Variability in HIV-1 Integrase Gene and 3′-Polypurine Tract Sequences in Cameroon Clinical Isolates, and Implications for Integrase Inhibitors Efficacy

Abstract
:1. Introduction
2. Results
2.1. Patient Demographics
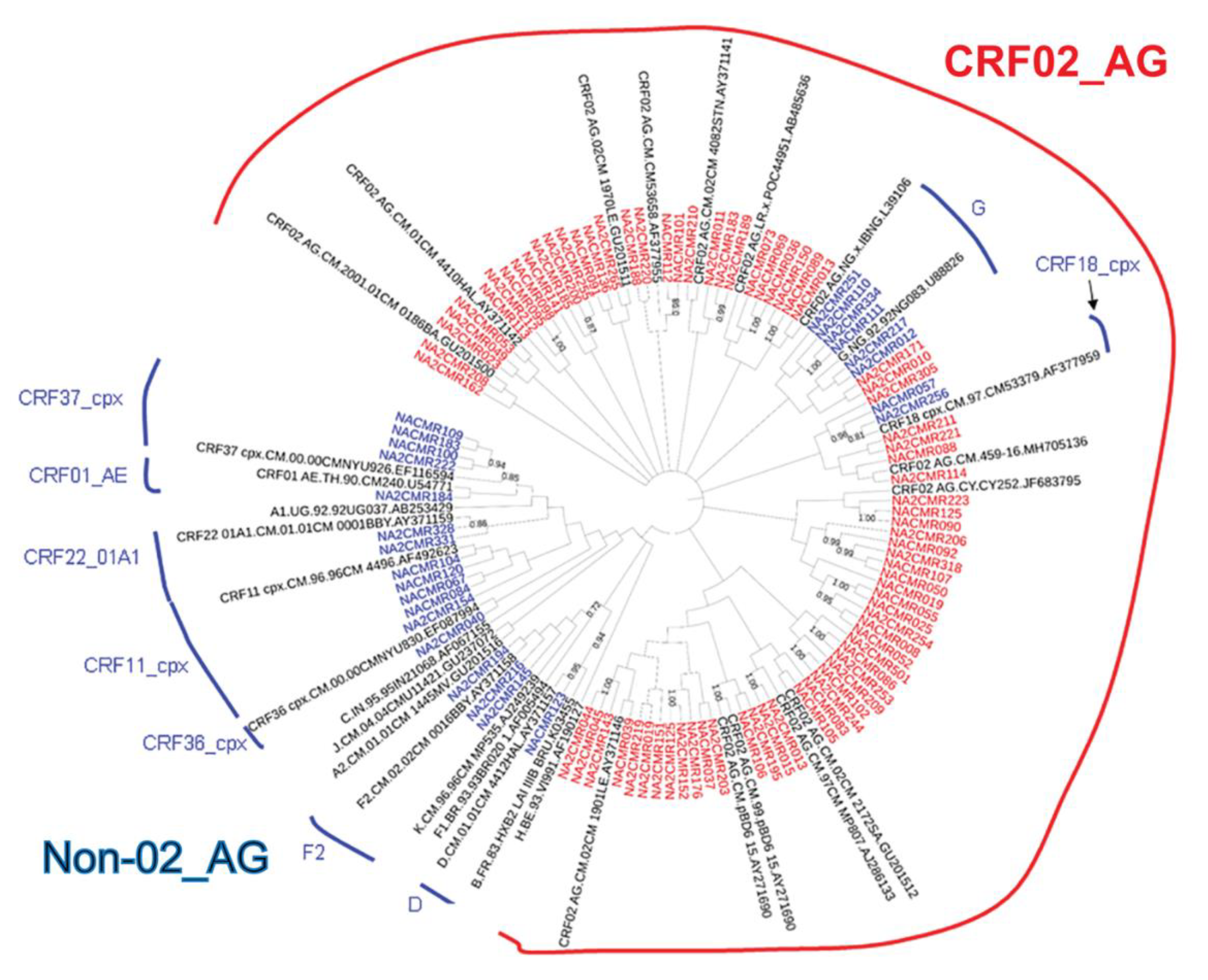
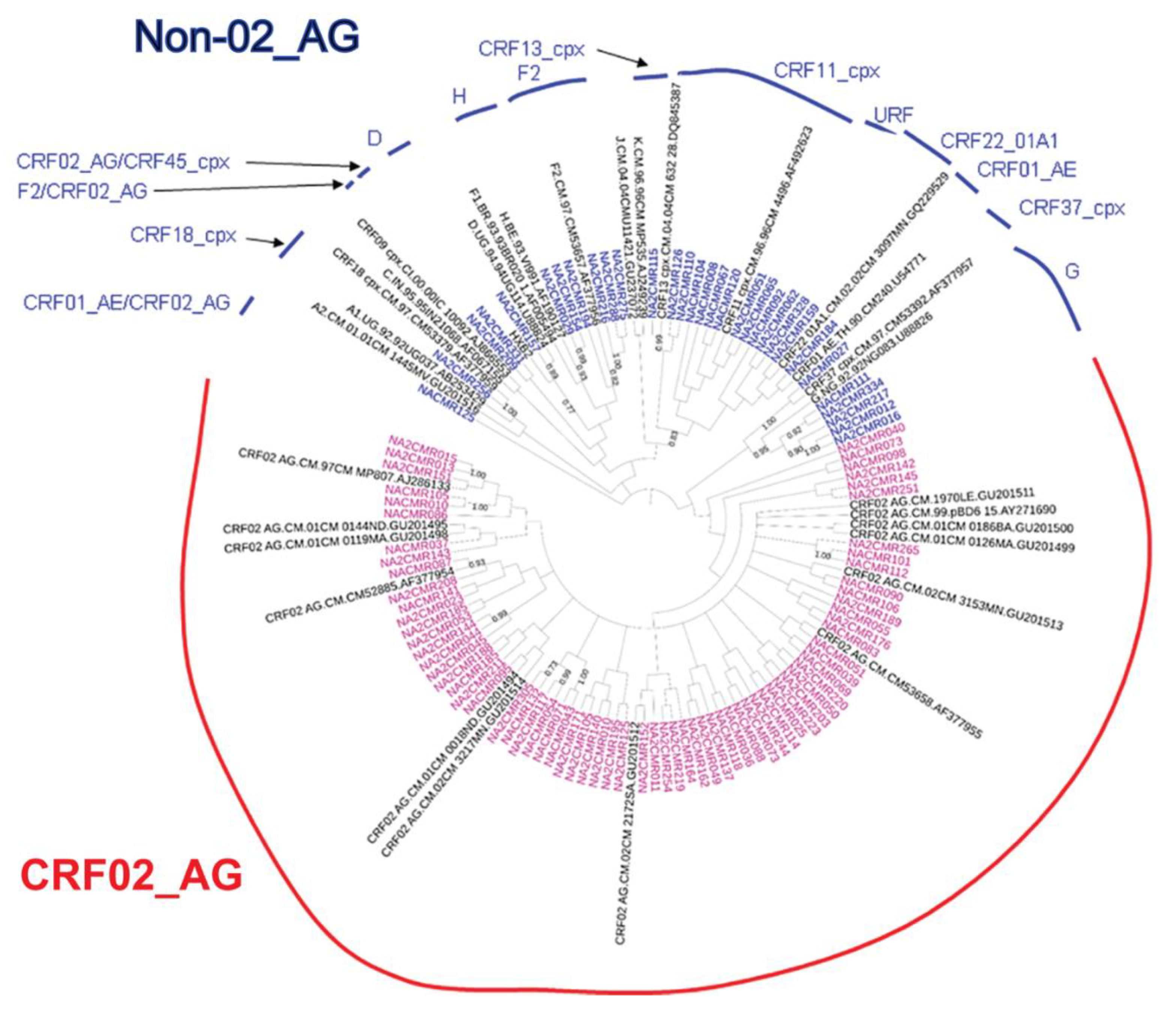
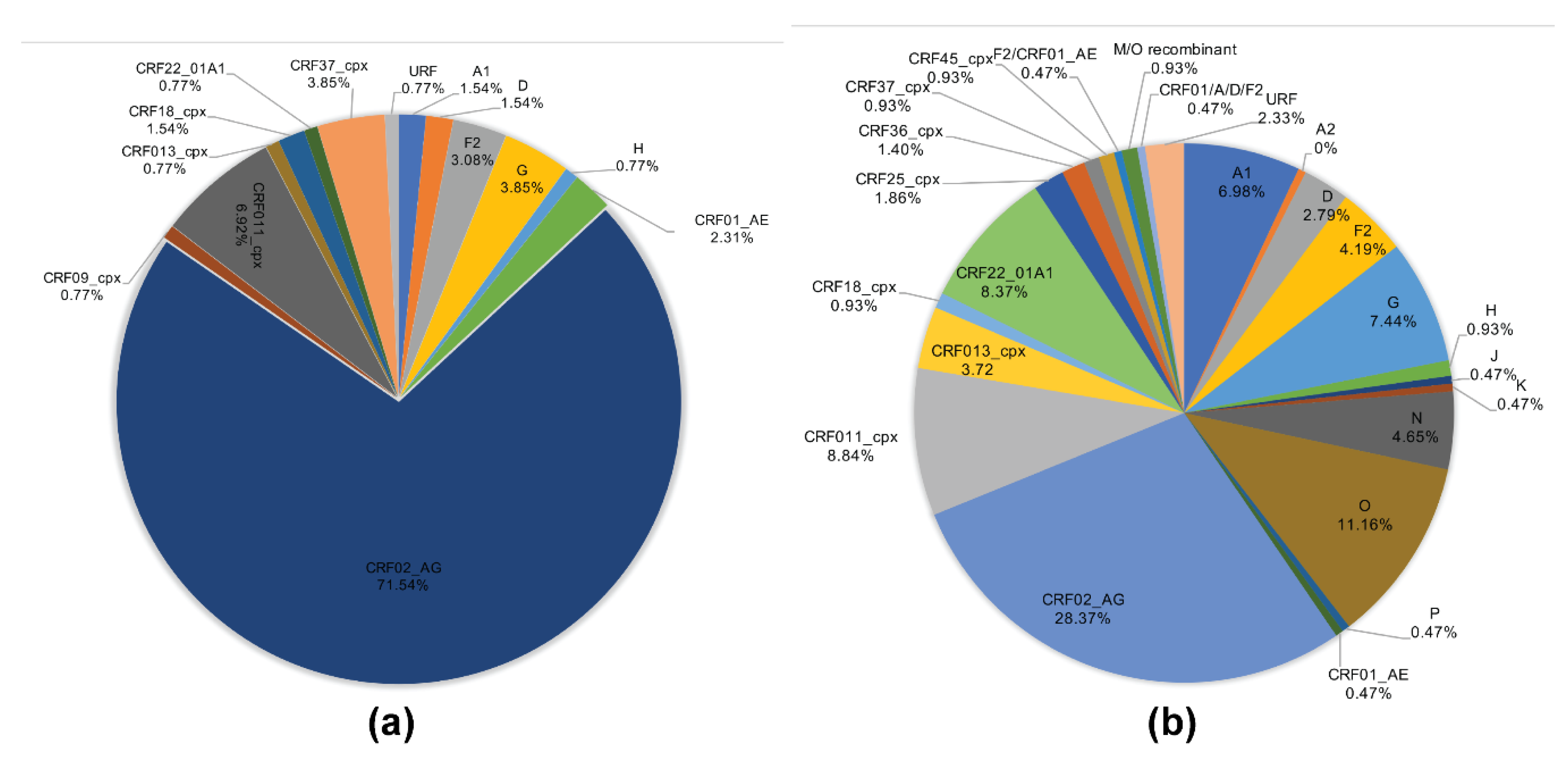
2.2. HIV-1 Subtyping
2.3. INSTI Resistance Mutations in Cohort Samples
2.4. INSTI Resistance Mutations in Database Samples
2.5. INSTIs Resistance-Associated Mutations in Subjects Infected with HIV-1 CRF02_AG and Non-AG Viruses
2.6. Integrase Natural Polymorphisms in Subjects Infected with HIV-1 CRF02_AG and Non-AG Viruses
2.7. Effects of ART and Immune Function on Integrase RAMs and Natural Polymorphisms
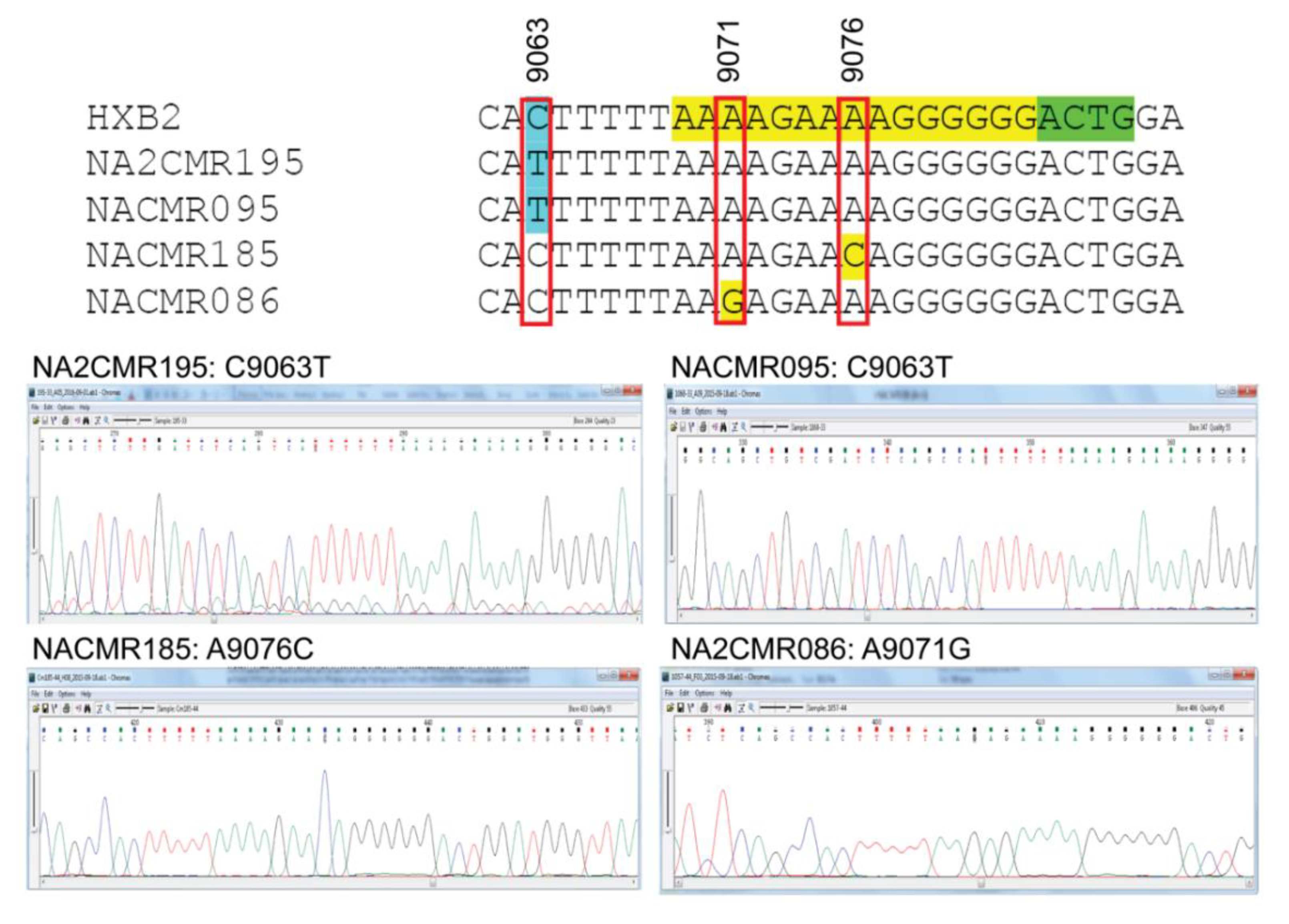
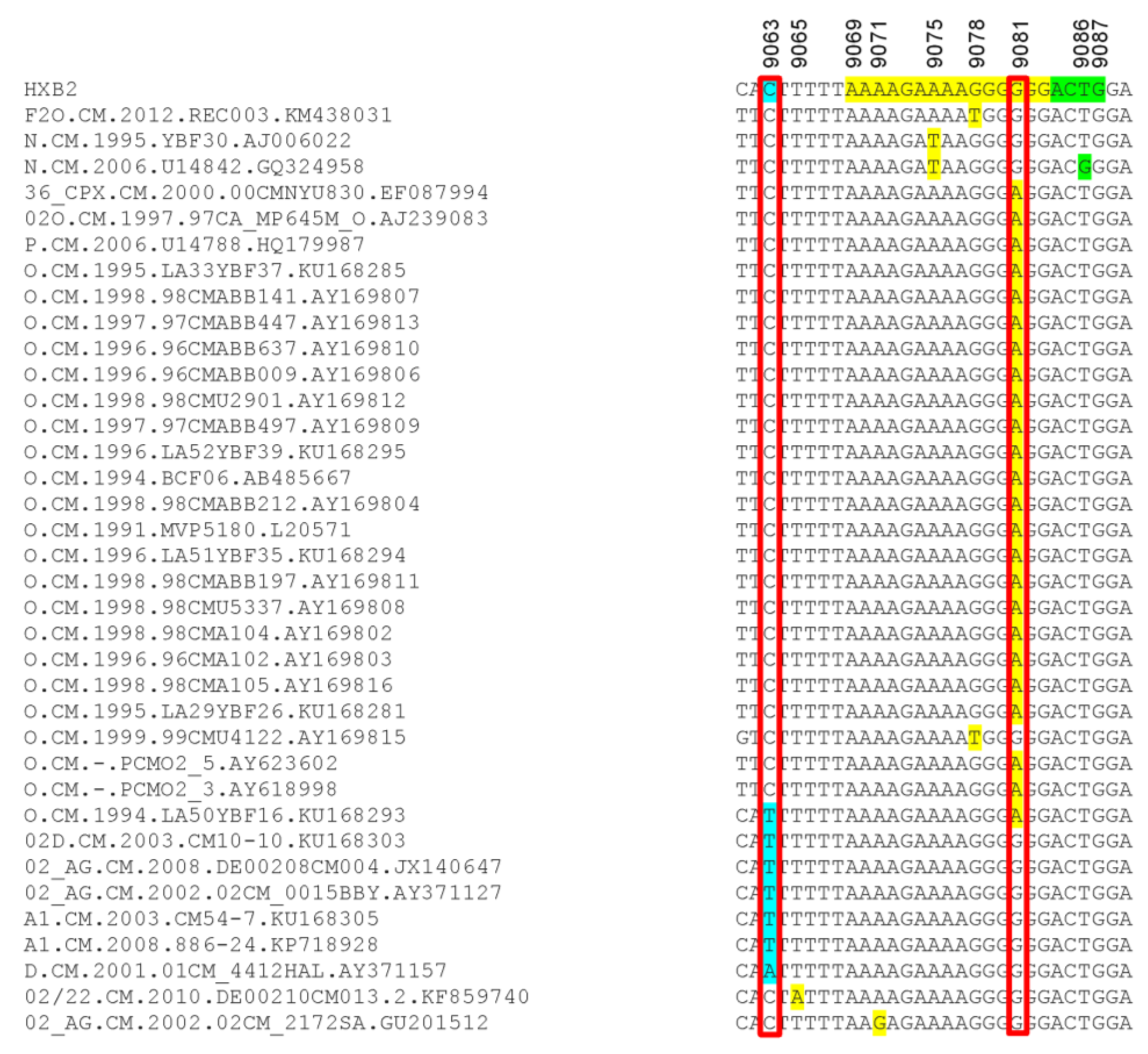
2.8. Analysis of 3′-PPT and 5′ Terminal Nucleotides of 3′ Long Terminal Repeat
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Specimen Collection, HIV Serology, CD4 Cell Counts, and Viral Loads
4.3. RNA Extraction and PCR Amplification
4.4. Gene Sequencing
4.5. Sequence Analysis of the Study Cohort Samples
4.6. Sequence Analysis of the Database Samples
4.7. Statistical Analyses
4.8. Data Availability
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HIV | Human immunodeficiency virus |
| INSTIs | Integrase strand-transfer inhibitors |
| ART | Antiretroviral therapy |
| 3′-PPT | 3′polypurine tract |
| RAMs | Resistance-associated mutations |
| CRF | Circulating recombinant form |
| T | Threonine |
| A | Alanine |
| N | Asparagine |
| K | Lysine |
| E | Glutamic Acid |
| Q | Glutamine |
| M | Methionine |
| I | Isoleucine |
| S | Serine |
| R | Arginine |
| L | Leucine |
| D | Aspartic Acid |
| V | Valine |
| G | Glycine |
| C | Cysteine |
| DNA | Deoxyribonucleic acid |
| kDa | Kilodalton |
| Aa | Amino acids |
| AIDS | Acquired immunodeficiency syndrome |
| FDA | United States Food and Drug Administration |
| RAL | Raltegravir |
| EVG | Elvitegravir |
| DTG | Dolutegravir |
| WHO | World Health Organization |
| SSA | Sub-Saharan Africa |
| PLWH | People living with HIV |
| DRMs | Drug-resistance mutations |
| URF | Unique recombinant form |
| LTR | Long terminal repeats |
| UNMC | University of Nebraska Medical Center |
| ELISA | Enzyme-linked immunosorbent assay |
| CD4 | cluster of differentiation 4 |
| FACS | Fluorescence-Activated Cell Sorting |
| RT-PCR | Reverse transcription polymerase chain reaction |
| RNA | Ribonucleic acid |
| cDNA | complementary DNA |
| PCR | Polymerase chain reaction |
| NCBI | National Center for Biotechnology Information |
| FDR | False discovery rate |
References
- Ceccherini-Silberstein, F.; Malet, I.; D’Arrigo, R.; Antinori, A.; Marcelin, A.G.; Perno, C.F. Characterization and structural analysis of HIV-1 integrase conservation. Aids Rev. 2009, 11, 17–29. [Google Scholar] [PubMed]
- Li, X.; Krishnan, L.; Cherepanov, P.; Engelman, A. Structural biology of retroviral DNA integration. Virology 2011, 411, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesbats, P.; Engelman, A.N.; Cherepanov, P. Retroviral DNA Integration. Chem. Rev. 2016, 116, 12730–12757. [Google Scholar] [CrossRef] [Green Version]
- Asante-Appiah, E.; Skalka, A.M. HIV-1 integrase: Structural organization, conformational changes, and catalysis. Adv. Virus. Res. 1999, 52, 351–369. [Google Scholar]
- Zheng, R.; Jenkins, T.M.; Craigie, R. Zinc folds the N-terminal domain of HIV-1 integrase, promotes multimerization, and enhances catalytic activity. Proc. Natl. Acad. Sci. USA 1996, 93, 13659–13664. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Zheng, R.; Caffrey, M.; Craigie, R.; Clore, G.M.; Gronenborn, A.M. Solution structure of the N-terminal zinc binding domain of HIV-1 integrase. Nat. Struct. Biol. 1997, 4, 567–577. [Google Scholar] [CrossRef]
- DHHS. Guidelines for the Use of Antiretroviral Agents in Adults and Adolescents with HIV. AIDSinfo 2019. Available online: https://aidsinfo.nih.gov/guidelines/html/1/adult-and-adolescent-arv/0 (accessed on 22 June 2019).
- Wong, E.; Trustman, N.; Yalong, A. HIV pharmacotherapy: A review of integrase inhibitors. JAAPA 2016, 29, 36–40. [Google Scholar] [CrossRef]
- Psichogiou, M.; Poulakou, G.; Basoulis, D.; Paraskevis, D.; Markogiannakis, A.; Daikos, G.L. Recent Advances in Antiretroviral Agents: Potent Integrase Inhibitors. Curr Pharm Des. 2017, 23, 2552–2567. [Google Scholar] [CrossRef]
- EACS. Guidelines. European AIDS Clinical Society, 2019. Available online: https://www.eacsociety.org/files/2019_guidelines-10.0_final.pdf (accessed on 21 June 2019).
- WHO. Consolidated Guidelines On THE USE OF ANTIRETROVIRAL DRUGS FOR TREATING AND PREVENTING HIV INFECTION. Recommendations for a Public Health Approach. Second Edition. World Health Organization, 2016. Available online: https://apps.who.int/iris/bitstream/handle/10665/208825/9789241549684_eng.pdf?sequence=1 (accessed on 21 June 2019).
- WHO. Update of Recommendations on First- and Second-Line Antiretroviral Regimens. World Health Organization, 2019. Available online: https://www.who.int/hiv/pub/arv/arv-update-2019-policy/en/ (accessed on 21 February 2020).
- Inzaule, S.C.; Hamers, R.L.; Noguera-Julian, M.; Casadella, M.; Parera, M.; Rinke de Wit, T.F.; Paredes, R. Primary resistance to integrase strand transfer inhibitors in patients infected with diverse HIV-1 subtypes in sub-Saharan Africa. J. Antimicrob. Chemother. 2018, 73, 1167–1172. [Google Scholar] [CrossRef]
- WHO. Guidelines on THE PUBLIC HEALTH RESPONSE TO PRETREATMENT HIV DRUG RESISTANCE World Health Organization 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/255880/9789241550055-eng.pdf;jsessionid=E4F7C6DA3BC67323033C2666046BBC99?sequence=1 (accessed on 21 June 2019).
- MPP. Update on Progress of Sublicensees. Medicines Patent Pool, 2019. Available online: https://medicinespatentpool.org/what-we-do/global-licence-overview/update-on-progress-of-mpp-sublicensees/ (accessed on 4 February 2019).
- Gupta, A.; Juneja, S.; Vitoria, M.; Habiyambere, V.; Nguimfack, B.D.; Doherty, M.; Low-Beer, D. Projected Uptake of New Antiretroviral (ARV) Medicines in Adults in Low- and Middle-Income Countries: A Forecast Analysis 2015-2025. PLoS ONE 2016, 11, e0164619. [Google Scholar] [CrossRef]
- MPP. Five Years on, 3.9 Million People in the Developing World Have Access to HIV Treatment Dolutegravir, Thanks to Access-oriented Voluntary Licensing Agreements. Medicines Patent Pool, 2019. Available online: https://medicinespatentpool.org/mpp-media-post/five-years-on-3-9-million-people-in-the-developing-world-have-access-to-hiv-treatment-dolutegravir-thanks-to-access-oriented-voluntary-licensing-agreements/ (accessed on 4 February 2019).
- WHO. GLOBAL ACTION PLAN ON HIV DRUG RESISTANCE 2017–2021. World Health Organization, 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/255883/9789241512848-eng.pdf?sequence=1 (accessed on 21 June 2019).
- WHO. HIV Drug Resistance Report 2019. World Health Organization, 2019. Available online: https://www.who.int/hiv/pub/drugresistance/hivdr-report-2019/en/ (accessed on 21 February 2020).
- Lepik, K.J.; Harrigan, P.R.; Yip, B.; Wang, L.; Robbins, M.A.; Zhang, W.W.; Toy, J.; Akagi, L.; Lima, V.D.; Guillemi, S.; et al. Emergent drug resistance with integrase strand transfer inhibitor-based regimens. AIDS 2017, 31, 1425–1434. [Google Scholar] [CrossRef]
- Hurt, C.B.; Sebastian, J.; Hicks, C.B.; Eron, J.J. Resistance to HIV integrase strand transfer inhibitors among clinical specimens in the United States, 2009–2012. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2014, 58, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Anstett, K.; Brenner, B.; Mesplede, T.; Wainberg, M.A. HIV drug resistance against strand transfer integrase inhibitors. Retrovirology 2017, 14, 36. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Liu, T.F.; Kiuchi, M.; Zioni, R.; Gifford, R.J.; Holmes, S.P.; Shafer, R.W. Natural variation of HIV-1 group M integrase: Implications for a new class of antiretroviral inhibitors. Retrovirology 2008, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- UNAIDS. Global HIV & AIDS statistics—2019 fact sheet. HIV Epidemic Update. 2019. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 21 February 2019).
- Malet, I.; Subra, F.; Charpentier, C.; Collin, G.; Descamps, D.; Calvez, V.; Marcelin, A.G.; Delelis, O. Mutations Located outside the Integrase Gene Can Confer Resistance to HIV-1 Integrase Strand Transfer Inhibitors. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Wijting, I.E.A.; Lungu, C.; Rijnders, B.J.A.; van der Ende, M.E.; Pham, H.T.; Mesplede, T.; Pas, S.D.; Voermans, J.J.C.; Schuurman, R.; van de Vijver, D.; et al. HIV-1 Resistance Dynamics in Patients With Virologic Failure to Dolutegravir Maintenance Monotherapy. J. Infect. Dis. 2018, 218, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Das, A.T.; Berkhout, B. How Polypurine Tract Changes in the HIV-1 RNA Genome Can Cause Resistance against the Integrase Inhibitor Dolutegravir. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- LANL. HIV Reference Sequence Database; Los Alamos National Library: Los Alamos, NM, USA, 2019. Available online: https://www.hiv.lanl.gov/content/sequence/HIV/mainpage.html (accessed on 21 February 2020).
- Peeters, M.; Liegeois, F.; Torimiro, N.; Bourgeois, A.; Mpoudi, E.; Vergne, L.; Saman, E.; Delaporte, E.; Saragosti, S. Characterization of a highly replicative intergroup M/O human immunodeficiency virus type 1 recombinant isolated from a Cameroonian patient. J. Virol. 1999, 73, 7368–7375. [Google Scholar] [CrossRef] [Green Version]
- Powell, R.L.; Zhao, J.; Konings, F.A.; Tang, S.; Nanfack, A.; Burda, S.; Urbanski, M.M.; Saa, D.R.; Hewlett, I.; Nyambi, P.N. Identification of a novel circulating recombinant form (CRF) 36_cpx in Cameroon that combines two CRFs (01_AE and 02_AG) with ancestral lineages of subtypes A and G. Aids Res. Hum. Retrovir. 2007, 23, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.A.; Bodelle, P.; Coffey, R.; Devare, S.G.; Golden, A.; Hackett, J., Jr.; Harris, B.; Holzmayer, V.; Luk, K.C.; Schochetman, G.; et al. The prevalence of diverse HIV-1 strains was stable in Cameroonian blood donors from 1996 to 2004. J. Acquir. Immune. Defic. Syndr. 2008, 49, 432–439. [Google Scholar] [CrossRef]
- Teto, G.; Fonsah, J.Y.; Tagny, C.T.; Mbanya, D.; Nchindap, E.; Kenmogne, L.; Fokam, J.; Njamnshi, D.M.; Kouanfack, C.; Njamnshi, A.K.; et al. Molecular and Genetic Characterization of HIV-1 Tat Exon-1 Gene from Cameroon Shows Conserved Tat HLA-Binding Epitopes: Functional Implications. Viruses 2016, 8, 196. [Google Scholar] [CrossRef]
- Teto, G.; Tagny, C.T.; Mbanya, D.; Fonsah, J.Y.; Fokam, J.; Nchindap, E.; Kenmogne, L.; Njamnshi, A.K.; Kanmogne, G.D. Gag P2/NC and pol genetic diversity, polymorphism, and drug resistance mutations in HIV-1 CRF02_AG- and non-CRF02_AG-infected patients in Yaounde, Cameroon. Sci. Rep. 2017, 7, 14136. [Google Scholar] [CrossRef] [Green Version]
- Villabona-Arenas, C.J.; Domyeum, J.; Mouacha, F.; Butel, C.; Delaporte, E.; Peeters, M.; Mpoudi-Ngole, E.; Aghokeng, A.F. HIV-1 group O infection in Cameroon from 2006 to 2013: Prevalence, genetic diversity, evolution and public health challenges. Infection, genetics and evolution. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2015, 36, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Margot, N.A.; Hluhanich, R.M.; Jones, G.S.; Andreatta, K.N.; Tsiang, M.; McColl, D.J.; White, K.L.; Miller, M.D. In vitro resistance selections using elvitegravir, raltegravir, and two metabolites of elvitegravir M1 and M4. Antivir. Res. 2012, 93, 288–296. [Google Scholar] [CrossRef]
- Abram, M.E.; Hluhanich, R.M.; Goodman, D.D.; Andreatta, K.N.; Margot, N.A.; Ye, L.; Niedziela-Majka, A.; Barnes, T.L.; Novikov, N.; Chen, X.; et al. Impact of primary elvitegravir resistance-associated mutations in HIV-1 integrase on drug susceptibility and viral replication fitness. Antimicrob. Agents Chemother. 2013, 57, 2654–2663. [Google Scholar] [CrossRef] [Green Version]
- Van der Borght, K.; Verheyen, A.; Feyaerts, M.; Van Wesenbeeck, L.; Verlinden, Y.; Van Craenenbroeck, E.; van Vlijmen, H. Quantitative prediction of integrase inhibitor resistance from genotype through consensus linear regression modeling. Virol. J. 2013, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Winters, M.A.; Lloyd, R.M., Jr.; Shafer, R.W.; Kozal, M.J.; Miller, M.D.; Holodniy, M. Development of elvitegravir resistance and linkage of integrase inhibitor mutations with protease and reverse transcriptase resistance mutations. PLoS ONE 2012, 7, e40514. [Google Scholar] [CrossRef] [Green Version]
- Van Wesenbeeck, L.; Rondelez, E.; Feyaerts, M.; Verheyen, A.; Van der Borght, K.; Smits, V.; Cleybergh, C.; De Wolf, H.; Van Baelen, K.; Stuyver, L.J. Cross-resistance profile determination of two second-generation HIV-1 integrase inhibitors using a panel of recombinant viruses derived from raltegravir-treated clinical isolates. Antimicrob. Agents Chemother. 2011, 55, 321–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fransen, S.; Gupta, S.; Frantzell, A.; Petropoulos, C.J.; Huang, W. Substitutions at amino acid positions 143, 148, and 155 of HIV-1 integrase define distinct genetic barriers to raltegravir resistance in vivo. J. Virol. 2012, 86, 7249–7255. [Google Scholar] [CrossRef] [Green Version]
- Carganico, A.; Dupke, S.; Ehret, R.; Berg, T.; Baumgarten, A.; Obermeier, M.; Walter, H. New dolutegravir resistance pattern identified in a patient failing antiretroviral therapy. J. Int. AIDS Soc. 2014, 17, 19749. [Google Scholar] [CrossRef]
- Hardy, I.; Brenner, B.; Quashie, P.; Thomas, R.; Petropoulos, C.; Huang, W.; Moisi, D.; Wainberg, M.A.; Roger, M. Evolution of a novel pathway leading to dolutegravir resistance in a patient harbouring N155H and multiclass drug resistance. J. Antimicrob. Chemother. 2015, 70, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Nyamache, A.K.; Muigai, A.W.; Nganga, Z.; Khamadi, S.A. HIV Type 1 genetic diversity and naturally occurring polymorphisms in HIV type 1 Kenyan isolates: Implications for integrase inhibitors. Aids Res. Hum. Retrovir. 2012, 28, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Brado, D.; Obasa, A.E.; Ikomey, G.M.; Cloete, R.; Singh, K.; Engelbrecht, S.; Neogi, U.; Jacobs, G.B. Analyses of HIV-1 integrase sequences prior to South African national HIV-treatment program and available of integrase inhibitors in Cape Town, South Africa. Sci. Rep. 2018, 8, 4709. [Google Scholar] [CrossRef]
- Ndashimye, E.; Avino, M.; Kyeyune, F.; Nankya, I.; Gibson, R.M.; Nabulime, E.; Poon, A.F.Y.; Kityo, C.; Mugyenyi, P.; Quinones-Mateu, M.E.; et al. Absence of HIV-1 Drug Resistance Mutations Supports the Use of Dolutegravir in Uganda. Aids Res. Hum. Retrovir. 2018, 34, 404–414. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, E.J.; Choi, J.Y.; Kwon, O.K.; Kim, G.J.; Choi, S.Y.; Kim, S.S. Genetic variation of the HIV-1 integrase region in newly diagnosed anti-retroviral drug-naive patients with HIV/AIDS in Korea. Clinical microbiology and infection. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2011, 17, 1155–1159. [Google Scholar] [CrossRef] [Green Version]
- Nouhin, J.; Donchai, T.; Hoang, K.T.; Ken, S.; Kamkorn, J.; Tran, T.; Ayouba, A.; Peeters, M.; Chaix, M.L.; Lien, T.X.; et al. Natural polymorphisms of HIV-1 CRF01_AE integrase coding region in ARV-naive individuals in Cambodia, Thailand and Vietnam: An ANRS AC12 working group study. Infection, genetics and evolution. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2011, 11, 38–43. [Google Scholar] [CrossRef]
- Kotaki, T.; Khairunisa, S.Q.; Sukartiningrum, S.D.; Witaningrum, A.M.; Rusli, M.; Diansyah, M.N.; Arfijanto, M.V.; Rahayu, R.P.; Nasronudin; Kameoka, M. Detection of drug resistance-associated mutations in human immunodeficiency virus type 1 integrase derived from drug-naive individuals in Surabaya, Indonesia. Aids Res. Hum. Retrovir. 2014, 30, 489–492. [Google Scholar] [CrossRef]
- Kim, Y.; Chin, B.S.; Kim, G.; Shin, H.S. Integrase Strand Transfer Inhibitor Resistance Mutations in Antiretroviral Treatment-naive Patients in Korea: A Prospective, Observational Study. J. Korean. Med. Sci. 2018, 33, e173. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.C.; Chen, I.T.; Wu, K.S.; Tseng, Y.T.; Sy, C.L.; Chen, J.K.; Lee, S.S.; Chen, Y.S. HIV-1 integrase strand-transfer inhibitor resistance in southern Taiwan. Oncotarget 2018, 9, 24927–24935. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.-Y.; Lin, P.-H.; Cheng, C.-L.; Chen, M.-Y.; Sun, H.-Y.; Hsieh, S.-M.; Sheng, W.-H.; Su, Y.-C.; Su, L.-H.; Chang, S.-F.; et al. Prevalence of Integrase Strand Transfer Inhibitors (INSTI) Resistance Mutations in Taiwan. Sci. Rep. 2016, 6, 35779. [Google Scholar] [CrossRef] [Green Version]
- Meixenberger, K.; Yousef, K.P.; Smith, M.R.; Somogyi, S.; Fiedler, S.; Bartmeyer, B.; Hamouda, O.; Bannert, N.; von Kleist, M.; Kucherer, C. Molecular evolution of HIV-1 integrase during the 20 years prior to the first approval of integrase inhibitors. Virol. J. 2017, 14, 223. [Google Scholar] [CrossRef] [Green Version]
- Avi, R.; Huik, K.; Sadam, M.; Karki, T.; Krispin, T.; Ainsalu, K.; Paap, P.; Schmidt, J.; Nikitina, N.; Lutsar, I. Characterization of integrase region polymorphisms in HIV type 1 CRF06_cpx viruses in treatment-naive patients in Estonia. Aids Res. Hum. Retrovir. 2010, 26, 1109–1113. [Google Scholar] [CrossRef]
- Tostevin, A.; White, E.; Dunn, D.; Croxford, S.; Delpech, V.; Williams, I.; Asboe, D.; Pozniak, A.; Churchill, D.; Geretti, A.M.; et al. Recent trends and patterns in HIV-1 transmitted drug resistance in the United Kingdom. Hiv Med. 2017, 18, 204–213. [Google Scholar] [CrossRef]
- Passaes, C.B.; Guimaraes, M.L.; Fernandez, S.L.; Lorete Rdos, S.; Teixeira, S.L.; Fernandez, J.C.; Morgado, M.G. Lack of primary mutations associated with integrase inhibitors among HIV-1 subtypes B, C, and F circulating in Brazil. J. Acquir. Immune. Defic. Syndr. 2009, 51, 7–12. [Google Scholar] [CrossRef]
- Mouscadet, J.F.; Delelis, O.; Marcelin, A.G.; Tchertanov, L. Resistance to HIV-1 integrase inhibitors: A structural perspective. Drug Resist. Updates: Rev. Comment. Antimicrob. Anticancer Chemother. 2010, 13, 139–150. [Google Scholar] [CrossRef]
- Li, Y.; Xuan, S.; Feng, Y.; Yan, A. Targeting HIV-1 integrase with strand transfer inhibitors. Drug Discov. Today 2015, 20, 435–449. [Google Scholar] [CrossRef]
- Mulu, A.; Maier, M.; Liebert, U.G. Lack of integrase inhibitors associated resistance mutations among HIV-1C isolates. J. Transl. Med. 2015, 13, 377. [Google Scholar] [CrossRef] [Green Version]
- Pham, H.T.; Labrie, L.; Wijting, I.E.A.; Hassounah, S.; Lok, K.Y.; Portna, I.; Goring, M.E.; Han, Y.; Lungu, C.; van der Ende, M.E.; et al. The S230R Integrase Substitution Associated With Virus Load Rebound During Dolutegravir Monotherapy Confers Low-Level Resistance to Integrase Strand-Transfer Inhibitors. J. Infect. Dis. 2018, 218, 698–706. [Google Scholar] [CrossRef]
- Fransen, S.; Gupta, S.; Danovich, R.; Hazuda, D.; Miller, M.; Witmer, M.; Petropoulos, C.J.; Huang, W. Loss of raltegravir susceptibility by human immunodeficiency virus type 1 is conferred via multiple nonoverlapping genetic pathways. J. Virol. 2009, 83, 11440–11446. [Google Scholar] [CrossRef] [Green Version]
- Eron, J.J.; Clotet, B.; Durant, J.; Katlama, C.; Kumar, P.; Lazzarin, A.; Poizot-Martin, I.; Richmond, G.; Soriano, V.; Ait-Khaled, M.; et al. Safety and efficacy of dolutegravir in treatment-experienced subjects with raltegravir-resistant HIV type 1 infection: 24-week results of the VIKING Study. J. Infect. Dis. 2013, 207, 740–748. [Google Scholar] [CrossRef]
- Naeger, L.K.; Harrington, P.; Komatsu, T.; Deming, D. Effect of dolutegravir functional monotherapy on HIV-1 virological response in integrase strand transfer inhibitor resistant patients. Antivir. Ther. 2016, 21, 481–488. [Google Scholar] [CrossRef] [Green Version]
- George, J.M.; Kuriakose, S.S.; Dee, N.; Stoll, P.; Lalani, T.; Dewar, R.; Khan, M.A.; Rehman, M.T.; Grossman, Z.; Maldarelli, F.; et al. Rapid Development of High-Level Resistance to Dolutegravir With Emergence of T97A Mutation in 2 Treatment-Experienced Individuals With Baseline Partial Sensitivity to Dolutegravir. Open Forum. Infect. Dis. 2018, 5, ofy221. [Google Scholar] [CrossRef]
- Charpentier, C.; Malet, I.; Andre-Garnier, E.; Storto, A.; Bocket, L.; Amiel, C.; Morand-Joubert, L.; Tumiotto, C.; Nguyen, T.; Maillard, A.; et al. Phenotypic analysis of HIV-1 E157Q integrase polymorphism and impact on virological outcome in patients initiating an integrase inhibitor-based regimen. J. Antimicrob. Chemother. 2018, 73, 1039–1044. [Google Scholar] [CrossRef]
- Hombrouck, A.; Voet, A.; Van Remoortel, B.; Desadeleer, C.; De Maeyer, M.; Debyser, Z.; Witvrouw, M. Mutations in human immunodeficiency virus type 1 integrase confer resistance to the naphthyridine L-870,810 and cross-resistance to the clinical trial drug GS-9137. Antimicrob. Agents Chemother. 2008, 52, 2069–2078. [Google Scholar] [CrossRef] [Green Version]
- Quashie, P.K.; Oliviera, M.; Veres, T.; Osman, N.; Han, Y.S.; Hassounah, S.; Lie, Y.; Huang, W.; Mesplede, T.; Wainberg, M.A. Differential effects of the G118R, H51Y, and E138K resistance substitutions in different subtypes of HIV integrase. J. Virol. 2015, 89, 3163–3175. [Google Scholar] [CrossRef] [Green Version]
- Bar-Magen, T.; Sloan, R.D.; Faltenbacher, V.H.; Donahue, D.A.; Kuhl, B.D.; Oliveira, M.; Xu, H.; Wainberg, M.A. Comparative biochemical analysis of HIV-1 subtype B and C integrase enzymes. Retrovirology 2009, 6, 103. [Google Scholar] [CrossRef] [Green Version]
- Bar-Magen, T.; Donahue, D.A.; McDonough, E.I.; Kuhl, B.D.; Faltenbacher, V.H.; Xu, H.; Michaud, V.; Sloan, R.D.; Wainberg, M.A. HIV-1 subtype B and C integrase enzymes exhibit differential patterns of resistance to integrase inhibitors in biochemical assays. AIDS 2010, 24, 2171–2179. [Google Scholar] [CrossRef]
- Fish, M.Q.; Hewer, R.; Wallis, C.L.; Venter, W.D.; Stevens, W.S.; Papathanasopoulos, M.A. Natural polymorphisms of integrase among HIV type 1-infected South African patients. Aids Res. Hum. Retrovir. 2010, 26, 489–493. [Google Scholar] [CrossRef]
- Maiga, A.I.; Malet, I.; Soulie, C.; Derache, A.; Koita, V.; Amellal, B.; Tchertanov, L.; Delelis, O.; Morand-Joubert, L.; Mouscadet, J.F.; et al. Genetic barriers for integrase inhibitor drug resistance in HIV type-1 B and CRF02_AG subtypes. Antivir. Ther. 2009, 14, 123–129. [Google Scholar]
- Han, Y.S.; Mesplede, T.; Wainberg, M.A. Differences among HIV-1 subtypes in drug resistance against integrase inhibitors. Infection, genetics and evolution. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 46, 286–291. [Google Scholar] [CrossRef]
- Hill, K.J.; Rogers, L.C.; Njenda, D.T.; Burke, D.H.; Sarafianos, S.G.; Sonnerborg, A.; Neogi, U.; Singh, K. Strain-specific effect on biphasic DNA binding by HIV-1 integrase. AIDS 2019, 33, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Cahn, P.; Pozniak, A.L.; Mingrone, H.; Shuldyakov, A.; Brites, C.; Andrade-Villanueva, J.F.; Richmond, G.; Buendia, C.B.; Fourie, J.; Ramgopal, M.; et al. Dolutegravir versus raltegravir in antiretroviral-experienced, integrase-inhibitor-naive adults with HIV: Week 48 results from the randomised, double-blind, non-inferiority SAILING study. Lancet 2013, 382, 700–708. [Google Scholar] [CrossRef]
- Theys, K.; Libin, P.J.K.; Van Laethem, K.; Abecasis, A.B. An Evolutionary Model-Based Approach To Quantify the Genetic Barrier to Drug Resistance in Fast-Evolving Viruses and Its Application to HIV-1 Subtypes and Integrase Inhibitors. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, C.; Soriano, V.; Geretti, A.M.; Zahonero, N.; Garcia, S.; Booth, C.; Gutierrez, F.; Viciana, I.; de Mendoza, C. Resistance associated mutations to dolutegravir (S/GSK1349572) in HIV-infected patients--impact of HIV subtypes and prior raltegravir experience. Antivir. Res. 2011, 90, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Modica, S.; Rossetti, B.; Lombardi, F.; Lagi, F.; Maffeo, M.; D’Autilia, R.; Pecorari, M.; Vicenti, I.; Bruzzone, B.; Magnani, G.; et al. Prevalence and determinants of resistance mutations in HIV-1-infected patients exposed to integrase inhibitors in a large Italian cohort. Hiv Med. 2019, 20, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Saladini, F.; Meini, G.; Bianco, C.; Monno, L.; Punzi, G.; Pecorari, M.; Borghi, V.; Di Pietro, M.; Filice, G.; Gismondo, M.R.; et al. Prevalence of HIV-1 integrase mutations related to resistance to dolutegravir in raltegravir naive and pretreated patients. Clin. Microbiol. Infect 2012, 18, E428–E430. [Google Scholar] [CrossRef] [Green Version]
- Abecasis, A.B.; Deforche, K.; Bacheler, L.T.; McKenna, P.; Carvalho, A.P.; Gomes, P.; Vandamme, A.M.; Camacho, R.J. Investigation of baseline susceptibility to protease inhibitors in HIV-1 subtypes C, F, G and CRF02_AG. Antivir. Ther. 2006, 11, 581–589. [Google Scholar]
- Theys, K.; Vercauteren, J.; Snoeck, J.; Zazzi, M.; Camacho, R.J.; Torti, C.; Schulter, E.; Clotet, B.; Sonnerborg, A.; De Luca, A.; et al. HIV-1 subtype is an independent predictor of reverse transcriptase mutation K65R in HIV-1 patients treated with combination antiretroviral therapy including tenofovir. Antimicrob. Agents Chemother. 2013, 57, 1053–1056. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.; Brenner, B.; Moisi, D.; Detorio, M.; Cesaire, R.; Kurimura, T.; Mori, H.; Essex, M.; Maayan, S.; Wainberg, M.A. Nucleotide and amino acid polymorphisms at drug resistance sites in non-B-subtype variants of human immunodeficiency virus type 1. Antimicrob. Agents Chemother. 2004, 48, 2993–2998. [Google Scholar] [CrossRef] [Green Version]
- Rausch, J.W.; Le Grice, S.F. ‘Binding, bending and bonding’: Polypurine tract-primed initiation of plus-strand DNA synthesis in human immunodeficiency virus. Int. J. Biochem. Cell Biol. 2004, 36, 1752–1766. [Google Scholar] [CrossRef]
- Julias, J.G.; McWilliams, M.J.; Sarafianos, S.G.; Alvord, W.G.; Arnold, E.; Hughes, S.H. Effects of mutations in the G tract of the human immunodeficiency virus type 1 polypurine tract on virus replication and RNase H cleavage. J. Virol. 2004, 78, 13315–13324. [Google Scholar] [CrossRef] [Green Version]
- Malet, I.; Delelis, O.; Valantin, M.A.; Montes, B.; Soulie, C.; Wirden, M.; Tchertanov, L.; Peytavin, G.; Reynes, J.; Mouscadet, J.F.; et al. Mutations associated with failure of raltegravir treatment affect integrase sensitivity to the inhibitor in vitro. Antimicrob. Agents Chemother. 2008, 52, 1351–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubier-Maurin, V.; Saragosti, S.; Perret, J.L.; Mpoudi, E.; Esu-Williams, E.; Mulanga, C.; Liegeois, F.; Ekwalanga, M.; Delaporte, E.; Peeters, M. Genetic characterization of the nef gene from human immunodeficiency virus type 1 group M strains representing genetic subtypes A, B, C, E, F, G, and H. Aids Res. Hum. Retrovir. 1999, 15, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiyar, A. The use of CLUSTAL W and CLUSTAL X for multiple sequence alignment. Methods Mol. Biol. 2000, 132, 221–241. [Google Scholar] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Integrase Strand Transfer Inhibitors Resistance Notes. HIV Drug Resistance Database 2019, Stanford University. HIVdb version 8.9-1. Available online: https://hivdb.stanford.edu/dr-summary/resistance-notes/INSTI/ (accessed on 21 February 2020).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Male | Female | p Value |
|---|---|---|---|
| N (%) | 39 (30%) | 91 (70%) | |
| Age (years; mean ± SD) | 37.28 ± 9.94 | 35.43 ± 8.75 | 0.29 |
| Age range (years) | 18–58 | 20–56 | |
| Education (years; mean ± SD) | 10.50 ± 3.47 | 9.09 ± 4.03 | 0.062 |
| CD4 (mean ± SD) (cells /µL) | 360.3 ± 320.4 | 311.2 ± 184.5 | 0.285 |
| CD4 range (cells /µL) | 4–1657 | 5–894 | |
| Viral load (mean ± SD) (log copies/mL) | 4.64 ± 1.54 | 4.24 ± 1.5 | 0.174 |
| Viral load range (log copies/mL) | 1.60–7.56 | 1.60–7 | |
| ART-Naïve (N) | 33 | 66 | |
| ART-Experienced (N) | 5 | 25 |
| Mutations | HIV-1 Subtype/CRFs | |||||||
|---|---|---|---|---|---|---|---|---|
| A1 | CRF01_AE | CRF02_AG | CRF09_cpx | CRF11_cpx | CRF18_cpx | CRF36_cpx | Total (%) | |
| INSTIs Accessory Resistance Mutations: | ||||||||
| T97A | 1 (1.00) | 1 (1.00) | 2 (2.00) | |||||
| E157Q | 6 (6.00) | 6 (6.00) | ||||||
| Other Mutations: | ||||||||
| M50I | 2 (2.00) | 1 (1.00) | 2 (2.00) | 1 (1.00) | 6 (6.00) | |||
| S119R | 4 (4.00) | 4 (4.00) | ||||||
| L74M | 8 (8.00) | 1 (1.00) | 9 (9.00) | |||||
| L74I | 1 (1.00) | 22 (22.00) | 1 (1.00) | 1 (1.00) | 25 (25.00) | |||
| Mutation | HIV-1 Subtypes/CRFs | ||||||||||
| A1 | A2 | D | F2 | G | K | CRF01_AE | F2/CRF01_AE | CRF02_AG | CRF11_cpx | CRF13_cpx | |
| INSTIs Major Resistance Mutations | |||||||||||
| T66A | |||||||||||
| N155K | |||||||||||
| INSTIs Accessory Resistance Mutations | |||||||||||
| T97A | 1 (0.47) | 1 (0.47) | 1 (0.47) | 1 (0.47) | 5 (2.32) | ||||||
| A128T | |||||||||||
| E157Q | 1 (0.47) | 1 (0.47) | |||||||||
| Other Mutations | |||||||||||
| M50I | 9 (4.18) | 1 (0.47) | 1 (0.47) | 5 (2.32) | 4 (1.86) | 1 (0.47) | 1 (0.47) | 1 (0.47) | 5 (2.32) | 6 (2.79) | 2 (0.93) |
| L74M | 1 (0.47) | 6 (2.79) | 1 (0.47) | 1 (0.47) | |||||||
| L74I | 1 (0.47) | 1 (0.47) | 17 (7.91) | 1 (0.47) | |||||||
| S119R | 1 (0.47) | ||||||||||
| S230N | 1 (0.47) | ||||||||||
| E138D | 1 (0.47) | ||||||||||
| Mutation | HIV-1 Subtypes/CRFs | ||||||||||
| CRF18_cpx | CRF22_01A1 | CRF36_cpx | CRF45_cpx | URF | M/O recombinant | CRF01/A/D/F2 | N | O | P | Total | |
| INSTIs Major Resistance Mutations | |||||||||||
| T66A | 1 (0.47) | 1 (0.47) | |||||||||
| N155K | 1 (0.47) | 1 (0.47) | |||||||||
| INSTIs Accessory Resistance Mutations | |||||||||||
| T97A | 1 (0.47) | 1 (0.47) | 4 (1.86) | 1 (0.47) | 16 (7.44) | ||||||
| A128T | 1 (0.47) | 1 (0.47) | |||||||||
| E157Q | 1 (0.47) | 3 (1.39) | |||||||||
| Other Mutations | |||||||||||
| M50I | 1 (0.47) | 13 (6.05) | 2 (0.93) | 2 (0.93) | 1 (0.47) | 2 (0.93) | 10 (4.65) | 67 (31.16) | |||
| L74M | 9 (4.18) | ||||||||||
| L74I | 3 (1.39) | 1 (0.47) | 1 (0.47) | 24 (11.16) | 49 (22.79) | ||||||
| S119R | 1 (0.47) | ||||||||||
| S230N | 1 (0.47) | 2 (0.93) | |||||||||
| E138D | 1 (0.47) | ||||||||||
| Cohort Samples | ||||
| Gene M | AG (n = 75) (% with M) | Non-AG (n = 25) (% with M) | p Value Fisher | p Value FDR |
| T97A | 1.33 | 4.00 | 0.43939 | 0.53221 |
| E157Q | 8.00 | 0.00 | 0.33214 | 0.53221 |
| M50I | 2.67 | 16.00 | 0.03294 | 0.19764 |
| L74M | 10.67 | 4.00 | 0.44351 | 0.53221 |
| L74I | 29.33 | 12.00 | 0.11097 | 0.33291 |
| S119R | 5.33 | 0.00 | 0.56949 | 0.56949 |
| Database Samples | ||||
| Gene M | AG (n = 61) (% with M) | Non-AG (n =1 19) (% with M) | p Value Fisher | p Value FDR |
| T66A | 0.00 | 1.40 | 0.54941 | 0.73722 |
| N155K | 0.00 | 1.40 | 0.54941 | 0.73722 |
| T97A | 17.62 | 15.36 | 0.67020 | 0.73722 |
| A128T | 0.00 | 1.40 | 0.54941 | 0.73722 |
| E157Q | 3.52 | 2.79 | 1.00000 | 1.00000 |
| M50I | 17.62 | 86.56 | 0.00000 | 0.00000 |
| L74M | 21.15 | 4.19 | 0.00091 | 0.00501 |
| L74I | 59.92 | 44.67 | 0.05837 | 0.21402 |
| S119R | 0.00 | 1.40 | 0.54941 | 0.73722 |
| E138D | 0.00 | 1.40 | 0.54941 | 0.73722 |
| S230N | 3.52 | 1.40 | 0.60510 | 0.73722 |
| Cohort Samples | ||||
| Gene Polymorphism | AG (n = 75) (% with P) | Non-AG (n = 25) (% with P) | p Value Fisher | p Value FDR |
| E11D | 29.33 | 40.00 | 0.33308 | 0.40878 |
| K14R | 77.33 | 56.00 | 0.06957 | 0.11049 |
| R20K | 9.33 | 12.00 | 0.70761 | 0.73483 |
| A21T | 5.33 | 24.00 | 0.01449 | 0.03913 |
| S24N | 13.33 | 24.00 | 0.21976 | 0.28255 |
| V31I | 68.00 | 60.00 | 0.47471 | 0.53405 |
| L63I | 12.00 | 4.00 | 0.44389 | 0.52109 |
| I72V | 16.00 | 44.00 | 0.00638 | 0.02462 |
| L101I | 89.33 | 72.00 | 0.05136 | 0.08666 |
| T112I | 12.00 | 32.00 | 0.03120 | 0.06479 |
| T112V | 84.00 | 64.00 | 0.04702 | 0.08666 |
| T124A | 90.67 | 68.00 | 0.01034 | 0.03103 |
| T125A | 90.67 | 80.00 | 0.16850 | 0.22748 |
| G134D | 10.67 | 28.00 | 0.05136 | 0.08666 |
| G134N | 90.67 | 28.00 | 0.00000 | 0.00000 |
| I135V | 81.33 | 28.00 | 0.00000 | 0.00001 |
| K136Q | 0.00 | 44.00 | 0.00000 | 0.00000 |
| K136T | 92.00 | 36.00 | 0.00000 | 0.00000 |
| D167E | 8.00 | 28.00 | 0.01662 | 0.04079 |
| V201I | 96.00 | 100.00 | 0.57096 | 0.61664 |
| T206S | 97.33 | 52.00 | 0.00000 | 0.00000 |
| I208L | 8.00 | 20.00 | 0.13639 | 0.19381 |
| T218I | 28.00 | 32.00 | 0.80002 | 0.80002 |
| L234I | 97.33 | 88.00 | 0.09808 | 0.14712 |
| D256E | 20.00 | 48.00 | 0.00946 | 0.03103 |
| R269K | 30.67 | 0.00 | 0.00068 | 0.00305 |
| S283G | 64.00 | 88.00 | 0.02489 | 0.05601 |
| Database Samples | ||||
| Gene polymorphism | AG (n = 61) (% with P) | Non-AG (n = 119) (% with P) | p Value Fisher | p Value FDR |
| E11D | 26.23 | 46.10 | 0.01023 | 0.01726 |
| K14R | 75.41 | 57.79 | 0.02262 | 0.03393 |
| R20K | 1.64 | 5.84 | 0.26882 | 0.31558 |
| A21T | 8.20 | 25.97 | 0.00530 | 0.01022 |
| S24N | 9.84 | 5.19 | 0.22585 | 0.27718 |
| V31I | 80.33 | 50.65 | 0.00010 | 0.00024 |
| L63I | 9.84 | 4.55 | 0.18737 | 0.24090 |
| I72V | 31.15 | 39.61 | 0.32761 | 0.36856 |
| L101I | 91.80 | 55.84 | 0.00000 | 0.00000 |
| T112I | 9.84 | 15.58 | 0.36293 | 0.39196 |
| T112V | 90.16 | 76.62 | 0.02780 | 0.03950 |
| T124A | 95.08 | 78.57 | 0.00262 | 0.00545 |
| T125A | 95.08 | 66.23 | 0.00001 | 0.00002 |
| G134D | 11.48 | 29.22 | 0.00863 | 0.01554 |
| G134N | 80.33 | 39.61 | 0.00000 | 0.00000 |
| I135V | 75.41 | 9.09 | 0.00000 | 0.00000 |
| K136Q | 9.84 | 51.30 | 0.00000 | 0.00000 |
| K136T | 85.25 | 18.83 | 0.00000 | 0.00000 |
| D167E | 14.75 | 42.21 | 0.00020 | 0.00045 |
| V201I | 98.36 | 89.61 | 0.06340 | 0.08559 |
| T206S | 91.80 | 46.10 | 0.00000 | 0.00000 |
| I208L | 3.28 | 14.29 | 0.02228 | 0.03393 |
| T218I | 44.26 | 12.34 | 0.00000 | 0.00002 |
| L234I | 98.36 | 96.10 | 0.66536 | 0.69095 |
| D256E | 3.28 | 29.22 | 0.00001 | 0.00003 |
| R269K | 50.82 | 5.84 | 0.00000 | 0.00000 |
| S283G | 70.49 | 68.83 | 0.86579 | 0.86579 |
| Primer # | Position ^ | Sequence (5′ to 3′) |
|---|---|---|
| Integrase gene 1st PCR | ||
| IN12 | 4007 → 4026 | GCAGGATTCGGGATTAGAAG |
| IN13 | 5270 → 5251 | CTTTCTCCTGTATGCAGACC |
| Integrase gene 2nd PCR | ||
| IN1 | 4137 → 4157 | AAGGTCTATCTGGCATGGGTA |
| BH4 | 5222 → 5200 | TCCCCTAGTGGGATGTGTACTTC |
| Nef gene 1st PCR | ||
| Nef5-1e F1 | 8513 → 8533 | GTGCCTCTTCAGCTACCACCG |
| Nef3-3e R1 | 9488 → 9508 | AGCATCTGAGGGTTAGCCACT |
| Nef gene 2nd PCR | ||
| Nef5-1e F2 | 8698 → 8717 | TGGACAGAYAGGGTTATAGAA |
| Nef3-7e R2 | 9448 → 9468 | CACCTCCCCTGGAAAGTCCCC |
| Integrase gene sequencing primers | ||
| IN1 | 4137 → 4157 | AAGGTCTATCTGGCATGGGTA |
| BH4 | 5222 → 5200 | TCCCCTAGTGGGATGTGTACTTC |
| IN4764AS | 4764 → 4743 | CCATTTGTACTGCTGTCTTAA |
| IN4542S | 4542 → 4558 | GCAGGAAGATGGCCAGT |
| Nef gene sequencing primers | ||
| Nef5-1e F2 | 8698 → 8717 | TGGACAGAYAGGGTTATAGAA |
| Nef3-7e R2 | 9448 → 9468 | CACCTCCCCTGGAAAGTCCCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acharya, A.; Tagny, C.T.; Mbanya, D.; Fonsah, J.Y.; Nchindap, E.; Kenmogne, L.; Jihyun, M.; Njamnshi, A.K.; Kanmogne, G.D. Variability in HIV-1 Integrase Gene and 3′-Polypurine Tract Sequences in Cameroon Clinical Isolates, and Implications for Integrase Inhibitors Efficacy. Int. J. Mol. Sci. 2020, 21, 1553. https://doi.org/10.3390/ijms21051553
Acharya A, Tagny CT, Mbanya D, Fonsah JY, Nchindap E, Kenmogne L, Jihyun M, Njamnshi AK, Kanmogne GD. Variability in HIV-1 Integrase Gene and 3′-Polypurine Tract Sequences in Cameroon Clinical Isolates, and Implications for Integrase Inhibitors Efficacy. International Journal of Molecular Sciences. 2020; 21(5):1553. https://doi.org/10.3390/ijms21051553
Chicago/Turabian StyleAcharya, Arpan, Claude T. Tagny, Dora Mbanya, Julius Y. Fonsah, Emilienne Nchindap, Léopoldine Kenmogne, Ma Jihyun, Alfred K. Njamnshi, and Georgette D. Kanmogne. 2020. "Variability in HIV-1 Integrase Gene and 3′-Polypurine Tract Sequences in Cameroon Clinical Isolates, and Implications for Integrase Inhibitors Efficacy" International Journal of Molecular Sciences 21, no. 5: 1553. https://doi.org/10.3390/ijms21051553