Autographivirinae Bacteriophage Arno 160 Infects Pectobacterium carotovorum via Depolymerization of the Bacterial O-Polysaccharide


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Bacteriophage Arno 160
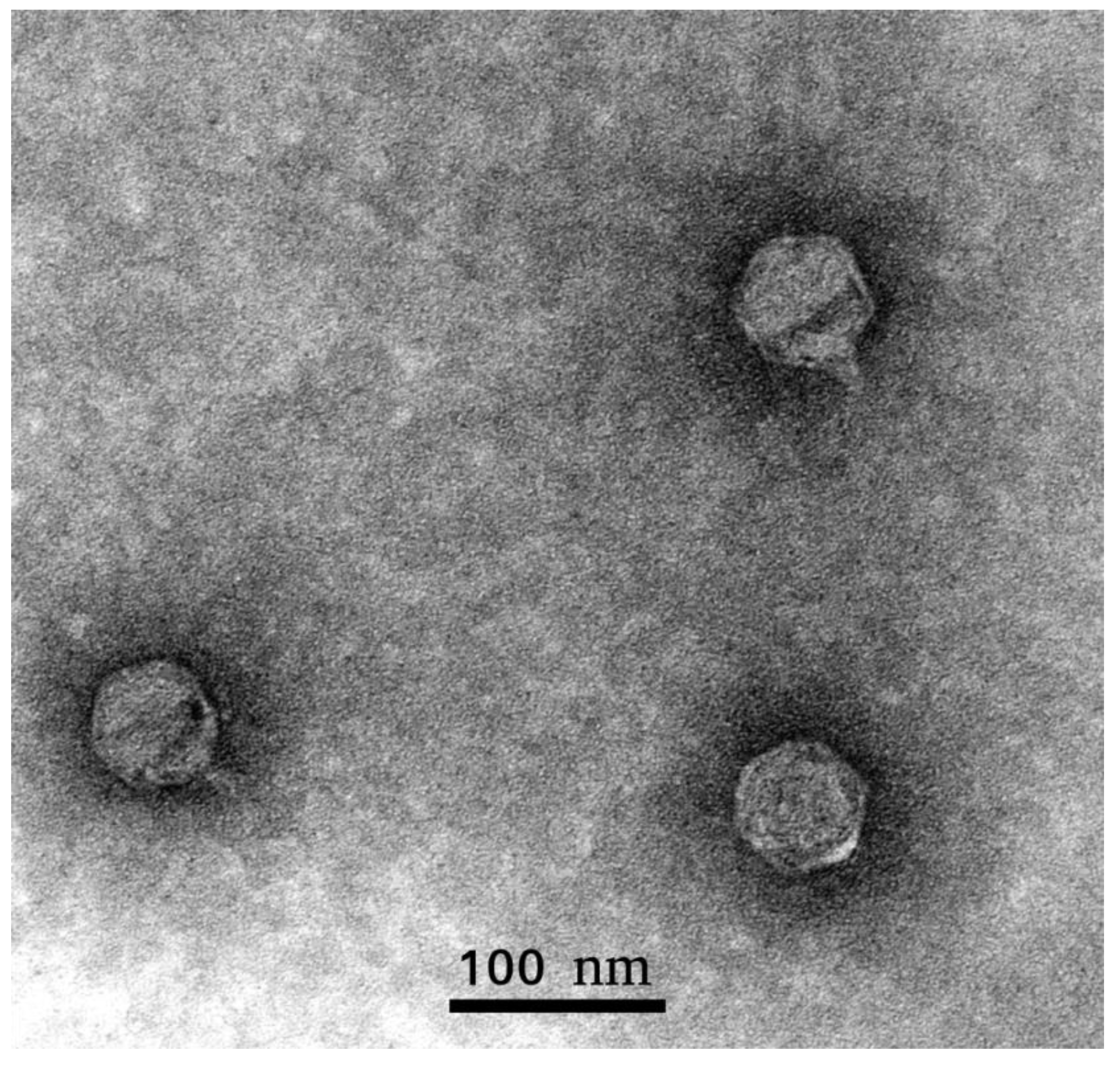
2.1.1. Biology and Morphology
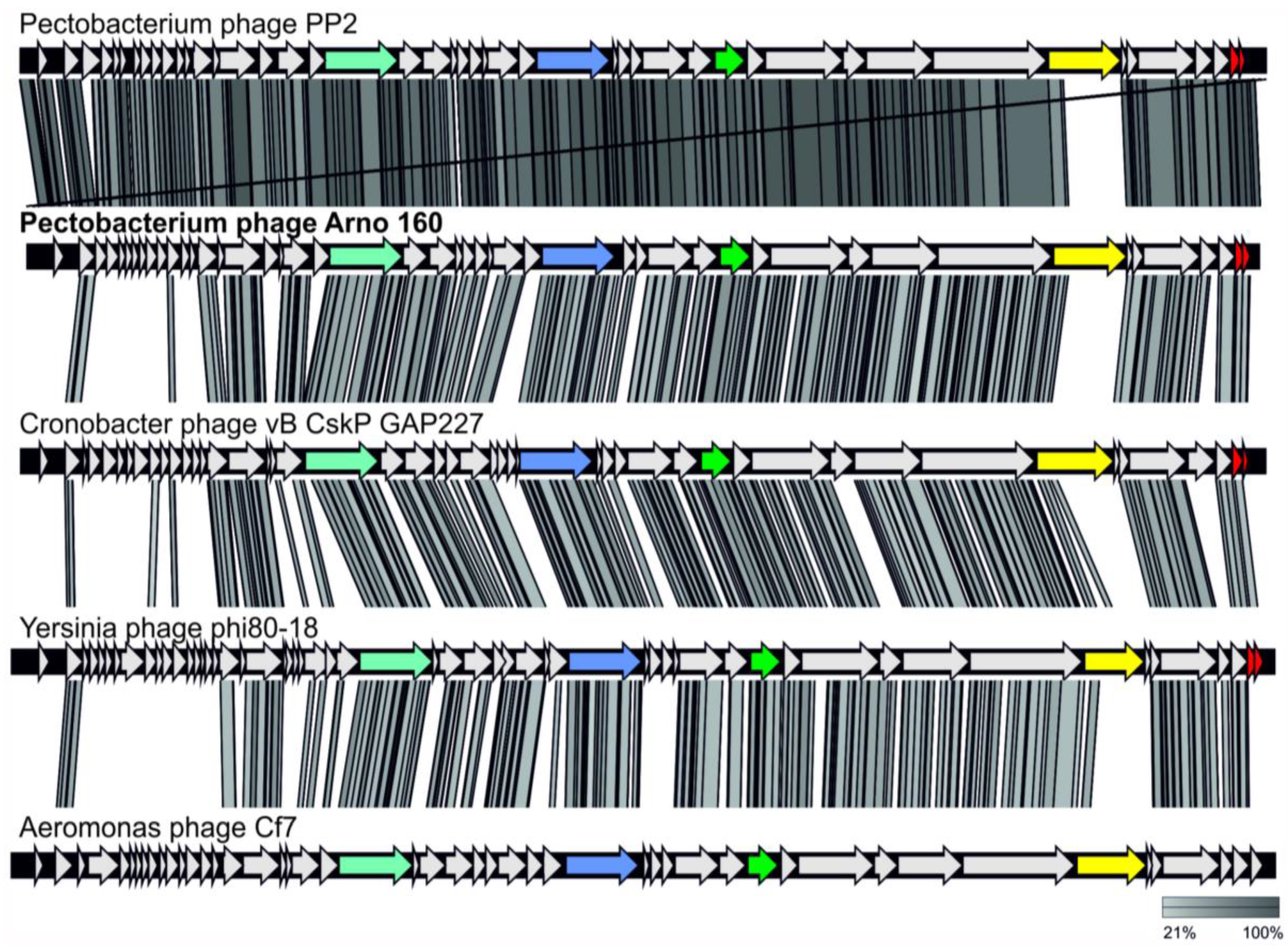
2.1.2. Genome Analysis
2.1.3. Taxonomy
2.1.4. Adsorption Proteins
2.1.5. Lysis Cassette
2.2. Pectobacterium Carotovorum Strain F160. Genomic Analysis
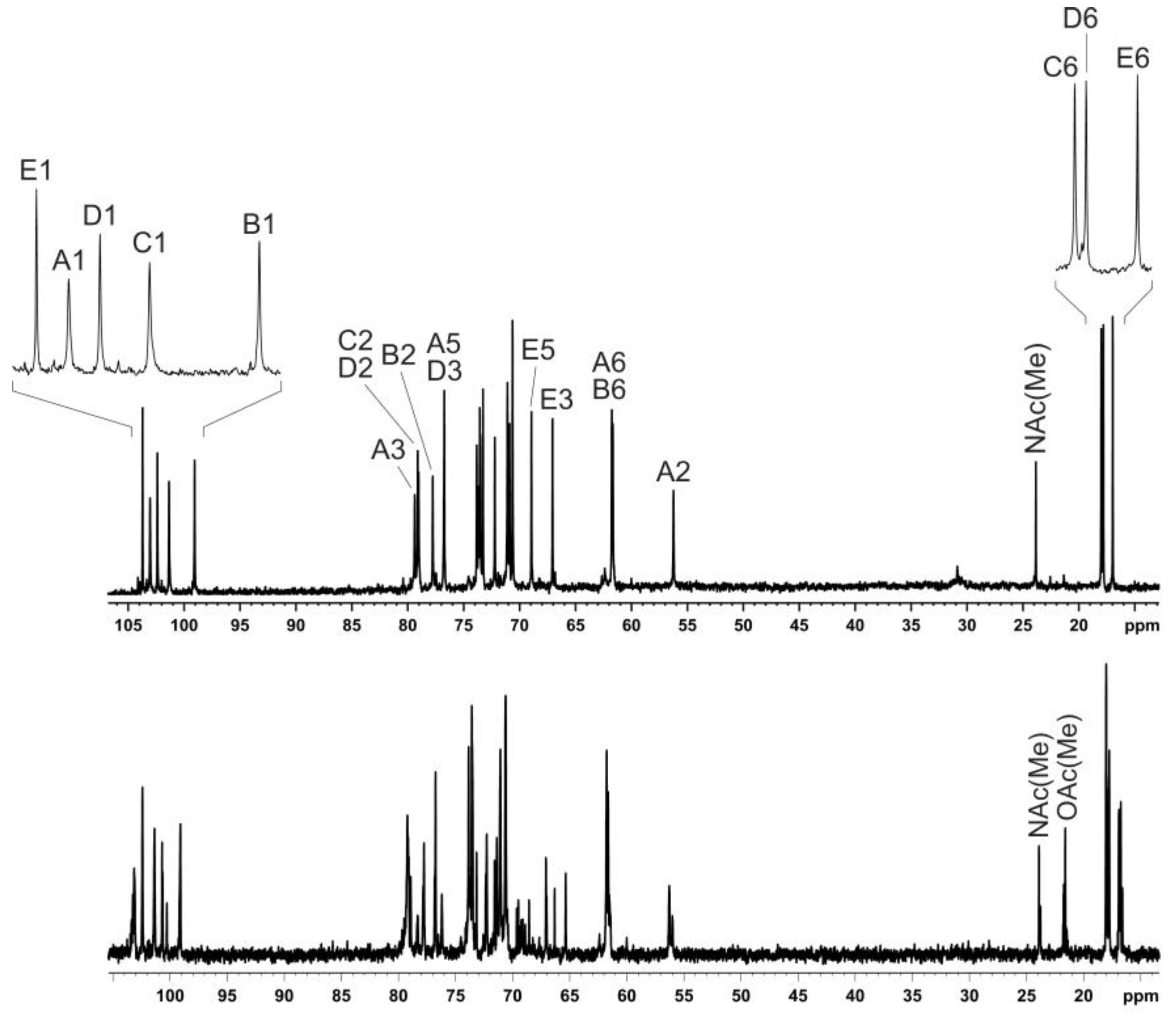
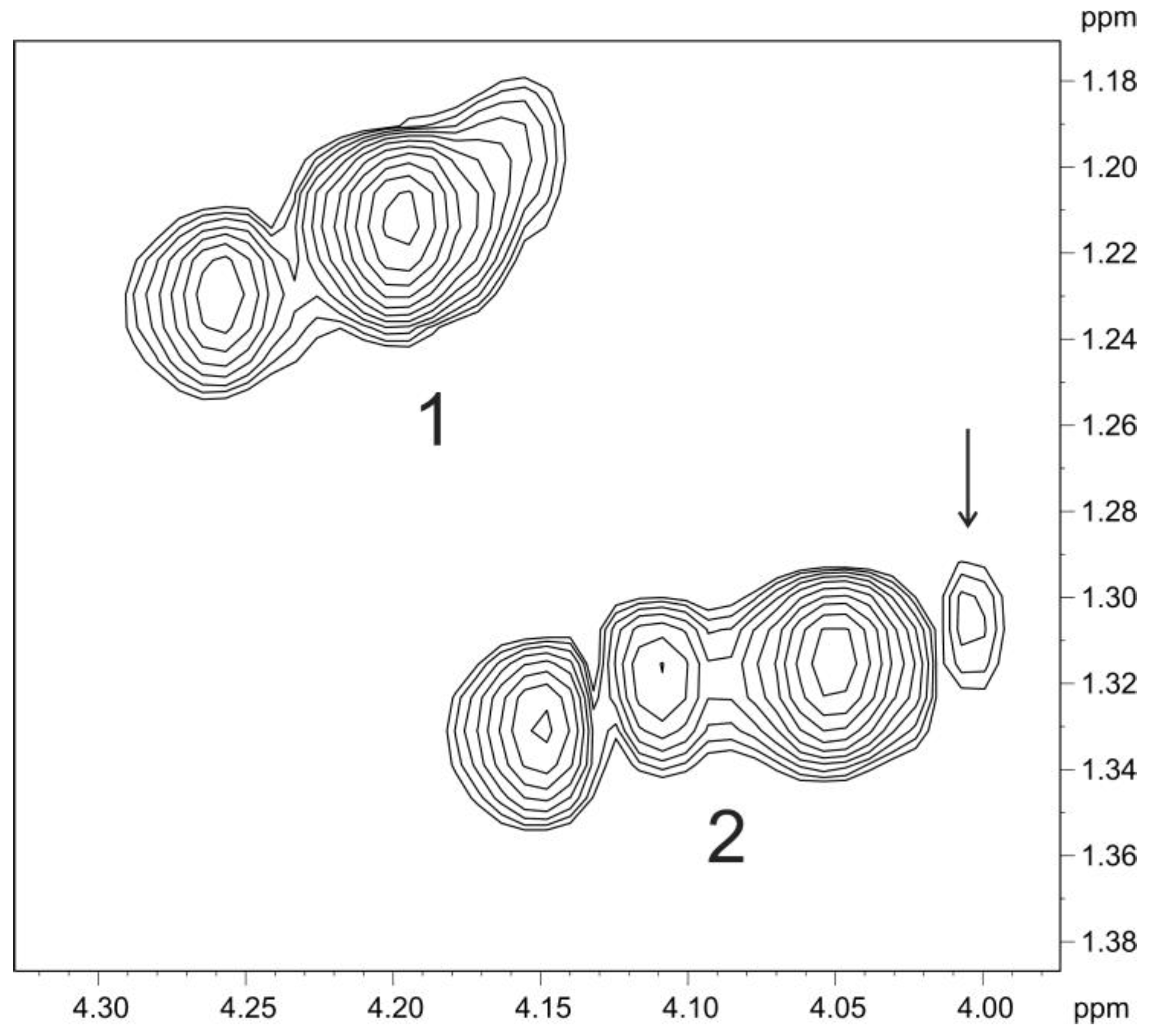
2.3. Structure of the O-polysaccharide of P. carotovorum F160
2.4. Depolymerization of Bacterial Polysaccharide by the Tail Spike Protein gp41
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Phage Isolation and Purification
4.3. Electron Microscopy
4.4. Host Range and General Characterization
4.5. Phage Sequencing and Annotation
4.6. Phylogeny and Taxonomy Studies
4.7. Genome Comparison, Gene and Protein Analysis
4.8. Molecular Cloning and TSP Purification
4.9. Isolation and O-Deacetylation of the O-Polysaccharide
4.10. Depolymerization of the Polysaccharide by the Tail Spike Protein gp41 and Isolation of the Degradation Product
4.11. Sugar Analysis
4.12. NMR Spectroscopy
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SRP | Soft Rot Pectobacteriaceae |
| NMR | Nuclear Magnetic Resonance |
| Rha | Rhamnose |
| Tal | Talose |
| Glc | Glucosamine |
| Pca | Pectobacterium carotovorum |
| OPS | O-polysaccharide |
| OS | Oligosaccharide |
| TSP | Tail spike protein |
| RM | Restriction-modification |
| DPS | O-deacetylated polysaccharide |
References
- Pérombelon, M.C.M. Potato diseases caused by soft rot erwinias: An overview of pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanun, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, Y.; Liang, S.; Tian, Y.; Yin, S.; Xie, S.; Xie, H. Comparative genomics of 84 Pectobacterium genomes reveals the variations related to a pathogenic lifestyle. BMC Genom. 2018, 19, 889. [Google Scholar] [CrossRef] [Green Version]
- Arízala-Quinto, D.; Arif, M. Arif Genome-Wide Analyses Revealed Remarkable Heterogeneity in Pathogenicity Determinants, Antimicrobial Compounds, and CRISPR-Cas Systems of Complex Phytopathogenic Genus Pectobacterium. Pathogens 2019, 8, 247. [Google Scholar] [CrossRef] [Green Version]
- Oulghazi, S.; Cigna, J.; Lau, Y.Y.; Moumni, M.; Chan, K.-G.; Faure, D. Transfer of the waterfall source isolate Pectobacterium carotovorum M022 to Pectobacterium fontis sp. nov., a deep-branching species within the genus Pectobacterium. Int. J. Syst. Evol. Microbiol. 2019, 69, 470–475. [Google Scholar] [CrossRef]
- Dees, M.W.; Lysøe, E.; Rossmann, S.; Perminow, J.; Brurberg, M.B. Pectobacterium polaris sp. nov., isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2017, 67, 5222–5229. [Google Scholar] [CrossRef]
- Pédron, J.; Bertrand, C.; Taghouti, G.; Portier, P.; Barny, M.-A. Pectobacterium aquaticum sp. nov., isolated from waterways. Int. J. Syst. Evol. Microbiol. 2019, 69, 745–751. [Google Scholar] [CrossRef]
- Pasanen, M.; Waleron, M.; Schott, T.; Cleenwerck, I.; Misztak, A.; Waleron, K.; Pritchard, L.; Bakr, R.; Degefu, Y.; Van Der Wolf, J.; et al. Pectobacterium parvum sp. nov., having a Salmonella SPI-1-like Type III secretion system and low virulence. Int. J. Syst. Evol. Microbiol. 2020. [Google Scholar] [CrossRef]
- Portier, P.; Pédron, J.; Taghouti, G.; Saux, M.F.-L.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [CrossRef]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Buttimer, C.; Hendrix, H.; Lucid, A.; Neve, H.; Noben, J.-P.; Franz, C.M.; O’Mahony, J.; Lavigne, R.; Coffey, A. Novel N4-Like Bacteriophages of Pectobacterium atrosepticum. Pharmaceuticals 2018, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- Carstens, A.B.; Djurhuus, A.M.; Kot, W.; Hansen, L.H. A novel six-phage cocktail reduces Pectobacterium atrosepticum soft rot infection in potato tubers under simulated storage conditions. FEMS Microbiol. Lett. 2019, 366, i97–i104. [Google Scholar] [CrossRef]
- Lim, J.-A. Biocontrol of Pectobacterium carotovorum subsp. carotovorum Using Bacteriophage PP1. J. Microbiol. Biotechnol. 2013, 23, 1147–1153. [Google Scholar] [CrossRef] [Green Version]
- Muturi, P.; Yu, J.; Maina, A.N.; Kariuki, S.; Mwaura, F.B.; Wei, H. Bacteriophages Isolated in China for the Control of Pectobacterium carotovorum Causing Potato Soft Rot in Kenya. Virol. Sin. 2019, 34, 287–294. [Google Scholar] [CrossRef]
- Kim, H.; Kim, M.; Bai, J.; Lim, J.-A.; Heu, S.; Ryu, S. Colanic Acid Is a Novel Phage Receptor of Pectobacterium carotovorum subsp. carotovorum Phage POP72. Front. Microbiol. 2019, 10, 143. [Google Scholar] [CrossRef]
- Ackermann, H.W. 5500 Phages examined in the electron microscope. Arch. Virol. 2006, 152, 227–243. [Google Scholar] [CrossRef]
- Lavigne, R.; Seto, N.; Mahadevan, P.; Ackermann, H.-W.; Kropinski, A.M. Unifying classical and molecular taxonomic classification: Analysis of the Podoviridae using BLASTP-based tools. Res. Microbiol. 2008, 159, 406–414. [Google Scholar] [CrossRef]
- Lim, J.-A.; Heu, S.; Park, J.; Roh, E. Genomic characterization of bacteriophage vB_PcaP_PP2 infecting Pectobacterium carotovorum subsp. carotovorum, a new member of a proposed genus in the subfamily Autographivirinae. Arch. Virol. 2017, 162, 2441–2444. [Google Scholar] [CrossRef] [Green Version]
- Guðbrandsson, J.; Ahi, E.P.; Franzdottir, S.R.; Kapralova, K.H.; Kristjánsson, B.K.; Steinhaeuser, S.S.; Maier, V.H.; Johannesson, I.M.; Snorrason, S.S.; Jónsson, Z.O.; et al. The developmental transcriptome of contrasting Arctic charr (Salvelinus alpinus) morphs. F1000Research 2016, 4, 136. [Google Scholar] [CrossRef]
- Abbasifar, R.; Kropinski, A.M.; Sabour, P.M.; Ackermann, H.-W.; Villa, A.A.; Abbasifar, A.; Griffiths, M.W. The Genome of Cronobacter sakazakii Bacteriophage vB_CsaP_GAP227 Suggests a New Genus within the Autographivirinae. Genome Announc. 2013, 1, e00122-12. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.-M.; Tu, I.-F.; Yang, F.-L.; Ko, T.-P.; Liao, J.-H.; Lin, N.-T.; Wu, C.-Y.; Ren, C.-T.; Wang, A.H.-J.; Chang, C.-M.; et al. Structural basis for fragmenting the exopolysaccharide of Acinetobacter baumannii by bacteriophage ΦAB6 tailspike protein. Sci. Rep. 2017, 7, 42711. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.M.I.; Prokhorov, N.; Guerrero-Ferreira, R.; Shneider, M.M.; Browning, C.; Goldie, K.N.; Stahlberg, H.; Leiman, P.G. Structure of the T4 baseplate and its function in triggering sheath contraction. Nature 2016, 533, 346–352. [Google Scholar] [CrossRef]
- Fraser, J.S.; Yu, Z.; Maxwell, K.L.; Davidson, A.R. Ig-Like Domains on Bacteriophages: A Tale of Promiscuity and Deceit. J. Mol. Boil. 2006, 359, 496–507. [Google Scholar] [CrossRef]
- Fokine, A.; Islam, M.Z.; Zhang, Z.; Bowman, V.D.; Rao, V.B.; Rossmann, M.G. Structure of the Three N-Terminal Immunoglobulin Domains of the Highly Immunogenic Outer Capsid Protein from a T4-Like Bacteriophage. J. Virol. 2011, 85, 8141–8148. [Google Scholar] [CrossRef] [Green Version]
- Vernhes, E.; Renouard, M.; Gilquin, B.; Cuniasse, P.; Durand, M.; England, P.; Hoos, S.; Huet, A.; Conway, J.F.; Glukhov, A.; et al. High affinity anchoring of the decoration protein pb10 onto the bacteriophage T5 capsid. Sci. Rep. 2017, 7, 41662. [Google Scholar] [CrossRef]
- Berry, J.; Summer, E.J.; Struck, U.K.; Young, R.; Young, R. The final step in the phage infection cycle: The Rz and Rz1 lysis proteins link the inner and outer membranes. Mol. Microbiol. 2008, 70, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Panda, P.; Lu, A.; Armstrong, K.; Pitman, A.R. Draft Genome Sequence for ICMP 5702, the Type Strain of Pectobacterium carotovorum subsp. carotovorum That Causes Soft Rot Disease on Potato. Genome Announc. 2015, 3, e00875-15. [Google Scholar] [CrossRef] [Green Version]
- Glasner, J.D.; Marquez-Villavicencio, M.; Kim, H.-S.; Jahn, C.E.; Ma, B.; Biehl, B.S.; Rissman, A.I.; Mole, B.; Yi, X.; Yang, C.-H.; et al. Niche-Specificity and the Variable Fraction of thePectobacteriumPan-Genome. Mol. Plant-Microbe Interact. 2008, 21, 1549–1560. [Google Scholar] [CrossRef]
- Hauben, L.; Moore, E.; Vauterin, L.; Steenackers, M.; Mergaert, J.; Verdonck, L.; Swings, J. Phylogenetic Position of Phytopathogens within the Enterobacteriaceae. Syst. Appl. Microbiol. 1998, 21, 384–397. [Google Scholar] [CrossRef]
- Gardan, L.; Shieh, W.-Y.; Chen, Y.-W.; Chaw, S.-M.; Chiu, H.-H. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 381–391. [Google Scholar] [CrossRef]
- Shashkov, A.S.; Lipkind, G.M.; Knirel, Y.A.; Kochetkov, N.K. Stereochemical factors determining the effects of glycosylation on the13C chemical shifts in carbohydrates. Magn. Reson. Chem. 1988, 26, 735–747. [Google Scholar] [CrossRef]
- Leonteln, K.; Lönngren, J. Determination of the absolute configuration of sugars by gas liquid chromatography of their acetylated 2-octyl glycosides. In Methods in Carbohydrate Chemistry; BeMiller, J.N., Whistler, R.L., Shaw, D.H., Eds.; John Wiley & Sons Inc.: New York, NY, USA, 1993; pp. 87–89. [Google Scholar]
- Lipkind, G.M.; Shashkov, A.S.; Knirel, Y.A.; Vinogradov, E.; Kochetkov, N.K. A computer-assisted structural analysis of regular polysaccharides on the basis of 13C-n.m.r. data. Carbohydr. Res. 1988, 175, 59–75. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Paramonov, N.A.; Shashkov, A.S.; Kochetkov, N.K.; Yarullin, R.G.; Farber, S.M.; Efremenko, V. Structure of the polysaccharide chains of Pseudomonas pseudomallei lipopolysaccharides. Carbohydr. Res. 1992, 233, 185–193. [Google Scholar] [CrossRef]
- Czerwicka, M.T.; Marszewska, K.; Bychowska, A.; Dziadziuszko, H.; Brzozowski, K.; Lojkowska, E.; Stepnowski, P.; Kaczyński, Z. Chemical structure of the O-polysaccharide isolated from Pectobacterium atrosepticum SCRI 1039. Carbohydr. Res. 2011, 346, 2978–2981. [Google Scholar] [CrossRef]
- Bock, K.; Pedersen, C. Carbon-13 Nuclear Magnetic Resonance Spectroscopy of Monosaccharides. Adv. Carbohydr. Chem. Biochem. 1983, 41, 27–66. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Shashkov, A.S.; Senchenkova, S.N.; Merino, S.; Tomas, J. Structure of the O-polysaccharide of Aeromonashydrophila O:34; a case of random O-acetylation of 6-deoxy-l-talose. Carbohydr. Res. 2002, 337, 1381–1386. [Google Scholar] [CrossRef]
- Zaczek, M.; Weber-Dąbrowska, B.; Górski, A. Phages in the global fruit and vegetable industry. J. Appl. Microbiol. 2014, 118, 537–556. [Google Scholar] [CrossRef]
- Svircev, A.M.; Roach, D.; Castle, A.J. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, Q.; Loria, R. A re-evaluation of the taxonomy of phytopathogenic genera Dickeya and Pectobacterium using whole-genome sequencing data. Syst. Appl. Microbiol. 2016, 39, 252–259. [Google Scholar] [CrossRef]
- Silva, J.B.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, T.; Ind, A.; Komitopoulou, E.; Salmond, G.P.C. Phage-selected lipopolysaccharide mutants ofPectobacterium atrosepticumexhibit different impacts on virulence. J. Appl. Microbiol. 2010, 109, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, A.P.; Shneider, M.M.; Korzhenkov, A.A.; Bugaeva, E.N.; Miroshnikov, K.K.; Zdorovenko, E.L.; Kulikov, E.E.; Toschakov, S.V.; Ignatov, A.N.; Knirel, Y.A.; et al. Host Specificity of the Dickeya Bacteriophage PP35 Is Directed by a Tail Spike Interaction With Bacterial O-Antigen, Enabling the Infection of Alternative Non-pathogenic Bacterial Host. Front. Microbiol. 2019, 9, 3288. [Google Scholar] [CrossRef] [Green Version]
- Lukianova, A.A.; Shneider, M.M.; Evseev, P.V.; Shpirt, A.M.; Bugaeva, E.N.; Kabanova, A.P.; Obraztsova, E.A.; Miroshnikov, K.K.; Senchenkova, S.N.; Shashkov, A.S.; et al. Morphologically Different Pectobacterium brasiliense Bacteriophages PP99 and PP101: Deacetylation of O-Polysaccharide by the Tail Spike Protein of Phage PP99 Accompanies the Infection. Front. Microbiol. 2020, 10, 3147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorshkov, V.; Islamov, B.; Mikshina, P.; Petrova, O.; Burygin, G.L.; Sigida, E.; Shashkov, A.; Daminova, A.; Ageeva, M.; Idiyatullin, B.; et al. Pectobacterium atrosepticum exopolysaccharides: Identification, molecular structure, formation under stress and in planta conditions. Glycobiology 2017, 27, 1016–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senchenkova, S.N.; Shashkov, A.S.; Knirel, Y.A.; Ahmed, M.; Mavridis, A.; Rudolph, K. Structure of the O-polysaccharide of Erwinia carotovora ssp. atroseptica GSPB 9205 containing a new higher branched monosaccharide. Russ. Chem. Bull. 2005, 54, 1276–1281. [Google Scholar] [CrossRef]
- Ossowska, K.; Czerwicka, M.T.; Sledz, W.; Żołędowska, S.; Motyka-Pomagruk, A.; Szulta, S.; Lojkowska, E.; Kaczyński, Z. The structure of O-polysaccharides isolated from plant pathogenic bacteria Pectobacterium wasabiae IFB5408 and IFB5427. Carbohydr. Res. 2016, 426, 46–49. [Google Scholar] [CrossRef]
- Senchenkova, S.N.; Knirel, Y.A.; Shashkov, A.S.; Ahmed, M.; Mavridis, A.; Rudolph, K. Structure of the O-polysaccharide of Erwinia carotovora ssp. carotovora GSPB 436. Carbohydr. Res. 2003, 338, 2025–2027. [Google Scholar] [CrossRef]
- Ossowska, K.; Czerwicka, M.T.; Sledz, W.; Żołędowska, S.; Motyka-Pomagruk, A.; Golanowska, M.; Condemine, G.; Lojkowska, E.; Kaczyński, Z. The uniform structure of O-polysaccharides isolated from Dickeya solani strains of different origin. Carbohydr. Res. 2017, 445, 40–43. [Google Scholar] [CrossRef]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages: methods and protocols Volume 1: Isolation, Characterization, and Interactions. Methods Mol. Biol. 2009, 502, 113–126. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning-Sambrook & Russel; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001; Volume 18, ISBN 0879695773. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, D.; Chen, G.; Locascio, P.F.; Land, M.; Larimer, F.W.; Hauser, L. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Delcher, A. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.L.; Chang, H.Y.; Daugherty, L.; Fraser, M.; Hunter, S.; López, R.; McAnulla, C.; McMenamin, C.; Nuka, G.; Pesseat, S.; et al. The InterPro protein families database: The classification resource after 15 years. Nucleic Acids Res. 2014, 43, D213–D221. [Google Scholar] [CrossRef] [PubMed]
- Soeding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ågren, J.; Sundström, A.; Håfström, T.; Segerman, B. Gegenees: Fragmented Alignment of Multiple Genomes for Determining Phylogenomic Distances and Genetic Signatures Unique for Specified Target Groups. PLoS ONE 2012, 7, e39107. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Marti-Renom, M.A.; Stuart, A.C.; Sali, A.; Sánchez, R.; Melo, F.; Sali, A. Comparative Protein Structure Modeling of Genes and Genomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291–325. [Google Scholar] [CrossRef] [Green Version]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2017, 27, 14–25. [Google Scholar] [CrossRef]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides. Extraction with phenol-water and further applications of procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- Zych, K.; Toukach, P.V.; Arbatsky, N.P.; Kołodziejska, K.; Senchenkova, S.N.; Shashkov, A.S.; Knirel, Y.A.; Sidorczyk, Z. Structure of the O-specific polysaccharide ofProteus mirabilisD52 and typing of this strain toProteusserogroup O33. JBIC J. Boil. Inorg. Chem. 2001, 268, 4346–4351. [Google Scholar] [CrossRef]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative Determination of Monosaccharides as Their Alditol Acetates by Gas Liquid Chromatography. Anal. Chem. 1965, 37, 1602–1604. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar Residue | C-1 | C-2 | C-3 | C-4 | C-5 | C-6 |
|---|---|---|---|---|---|---|
| H-1 | H-2 | H-3 | H-4 | H-5 | H-6 (6a,6b) | |
| →3)-β-d-GlcpNAc-(1→ | 103.0 | 56.2 | 79.4 | 72.2 | 76.7 | 61.8 |
| A | 4.68 | 3.75 | 3.91 | 3.69 | 3.39 | 3.88, 3.76 |
| →2)-α-d-Glcp- (1→ | 99.1 | 77.8 | 73.8 | 70.6 | 73.5 | 61.7 |
| B | 5.47 | 3.65 | 3.78 | 3.49 | 3.66 | 3.82 |
| →2)-α-l-Rhap-(1→ | 101.3 | 79.1 | 71.1 | 73.55 | 70.6 | 18.0 |
| C | 5.24 | 4.08 | 3.89 | 3.50 | 3.82 | 1.31 |
| →2)-α-l-Rhap-(1→ | 102.4 | 79.0 | 76.7 | 73.7 | 71.1 | 17.8 |
| D | 5.17 | 4.18 | 3.95 | 3.50 | 3.70 | 1.26 |
| α- l-6dTalp-(1 → | 103.7 | 70.9 | 67.1 | 73.3 | 68.9 | 17.0 |
| E | 5.20 | 3.94 | 3.88 | 3.79 | 4.01 | 1.31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shneider, M.M.; Lukianova, A.A.; Evseev, P.V.; Shpirt, A.M.; Kabilov, M.R.; Tokmakova, A.D.; Miroshnikov, K.K.; Obraztsova, E.A.; Baturina, O.A.; Shashkov, A.S.; et al. Autographivirinae Bacteriophage Arno 160 Infects Pectobacterium carotovorum via Depolymerization of the Bacterial O-Polysaccharide. Int. J. Mol. Sci. 2020, 21, 3170. https://doi.org/10.3390/ijms21093170
Shneider MM, Lukianova AA, Evseev PV, Shpirt AM, Kabilov MR, Tokmakova AD, Miroshnikov KK, Obraztsova EA, Baturina OA, Shashkov AS, et al. Autographivirinae Bacteriophage Arno 160 Infects Pectobacterium carotovorum via Depolymerization of the Bacterial O-Polysaccharide. International Journal of Molecular Sciences. 2020; 21(9):3170. https://doi.org/10.3390/ijms21093170
Chicago/Turabian StyleShneider, Mikhail M., Anna A. Lukianova, Peter V. Evseev, Anna M. Shpirt, Marsel R. Kabilov, Anna D. Tokmakova, Kirill K. Miroshnikov, Ekaterina A. Obraztsova, Olga A. Baturina, Alexander S. Shashkov, and et al. 2020. "Autographivirinae Bacteriophage Arno 160 Infects Pectobacterium carotovorum via Depolymerization of the Bacterial O-Polysaccharide" International Journal of Molecular Sciences 21, no. 9: 3170. https://doi.org/10.3390/ijms21093170