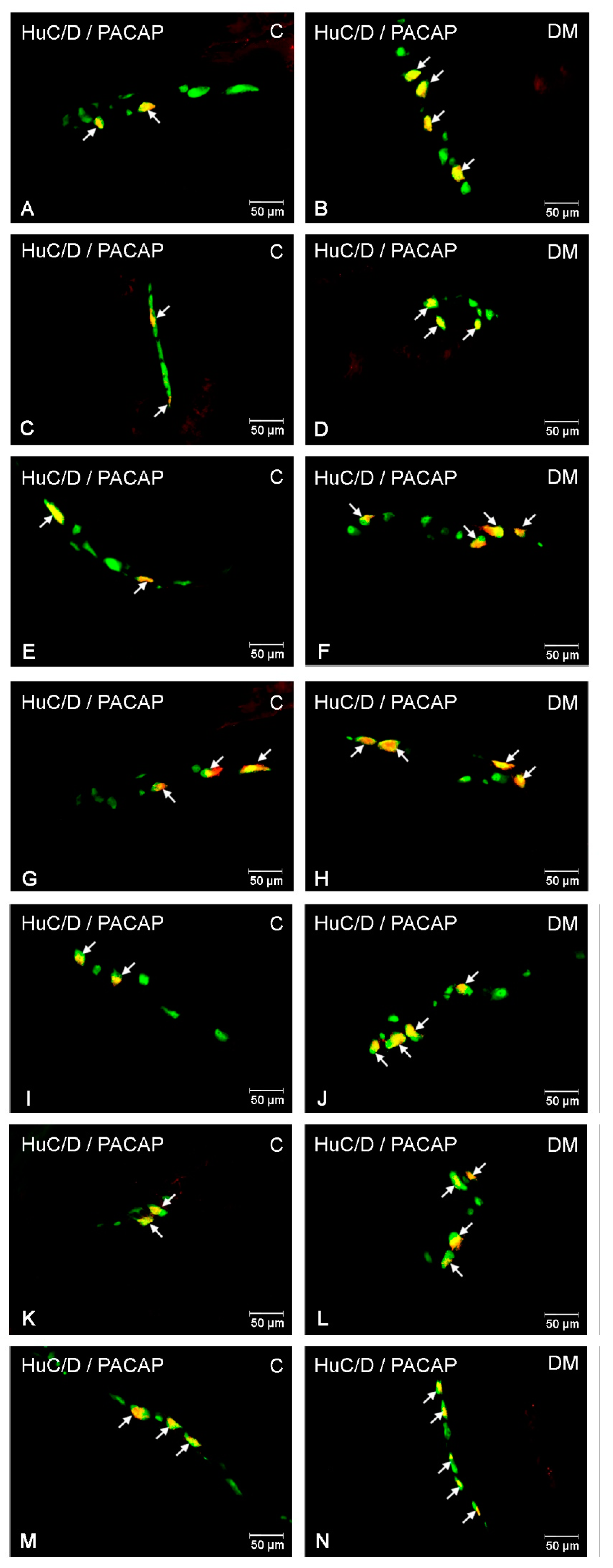
2.2.2. PACAP-LI Neurons in the Porcine Small Intestine
2.2.3. PACAP-LI Neuron in the Porcine Descending Colon
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017. [Google Scholar]
- Kurniawan, A.H.; Suwandi, B.H.; Kholili, U. Diabetic Gastroenteropathy: A Complication of Diabetes Mellitus. Acta Med. Indones 2019, 51, 263–271. [Google Scholar]
- Kornum, D.S.; Klinge, M.W.; Fassov, J.; Brock, C.; Bjerregård, N.C.; Lund, S.; Drewes, A.M.; Krogh, K. Symptoms of diabetic gastroenteropathy in patients with diabetes. Ugeskr. Laeger 2020, 182, V6200462. [Google Scholar]
- Callaghan, B.C.; Little, A.A.; Feldman, E.L.; Hughes, R.A. Enhanced glucose control for preventing and treating diabetic neuropathy. Cochrane Database Syst. Rev. 2012, 6, CD007543. [Google Scholar] [CrossRef] [PubMed]
- Nezami, B.G.; Srinivasan, S. Enteric nervous system in the small intestine: Pathophysiology and clinical implications. Curr. Gastroenterol. Rep. 2010, 12, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonkowski, S.; Całka, J. Changes in pituitary adenylate cyclase-activating Peptide 27-like immunoreactive nervous structures in the porcine descending colon during selected pathological processes. J. Mol. Neurosci. 2012, 48, 777–787. [Google Scholar] [CrossRef]
- Palus, K.; Bulc, M.; Całka, J. Changes in VIP-, SP- and CGRP- like immunoreactivity in intramural neurons within the pig stomach following supplementation with low and high doses of acrylamide. Neurotoxicology 2018, 69, 47–59. [Google Scholar] [CrossRef]
- Bulc, M.; Palus, K.; Dąbrowski, M.; Całka, J. Hyperglycaemia-induced downregulation in expression of nNOS intramural neurons of the small intestine in the pig. Int. J. Mol. Sci. 2019, 20, 1681. [Google Scholar] [CrossRef] [Green Version]
- Miyata, A.; Jiang, L.; Dahl, R.D.; Kitada, C.; Kubo, K.; Fujino, M.; Minamino, N.; Arimura, A. Isolation of a neuropeptide corresponding to the N-terminal 27 residues of the pituitary adenylate cyclase activating polypeptide with 38 residues (PACAP38). Biochem. Biophys. Res. Commun. 1990, 170, 643–648. [Google Scholar] [CrossRef]
- Horvath, G.; Opper, B.; Reglodi, D. The Neuropeptide Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) is Protective in Inflammation and Oxidative Stress-Induced Damage in the Kidney. Int. J. Mol. Sci. 2019, 20, 4944. [Google Scholar] [CrossRef] [Green Version]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Shen, Z.; Larsson, L.T.; Malmfors, G.; Absood, A.; Håkanson, R.; Sundler, F. A novel neuropeptide, pituitary adenylate cyclase-activating polypeptide (PACAP), in human intestine: Evidence for reduced content in Hirschsprung’s disease. Cell Tissue Res. 1992, 269, 369–374. [Google Scholar] [CrossRef]
- Hannibal, J.; Ekblad, E.; Mulder, H.; Sundler, F.; Fahrenkrug, J. Pituitary adenylate cyclase activating polypeptide (PACAP) in the gastrointestinal tract of the rat: Distribution and effects of capsaicin or denervation. Cell Tissue Res. 1998, 291, 65–79. [Google Scholar] [CrossRef]
- Uddman, R.; Luts, A.; Absood, A.; Arimura, A.; Ekelund, M.; Desai, H.; HÂkanson, R.; Hambreaus, G.; Sundler, F. PACAP, a VIP-like peptide, in neurons of the esophagus. Regul. Pept. 1991, 36, 415–422. [Google Scholar] [CrossRef]
- Sundler, F.; Ekblad, E.; Absood, A.; Håkanson, R.; Köves, K.; Arimura, A. Pituitary adenylate cyclase activating peptide: A novel vasoactive intestinal peptide-like neuropeptide in the gut. Neuroscience 1992, 46, 439–454. [Google Scholar] [CrossRef]
- Fang, Y.; Ren, R.; Shi, H.; Huang, L.; Lenahan, C.; Lu, Q.; Tang, L.; Huang, Y.; Tang, J.; Zhang, J.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide: A Promising Neuroprotective Peptide in Stroke. Aging Dis. 2020, 11, 1496–1512. [Google Scholar] [CrossRef] [PubMed]
- Solés-Tarrés, I.; Cabezas-Llobet, N.; Vaudry, D.; Xifró, X. Protective Effects of Pituitary Adenylate Cyclase-Activating Polypeptide and Vasoactive Intestinal Peptide against Cognitive Decline in Neurodegenerative Diseases. Front. Cell. Neurosci. 2020, 14, 221. [Google Scholar] [CrossRef] [PubMed]
- Marzagalli, R.; Scuderi, S.; Drago, F.; Waschek, J.A.; Castorina, A. Emerging Role of PACAP as a New Potential Therapeutic Target in Major Diabetes Complications. Int. J. Endocrinol. 2015, 2015, 160928. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Reitano, R.; Bucolo, C.; Saccone, S.; Drago, F.; D’Agata, V.J. PACAP Modulates Expression of Hypoxia-Inducible Factors in Streptozotocin-Induced Diabetic Rat Retina. Mol. Neurosci. 2015, 57, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef]
- Rerup, C.C. Drugs producing diabetes through damage of the insulin secreting cells. Pharmacol. Rev. 1970, 22, 485e518. [Google Scholar]
- Mungan, Z.; Arimura, A.; Ertan, A.; Rossowski, W.J.; Coy, D.H. Pituitary adenylate cyclase-activating polypeptide relaxes rat gastrointestinal smooth muscle. Scand. J. Gastroenterol. 1992, 27, 375–380. [Google Scholar] [CrossRef]
- Felley, C.P.; Qian, J.M.; Mantey, S.; Pradhan, T.; Jensen, R.T. Chief cells possess a receptor with high affinity for PACAP and VIP that stimulates pepsinogen release. Am. J. Physiol. 1992, 263 Pt 1, G901–G907. [Google Scholar] [CrossRef]
- Jayawardena, D.; Guzman, G.; Gill, R.K.; Alrefai, W.A.; Onyuksel, H.; Dudeja, P.K. Expression and localization of VPAC1, the major receptor of vasoactive intestinal peptide along the length of the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G16–G25. [Google Scholar] [CrossRef] [Green Version]
- Miampamba, M.; Germano, P.M.; Arli, S.; Wong, H.H.; Scott, D.; Taché, Y.; Pisegna, J.R. Expression of pituitary adenylate cyclase-activating polypeptide and PACAP type 1 receptor in the rat gastric and colonic myenteric neurons. Regul. Pept. 2002, 105, 145–154. [Google Scholar] [CrossRef]
- Zeng, N.; Kang, T.; Lyu, R.M.; Wong, H.; Wen, Y.; Walsh, J.H.; Sachs, G.; Pisegna, J.R. The pituitary adenylate cyclase-activating polypeptide type 1 receptor (PAC1-R) is expressed on gastric ECL cells: Evidence by immunocytochemistry and RT-PCR. Ann. N. Y. Acad. Sci. 1998, 865, 147–156. [Google Scholar] [CrossRef]
- Lin, Z.; Sarosiek, I.; Forster, J.; Damjanov, I.; Hou, Q.; McCallum, R.W. Association of the status of interstitial cells of Cajal and electrogastrogram parameters, gastric emptying and symptoms in patients with gastroparesis. Neurogastroenterol. Motil. 2010, 22, 56–61.e10. [Google Scholar]
- Frokjaer, J.B.; Andersen, S.D.; Ejskjaer, N.; Funch-Jensen, P.; Drewes, A.M.; Gregersen, H. Impaired contractility and remodeling of the upper gastrointestinal tract in diabetes mellitus type-1. World J. Gastroenterol. 2007, 13, 4881–4890. [Google Scholar] [CrossRef] [Green Version]
- Manavalan, S.; Getachew, B.; Manaye, K.F.; Khundmiri, S.J.; Csoka, A.B.; McKinley, R.; Tamas, A.; Reglodi, D.; Tizabi, Y. PACAP Protects Against Ethanol and Nicotine Toxicity in SH-SY5Y Cells: Implications for Drinking-Smoking Co-morbidity. Neurotox. Res. 2017, 32, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Atlasz, T.; Koszegi, Z.; Babai, N.; Tamas, A.; Reglodi, D.; Kovacs, P.; Hernádi, I.; Gábriel, R. Microiontophoretically applied PACAP blocks excitatory effects of kainic acid in vivo. Ann. N. Y. Acad. Sci. 2006, 1070, 143–148. [Google Scholar] [CrossRef]
- Vaudry, D.; Pamantung, T.F.; Basille, M.; Rousselle, C.; Fournier, A.; Vaudry, H.; Beauvillain, J.C.; Gonzalez, B.J. PACAP protects cerebellar granule neurons against oxidative stress-induced apoptosis. Eur. J. Neurosci. 2002, 15, 1451–1460. [Google Scholar] [CrossRef] [Green Version]
- Gui, L.R.; Zhou, Y.; Zhang, B.L.; Li, W.B. Pituitary adenylate cyclase activating polypeptide protects neuro-2a cells from beta amyloid protein cytotoxicity by modulating intracellular calcium. Sheng Li Xue Bao 2003, 55, 42–46. [Google Scholar]
- Amato, R.; Biagioni, M.; Cammalleri, M.; Dal Monte, M.; Casini, G. VEGF as a Survival Factor in Ex Vivo Models of Early Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3066–3076. [Google Scholar] [CrossRef] [Green Version]
- Nedvig, K.; Szabó, G.; Csukás, D.; Sándor, J.; Németh, J.; Kovács, K.; Reglődi, D.; Kemény, A.; Wéber, G.; Ferencz, A. Examination of cytoprotective and anti-inflammatory effect of PACAP-38 on small bowel autotransplantation. Magy. Seb. 2013, 66, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Czajkowska, M.; Całka, J. Neurochemistry of Enteric Neurons Following Prolonged Indomethacin Administration in the Porcine Duodenum. Front. Pharmacol. 2020, 11, 564457. [Google Scholar] [CrossRef]
- Gonkowski, S.; Obremski, K.; Calka, J. The Influence of Low Doses of Zearalenone on Distribution of Selected Active Substances in Nerve Fibers within the Circular Muscle Layer of Porcine Ileum. J. Mol. Neurosci. 2015, 56, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Maisey, A. A Practical Approach to Gastrointestinal Complications of Diabetes. Diabetes Ther. 2016, 7, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Sadanandan, N.; Cozene, B.; Park, Y.J.; Farooq, J.; Kingsbury, C.; Wang, Z.J.; Moscatello, A.; Saft, M.; Cho, J.; Gonzales-Portillo, B.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide: A Potent Therapeutic Agent in Oxidative Stress. Antioxidants 2021, 10, 354. [Google Scholar] [CrossRef]
- Kasica, N.; Podlasz, P.; Sundvik, M.; Tamas, A.; Reglodi, D.; Kaleczyc, J. Protective Effects of Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) Against Oxidative Stress in Zebrafish Hair Cells. Neurotox. Res. 2016, 30, 633–647. [Google Scholar] [CrossRef] [Green Version]
- Illes, A.; Opper, B.; Reglodi, D.; Kerenyi, M.; Czetany, P.; Boronkai, A.; Schafer, E.; Toth, G.; Fabian, E.; Horvath, G. Effects of pituitary adenylate cyclase activating polypeptide on small intestinal INT 407 cells. Neuropeptides 2017, 65, 106–113. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Dunay, I.R.; Schulze, S.; Fischer, A.; Grundmann, U.; Alutis, M.; Kühl, A.A.; Tamas, A.; Toth, G.; Dunay, M.P.; et al. Pituitary adenylate cyclase-activating polypeptide ameliorates experimental acute ileitis and extra-intestinal sequelae. PLoS ONE 2014, 9, e108389. [Google Scholar] [CrossRef] [Green Version]
- Azuma, Y.T.; Hagi, K.; Shintani, N.; Kuwamura, M.; Nakajima, H.; Hashimoto, H.; Baba, A.; Takeuchi, T. PACAP provides colonic protection against dextran sodium sulfate induced colitis. J. Cell. Physiol. 2008, 216, 111–119. [Google Scholar] [CrossRef]
- Läuff, J.M.; Modlin, I.M.; Tang, L.H. Biological relevance of pituitary adenylate cyclase-activating polypeptide (PACAP) in the gastrointestinal tract. Regul. Pept. 1999, 84, 1–12. [Google Scholar] [CrossRef]
- Palus, K.; Bulc, M.; Całka, J. Changes in Somatostatin-Like Immunoreactivity in the Sympathetic Neurons Projecting to the Prepyloric Area of the Porcine Stomach Induced by Selected Pathological Conditions. BioMed Res. Int. 2017, 2017, 9037476. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palus, K.; Bulc, M.; Całka, J.; Zielonka, Ł.; Nowicki, M. Diabetes Affects the Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)-Like Immunoreactive Enteric Neurons in the Porcine Digestive Tract. Int. J. Mol. Sci. 2021, 22, 5727. https://doi.org/10.3390/ijms22115727
Palus K, Bulc M, Całka J, Zielonka Ł, Nowicki M. Diabetes Affects the Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)-Like Immunoreactive Enteric Neurons in the Porcine Digestive Tract. International Journal of Molecular Sciences. 2021; 22(11):5727. https://doi.org/10.3390/ijms22115727
Chicago/Turabian StylePalus, Katarzyna, Michał Bulc, Jarosław Całka, Łukasz Zielonka, and Marcin Nowicki. 2021. "Diabetes Affects the Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)-Like Immunoreactive Enteric Neurons in the Porcine Digestive Tract" International Journal of Molecular Sciences 22, no. 11: 5727. https://doi.org/10.3390/ijms22115727