Celastrol Alleviates Gamma Irradiation-Induced Damage by Modulating Diverse Inflammatory Mediators

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
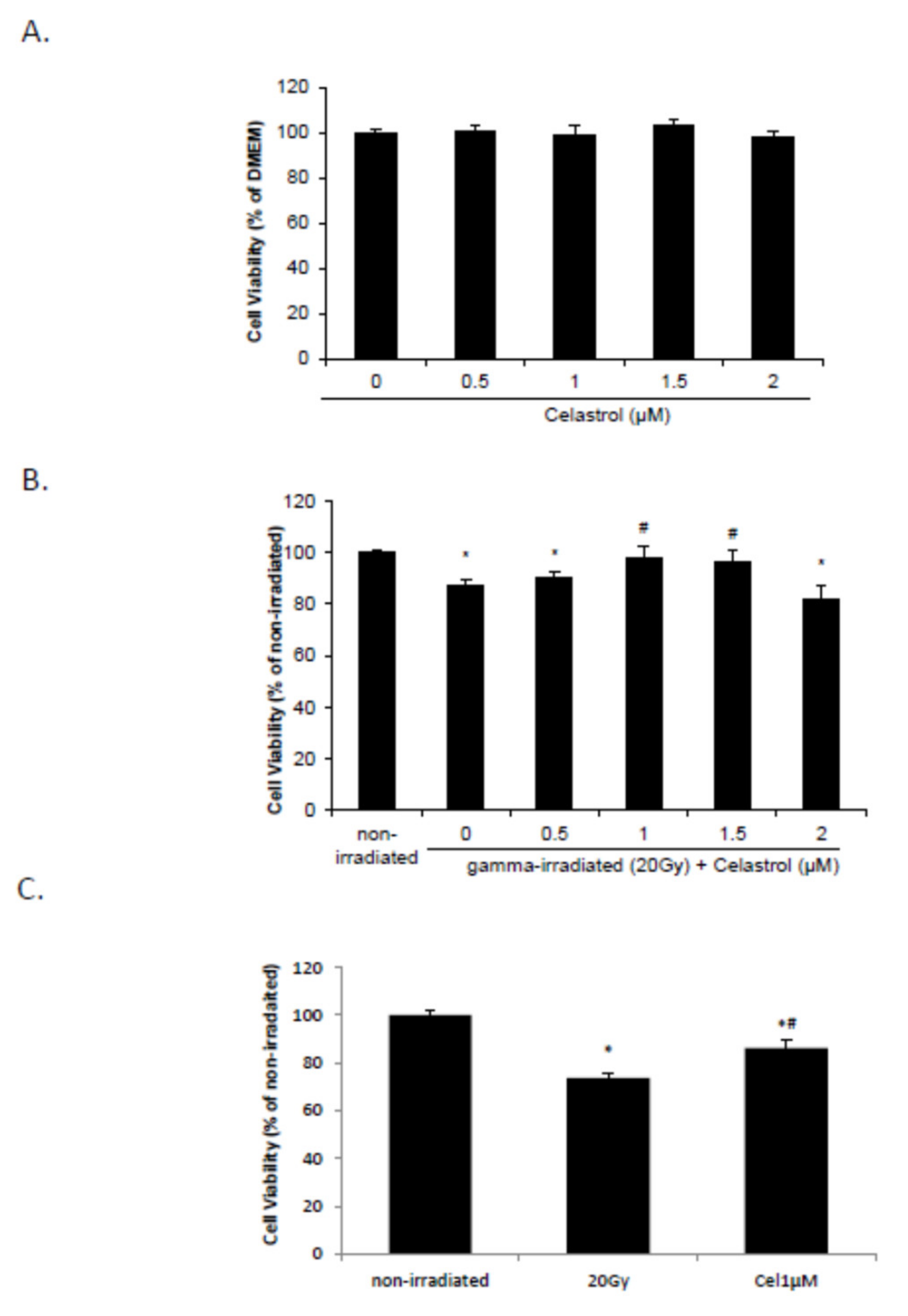
2.1. Celastrol Dose-Dependently Reversed Gamma Irradiation-Induced Decrease in Cell Viability
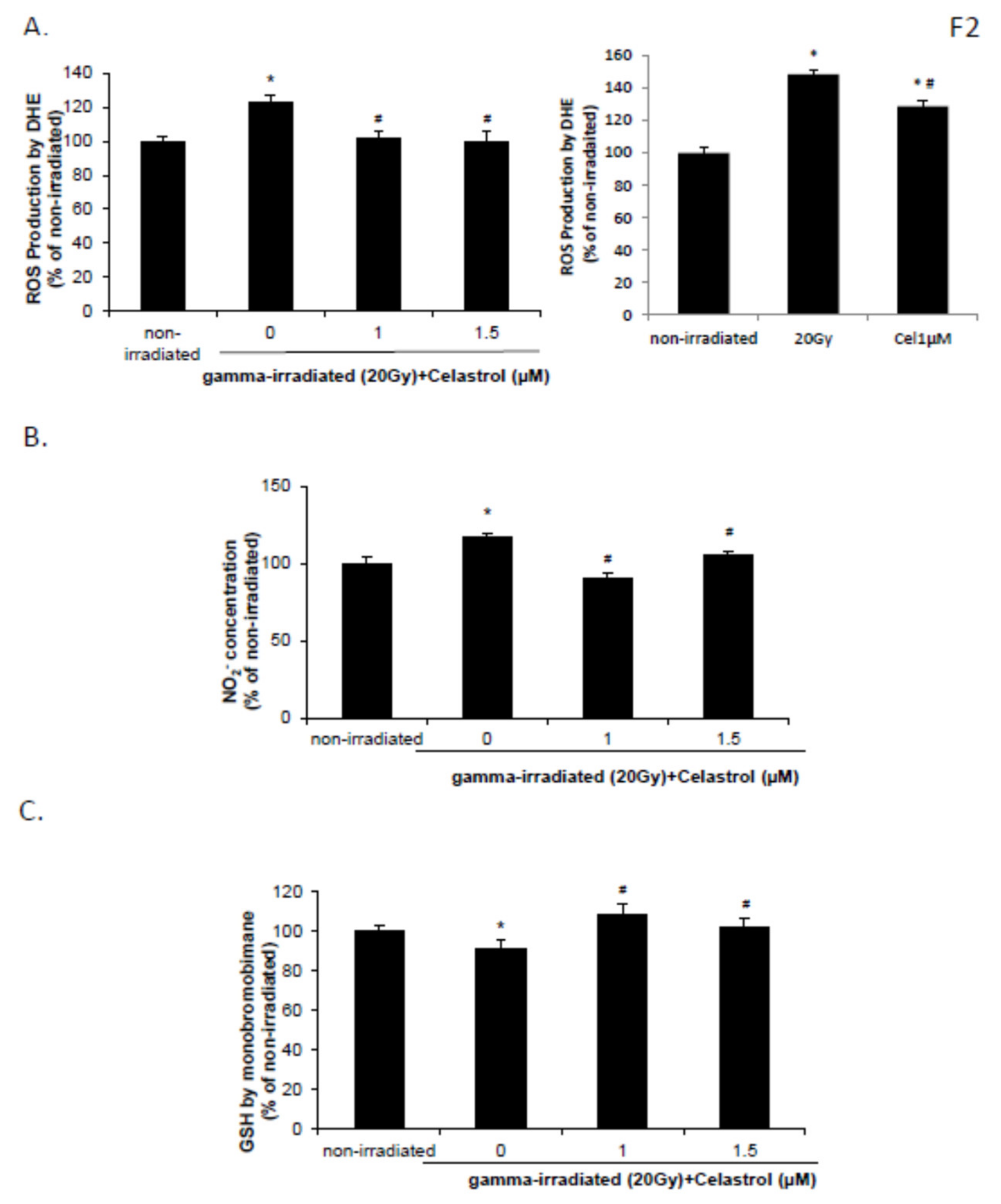
2.2. Celastrol Blocked the Increased Reactive Oxygen Species (ROS) and NO Production and Decreased Glutathione (GSH) Levels Induced by Gamma Irradiation
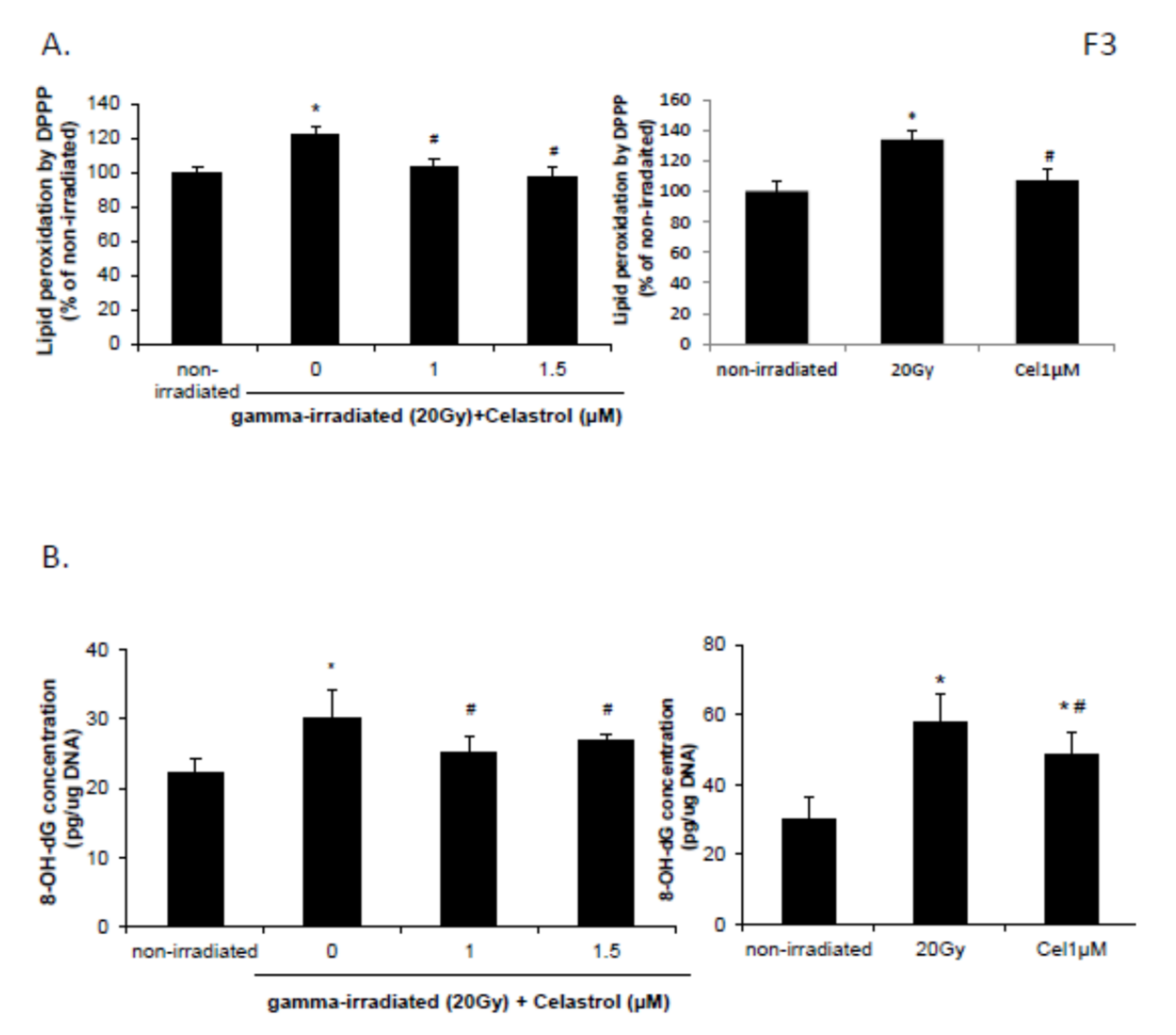
2.3. Celastrol Reversed the Increased Lipid Peroxidation and DNA Oxidative Damage Induced by Gamma Irradiation
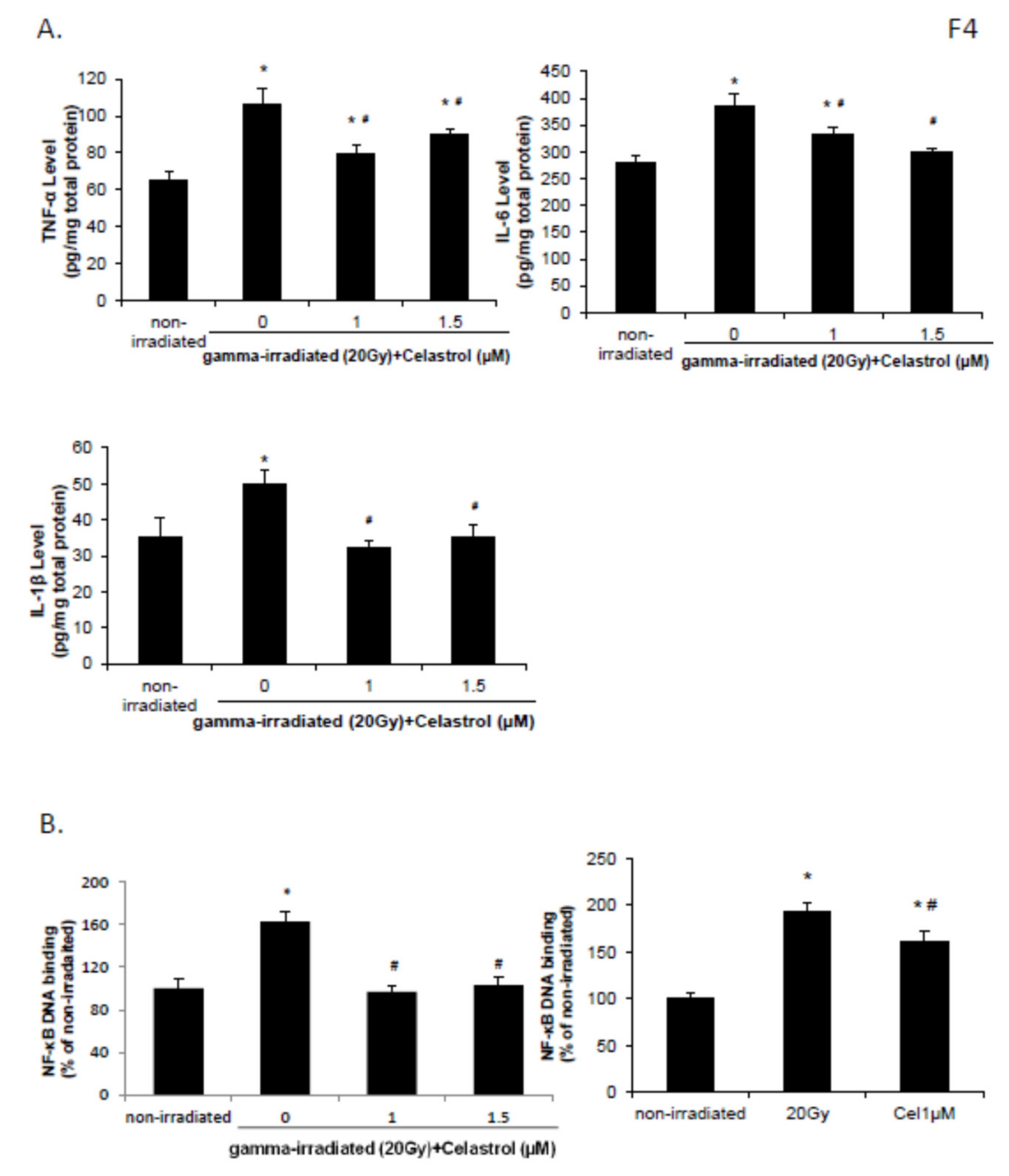
2.4. Celastrol Blocked the Increased Levels of proInflammation Cytokines and the Activation of NF-κB Pathway Induced by Gamma Irradiation
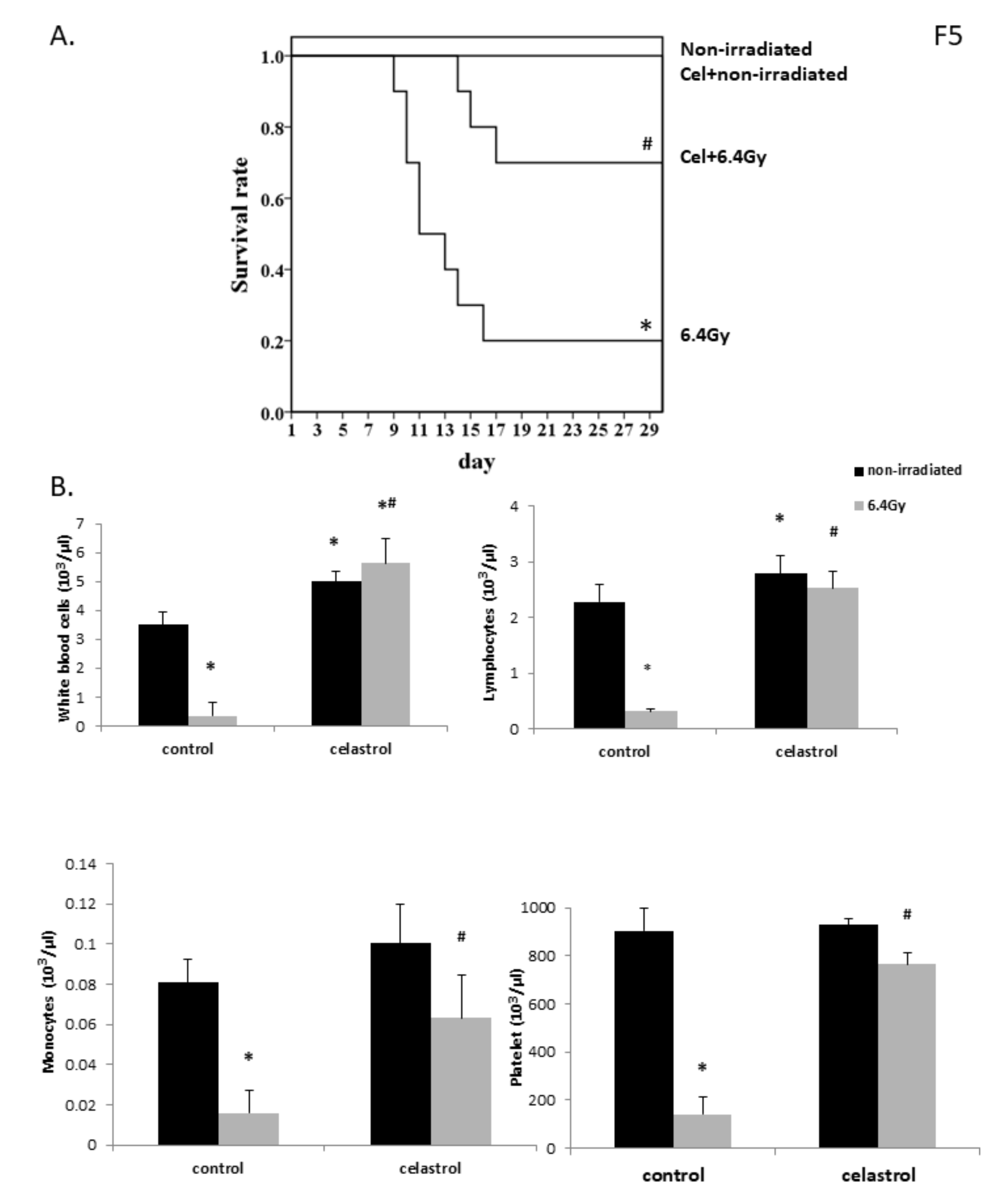
2.5. Celastrol Treatment Significantly Increased Mouse Survival Rate Following Gamma Radiation Exposure
2.6. Celastrol Treatment Exerted Protective Effects in Mice Against Myelosuppression Induced by Gamma Irradiation
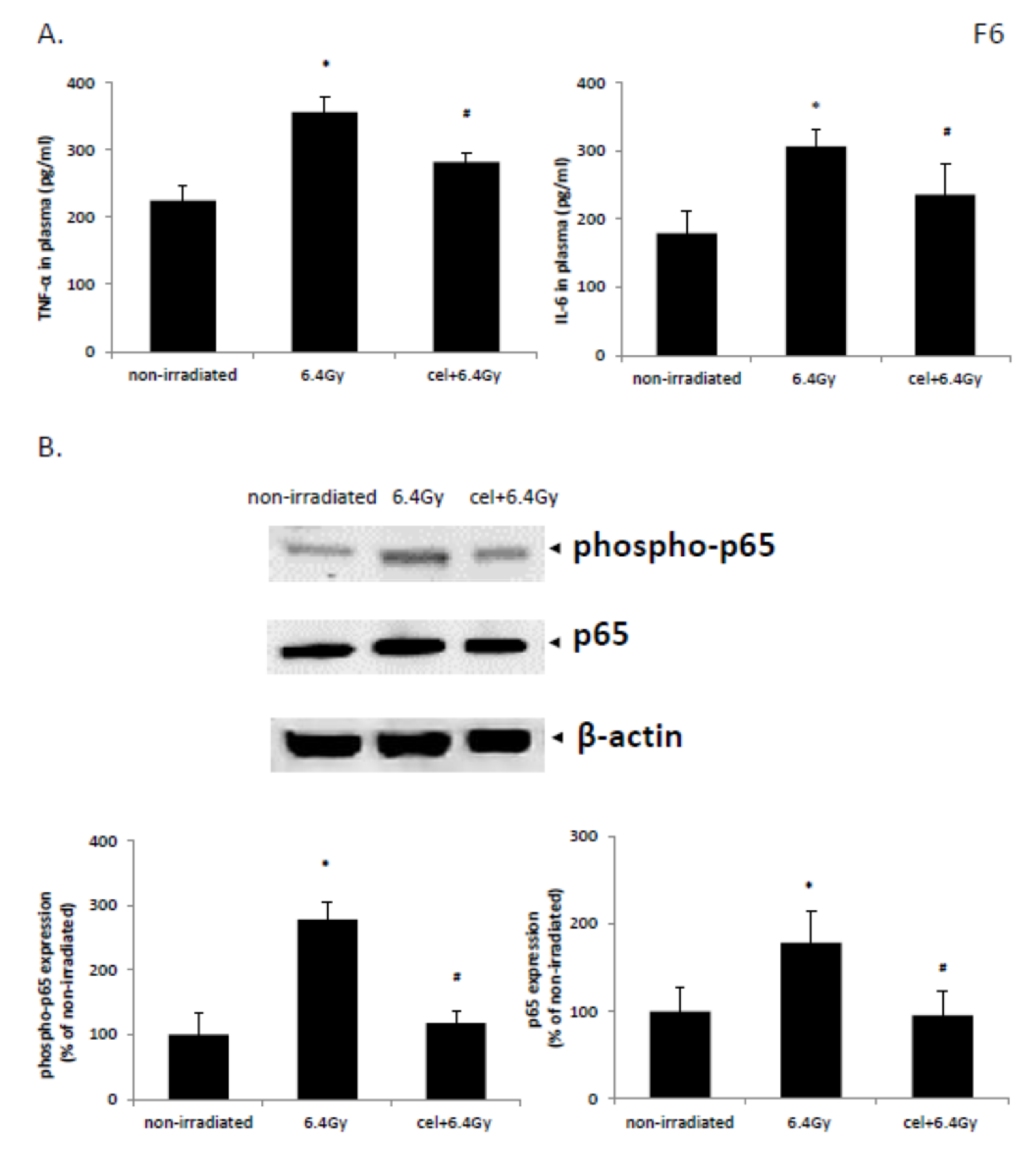
2.7. Celastrol Exhibited Anti-Inflammatory Activities in Animals Following Exposure to Gamma Irradiation
2.8. Celastrol Reversed NF-κB Pathway Activation Induced by Gamma Irradiation in Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines
4.3. Irradiation Procedure and Drug Treatment
4.4. Cell Viability Assay
4.5. Reactive Oxygen Species (ROS), NO, and Glutathione (GSH) Assays
4.6. Lipid Peroxidation Assay
4.7. DNA Oxidative Damage Assay
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Extraction of Nuclear and Cytoplasmic Fraction
4.10. NF-κB DNA Binding Activity Assay
4.11. In Vivo Irradiation Procedure in Mice
4.12. Whole Blood Count
4.13. TNF-α and IL-6 Assayed by ELISA in Animal Plasma
4.14. Western Blotting for Phospho-p65 and p65 in Mouse Colon Tissues
4.15. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ROS | reactive oxygen species |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| DMSO | dimethyl sulfoxide |
| DHE | dihydroethidium |
| SDS | sodium dodecyl sulphate |
| BSA | bovine serum albumin |
| DMEM | Dulbecco’s Modified Eagle Medium |
| FBS | fetal bovine serum |
| DPPP | diphenyl-1-pyrenylphosphine |
| DPPP-O | diphenyl-1-pyrenylphosphine oxide |
| GSH | glutathione |
| mBBr | monobromobimane |
| 8-OH-dG | 8-hydroxy-2-deoxy Guanosine |
| IL-6 | interleukin-6 |
| TNF-α | tumor necrosis factor alpha |
| IL-1β | interleukin-1beta |
| HRP | horseradish peroxidase |
| ELISA | enzyme-linked immunosorbent assay |
| ECL | chemiluminescence |
| IACUC | Institutional Animal Use and Care Committee |
| DIRP | Defence Innovative Research Programme |
References
- Wang, H.; Sethi, G.; Loke, W.K.; Sim, M.K. Des-Aspartate-Angiotensin I Attenuates Mortality of Mice Exposed to Gamma Radiation via a Novel Mechanism of Action. PLoS ONE 2015, 10, e0138009. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Kumar, N.C.; Glickman, R.D. Modulation of photochemical damage in normal and malignant cells by naturally occurring compounds. Photochem. Photobiol. 2012, 88, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Viadero, C.; Jimenez-Sanz, M.; Fernandez-Perez, A.; Verduga Velez, R.; Crespo Santiago, D. Inflammation and oxidation: Predictive and/or causative factors. Rev. Esp. Geriatr. Gerontol. 2016, 51, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Haase, G.M.; Prasad, K.N. Oxidative Damage and Inflammation Biomarkers: Strategy in Hearing Disorders. Otol. Neurotol. 2016, 37, e303–e308. [Google Scholar] [CrossRef]
- Fesenko, S.; Balonov, M.; Prohl, G.; Nakayama, S.; Howard, B.J. A Comparison of Remediation after the Chernobyl and Fukushima Daiichi Accidents. Radiat. Prot. Dosim. 2017, 173, 170–176. [Google Scholar] [CrossRef]
- Ikenoue, T.; Takata, H.; Kusakabe, M.; Kudo, N.; Hasegawa, K.; Ishimaru, T. Temporal variation of cesium isotope concentrations and atom ratios in zooplankton in the Pacific off the east coast of Japan. Sci. Rep. 2017, 7, 39874. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.A. Maintaining a proper perspective of risk associated with radiation exposure. J. Nucl. Med. Technol. 2001, 29, 137–142. [Google Scholar]
- Kannaiyan, R.; Hay, H.S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Vali, S.; Abbasi, T.; Kapoor, S.; Sharma, A.; Kumar, A.P.; et al. Celastrol inhibits proliferation and induces chemosensitization through down-regulation of NF-kappaB and STAT3 regulated gene products in multiple myeloma cells. Br. J. Pharmacol. 2011, 164, 1506–1521. [Google Scholar] [CrossRef] [Green Version]
- Kannaiyan, R.; Manu, K.A.; Chen, L.; Li, F.; Rajendran, P.; Subramaniam, A.; Lam, P.; Kumar, A.P.; Sethi, G. Celastrol inhibits tumor cell proliferation and promotes apoptosis through the activation of c-Jun N-terminal kinase and suppression of PI3 K/Akt signaling pathways. Apoptosis 2011, 16, 1028–1041. [Google Scholar] [CrossRef]
- Kannaiyan, R.; Shanmugam, M.K.; Sethi, G. Molecular targets of celastrol derived from Thunder of God Vine: Potential role in the treatment of inflammatory disorders and cancer. Cancer Lett. 2011, 303, 9–20. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Ahn, K.S.; Lee, J.H.; Kannaiyan, R.; Mustafa, N.; Manu, K.A.; Siveen, K.S.; Sethi, G.; Chng, W.J.; Kumar, A.P. Celastrol Attenuates the Invasion and Migration and Augments the Anticancer Effects of Bortezomib in a Xenograft Mouse Model of Multiple Myeloma. Front. Pharmacol. 2018, 9, 365. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Warrier, S.; Kumar, A.P.; Sethi, G.; Arfuso, F. Potential Role of Natural Compounds as Anti-Angiogenic Agents in Cancer. Curr. Vasc. Pharmacol. 2017, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Li, F.; Shanmugam, M.K.; Kannaiyan, R.; Goh, J.N.; Wong, K.F.; Wang, W.; Khin, E.; Tergaonkar, V.; Kumar, A.P.; et al. Celastrol suppresses growth and induces apoptosis of human hepatocellular carcinoma through the modulation of STAT3/JAK2 signaling cascade in vitro and in vivo. Cancer Prev. Res. 2012, 5, 631–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Ahn, K.S.; Pandey, M.K.; Aggarwal, B.B. Celastrol, a novel triterpene, potentiates TNF-induced apoptosis and suppresses invasion of tumor cells by inhibiting NF-kappaB-regulated gene products and TAK1-mediated NF-kappaB activation. Blood 2007, 109, 2727–2735. [Google Scholar] [CrossRef] [Green Version]
- He, W.Z.; Yin, Z.H.; Gao, J.H.; Ye, Z.Z.; Xie, Y.; Kong, W.H.; Chen, Y.S. Etanercept combined with Tripterygium wilfordii polyglycoside for treatment of rheumatoid arthritis in the elderly: A clinical study. Zhongguo Zhong Xi Yi Jie He Za Zhi Zhongguo Zhongxiyi Jiehe Zazhi = Chin. J. Integr. Tradit. West. Med. 2014, 34, 267–271. [Google Scholar]
- Lv, Q.W.; Zhang, W.; Shi, Q.; Zheng, W.J.; Li, X.; Chen, H.; Wu, Q.J.; Jiang, W.L.; Li, H.B.; Gong, L.; et al. Comparison of Tripterygium wilfordii Hook F with methotrexate in the treatment of active rheumatoid arthritis (TRIFRA): A randomised, controlled clinical trial. Ann. Rheum. Dis. 2015, 74, 1078–1086. [Google Scholar] [CrossRef]
- Jiang, M.; Zha, Q.; Zhang, C.; Lu, C.; Yan, X.; Zhu, W.; Liu, W.; Tu, S.; Hou, L.; Wang, C.; et al. Predicting and verifying outcome of Tripterygium wilfordii Hook F. based therapy in rheumatoid arthritis: From open to double-blinded randomized trial. Sci. Rep. 2015, 5, 9700. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Tang, X.P.; Chen, X.C.; Xiao, H.; Liu, P.; Jiao, J. Will Chinese external therapy with compound Tripterygium wilfordii hook F gel safely control disease activity in patients with rheumatoid arthritis: Design of a double-blinded randomized controlled trial. BMC Complement. Altern. Med. 2017, 17, 444. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zu, Y.; Huang, L.; Yu, J.; Zhao, H.; Wen, C.; Chen, Z.; Xu, Z. Treatment of rheumatoid arthritis with combination of methotrexate and Tripterygium wilfordii: A meta-analysis. Life Sci. 2017, 171, 45–50. [Google Scholar] [CrossRef]
- Wu, C.; Jin, H.Z.; Shu, D.; Li, F.; He, C.X.; Qiao, J.; Yu, X.L.; Zhang, Y.; He, Y.B.; Liu, T.J. Efficacy and safety of Tripterygium wilfordii hook F versus acitretin in moderate to severe psoriasis vulgaris: A randomized clinical trial. Chin. Med. J. 2015, 128, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, X.; Li, H.; Liang, Q.; Chen, J.; Chen, J. Comparison of tripterygium wilfordii multiglycosides and tacrolimus in the treatment of idiopathic membranous nephropathy: A prospective cohort study. BMC Nephrol. 2015, 16, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Gong, Z.; Chen, X.; Tang, L.; Zhao, X.; Yuan, Q.; Cai, G. Tripterygium wilfordii Hook F (a traditional Chinese medicine) for primary nephrotic syndrome. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.X.; Zhao, N.; Zhang, W. The effects and mechanism of Tripterygium wilfordii Hook F combination with irbesartan on urinary podocyte excretion in diabetic nephropathy patients. Zhonghua Nei Ke Za Zhi 2013, 52, 469–473. [Google Scholar]
- Ge, Y.; Xie, H.; Li, S.; Jin, B.; Hou, J.; Zhang, H.; Shi, M.; Liu, Z. Treatment of diabetic nephropathy with Tripterygium wilfordii Hook F extract: A prospective, randomized, controlled clinical trial. J. Transl. Med. 2013, 11, 134. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Shen, X.; Dong, J.; Wang, H.; Zuo, L.; Zhao, J.; Zhu, W.; Li, Y.; Gong, J.; Li, J. Tripterygium wilfordii Hook F as Maintenance Treatment for Crohn’s Disease. Am. J. Med. Sci. 2015, 350, 345–351. [Google Scholar] [CrossRef]
- Zhu, W.; Li, Y.; Gong, J.; Zuo, L.; Zhang, W.; Cao, L.; Gu, L.; Guo, Z.; Li, N.; Li, J. Tripterygium wilfordii Hook. f. versus azathioprine for prevention of postoperative recurrence in patients with Crohn’s disease: A randomized clinical trial. Dig. Liver Dis. 2015, 47, 14–19. [Google Scholar] [CrossRef]
- Pinna, G.F.; Fiorucci, M.; Reimund, J.M.; Taquet, N.; Arondel, Y.; Muller, C.D. Celastrol inhibits pro-inflammatory cytokine secretion in Crohn’s disease biopsies. Biochem. Biophys. Res. Commun. 2004, 322, 778–786. [Google Scholar] [CrossRef]
- Fang, Z.; He, D.; Yu, B.; Liu, F.; Zuo, J.; Li, Y.; Lin, Q.; Zhou, X.; Wang, Q. High-Throughput Study of the Effects of Celastrol on Activated Fibroblast-Like Synoviocytes from Patients with Rheumatoid Arthritis. Genes 2017, 8, 221. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Zhang, M.; Liang, Y.; Wang, D. Celastrol synergizes with oral nifedipine to attenuate hypertension in preeclampsia: A randomized, placebo-controlled, and double blinded trial. J. Am. Soc. Hypertens. 2017, 11, 598–603. [Google Scholar] [CrossRef]
- Cleren, C.; Calingasan, N.Y.; Chen, J.; Beal, M.F. Celastrol protects against MPTP- and 3-nitropropionic acid-induced neurotoxicity. J. Neurochem. 2005, 94, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.C.; Cacabelos, R.; Lombardi, V.R.; Alvarez, X.A.; Vigo, C. Celastrol, a potent antioxidant and anti-inflammatory drug, as a possible treatment for Alzheimer’s disease. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2001, 25, 1341–1357. [Google Scholar] [CrossRef]
- Yu, J.S.; Tseng, C.K.; Lin, C.K.; Hsu, Y.C.; Wu, Y.H.; Hsieh, C.L.; Lee, J.C. Celastrol inhibits dengue virus replication via up-regulating type I interferon and downstream interferon-stimulated responses. Antivir. Res. 2017, 137, 49–57. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, M.H.; Nam, D.H.; Song, H.K.; Kang, Y.S.; Lee, J.E.; Kim, H.W.; Cha, J.J.; Hyun, Y.Y.; Han, S.Y.; et al. Celastrol, an NF-kappaB inhibitor, improves insulin resistance and attenuates renal injury in db/db mice. PLoS ONE 2013, 8, e62068. [Google Scholar] [CrossRef] [Green Version]
- Abu Bakar, M.H.; Cheng, K.K.; Sarmidi, M.R.; Yaakob, H.; Huri, H.Z. Celastrol Protects against Antimycin A-Induced Insulin Resistance in Human Skeletal Muscle Cells. Molecules 2015, 20, 8242–8269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Lee, J.; Salazar Hernandez, M.A.; Mazitschek, R.; Ozcan, U. Treatment of obesity with celastrol. Cell 2015, 161, 999–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Choi, K.J.; Seo, W.D.; Jang, S.Y.; Kim, M.; Lee, B.W.; Kim, J.Y.; Kang, S.; Park, K.H.; Lee, Y.S.; et al. Enhancement of radiation sensitivity in lung cancer cells by celastrol is mediated by inhibition of Hsp90. Int. J. Mol. Med. 2011, 27, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Avilla, J.; Teixido, A.; Velazquez, C.; Alvarenga, N.; Ferro, E.; Canela, R. Insecticidal activity of Maytenus species (Celastraceae) nortriterpene quinone methides against codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). J. Agric. Food Chem. 2000, 48, 88–92. [Google Scholar] [CrossRef]
- Kim, D.H.; Shin, E.K.; Kim, Y.H.; Lee, B.W.; Jun, J.G.; Park, J.H.; Kim, J.K. Suppression of inflammatory responses by celastrol, a quinone methide triterpenoid isolated from Celastrus regelii. Eur. J. Clin. Investig. 2009, 39, 819–827. [Google Scholar] [CrossRef]
- Venkatesha, S.H.; Yu, H.; Rajaiah, R.; Tong, L.; Moudgil, K.D. Celastrus-derived celastrol suppresses autoimmune arthritis by modulating antigen-induced cellular and humoral effector responses. J. Biol. Chem. 2011, 286, 15138–15146. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yuan, Y.; Zhang, Y.; He, Q.; Xu, R.; Ge, F.; Wu, C. Celastrol inhibits IL-1beta-induced inflammation in orbital fibroblasts through the suppression of NF-kappaB activity. Mol. Med. Rep. 2016, 14, 2799–2806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, K.C.; Tsui, K.H.; Chung, L.C.; Yeh, C.N.; Chen, W.T.; Chang, P.L.; Juang, H.H. Celastrol blocks interleukin-6 gene expression via downregulation of NF-kappaB in prostate carcinoma cells. PLoS ONE 2014, 9, e93151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Zhou, S.; Pang, L.; Yang, J.; Li, H.J.; Huo, X.; Qian, S.Y. Celastrol suppresses nitric oxide synthases and the angiogenesis pathway in colorectal cancer. Free Radic. Res. 2019, 53, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xuan, J.; Gu, Y.T.; Shi, K.S.; Xie, J.J.; Chen, J.X.; Zheng, Z.M.; Chen, Y.; Chen, X.B.; Wu, Y.S.; et al. Celastrol reduces IL-1beta induced matrix catabolism, oxidative stress and inflammation in human nucleus pulposus cells and attenuates rat intervertebral disc degeneration in vivo. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 91, 208–219. [Google Scholar] [CrossRef] [PubMed]
- El-Tanbouly, G.S.; El-Awady, M.S.; Megahed, N.A.; Salem, H.A.; El-Kashef, H.A. The NF-kappaB inhibitor celastrol attenuates acute hepatic dysfunction induced by cecal ligation and puncture in rats. Environ. Toxicol. Pharmacol. 2017, 50, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Han, L.P.; Li, C.J.; Sun, B.; Xie, Y.; Guan, Y.; Ma, Z.J.; Chen, L.M. Protective Effects of Celastrol on Diabetic Liver Injury via TLR4/MyD88/NF-kappaB Signaling Pathway in Type 2 Diabetic Rats. J. Diabetes Res. 2016, 2016, 2641248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibuki, Y.; Mizuno, S.; Goto, R. gamma-Irradiation-induced DNA damage enhances NO production via NF-kappaB activation in RAW264.7 cells. Biochim. Biophys. Acta 2003, 1593, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Manna, K.; Das, U.; Das, D.; Kesh, S.B.; Khan, A.; Chakraborty, A.; Dey, S. Naringin inhibits gamma radiation-induced oxidative DNA damage and inflammation, by modulating p53 and NF-kappaB signaling pathways in murine splenocytes. Free Radic. Res. 2015, 49, 422–439. [Google Scholar] [CrossRef]
- Li, F.; Zhang, J.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Kumar, A.P.; Ahn, K.S.; Sethi, G. NF-kappaB in cancer therapy. Arch. Toxicol. 2015, 89, 711–731. [Google Scholar] [CrossRef]
- Li, F.; Sethi, G. Targeting transcription factor NF-kappaB to overcome chemoresistance and radioresistance in cancer therapy. Biochim. Biophys. Acta 2010, 1805, 167–180. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Chaturvedi, M.M.; Aggarwal, B.B. Simvastatin, 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitor, suppresses osteoclastogenesis induced by receptor activator of nuclear factor-kappaB ligand through modulation of NF-kappaB pathway. Int. J. Cancer 2008, 123, 1733–1740. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.S.; Shishodia, S.; Ahn, K.S.; Kunnumakkara, A.B.; Sethi, G.; Aggarwal, B.B. Deguelin, an Akt inhibitor, suppresses IkappaBalpha kinase activation leading to suppression of NF-kappaB-regulated gene expression, potentiation of apoptosis, and inhibition of cellular invasion. J. Immunol. 2006, 177, 5612–5622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, K.S.; Sethi, G.; Jain, A.K.; Jaiswal, A.K.; Aggarwal, B.B. Genetic deletion of NAD(P)H:quinone oxidoreductase 1 abrogates activation of nuclear factor-kappaB, IkappaBalpha kinase, c-Jun N-terminal kinase, Akt, p38, and p44/42 mitogen-activated protein kinases and potentiates apoptosis. J. Biol. Chem. 2006, 281, 19798–19808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S.K.; Aggarwal, R.S.; Sethi, G.; Aggarwal, B.B.; Ramesh, G.T. Morin (3,5,7,2′,4′-Pentahydroxyflavone) abolishes nuclear factor-kappaB activation induced by various carcinogens and inflammatory stimuli, leading to suppression of nuclear factor-kappaB-regulated gene expression and up-regulation of apoptosis. Clin. Cancer Res. 2007, 13, 2290–2297. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Sethi, G.; Aggarwal, B.B. Reversal of chemoresistance and enhancement of apoptosis by statins through down-regulation of the NF-kappaB pathway. Biochem. Pharmacol. 2008, 75, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Chua, A.W.; Hay, H.S.; Rajendran, P.; Shanmugam, M.K.; Li, F.; Bist, P.; Koay, E.S.; Lim, L.H.; Kumar, A.P.; Sethi, G. Butein downregulates chemokine receptor CXCR4 expression and function through suppression of NF-kappaB activation in breast and pancreatic tumor cells. Biochem. Pharmacol. 2010, 80, 1553–1562. [Google Scholar] [CrossRef]
- Siveen, K.S.; Mustafa, N.; Li, F.; Kannaiyan, R.; Ahn, K.S.; Kumar, A.P.; Chng, W.J.; Sethi, G. Thymoquinone overcomes chemoresistance and enhances the anticancer effects of bortezomib through abrogation of NF-kappaB regulated gene products in multiple myeloma xenograft mouse model. Oncotarget 2014, 5, 634–648. [Google Scholar] [CrossRef] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Li, F.; Chen, L.; Siveen, K.S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J. Mol. Med. 2014, 92, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Heinbockel, L.; Su, Q.; Gutsmann, T.; Brandenburg, K.; Weindl, G. Synthetic antimicrobial and LPS-neutralising peptides suppress inflammatory and immune responses in skin cells and promote keratinocyte migration. Sci. Rep. 2016, 6, 31577. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Won, M.; Yuk, J.M.; Park, C.Y.; Byun, H.S.; Park, K.A.; Lee, S.R.; Kang, Y.G.; Shen, H.M.; Lee, I.Y.; et al. IinQ attenuates systemic inflammatory responses via selectively impairing the Myddosome complex formation upon TLR4 ligation. Biochem. Pharmacol. 2016, 121, 52–66. [Google Scholar] [CrossRef]
- Dong, X.R.; Luo, M.; Fan, L.; Zhang, T.; Liu, L.; Dong, J.H.; Wu, G. Corilagin inhibits the double strand break-triggered NF-kappaB pathway in irradiated microglial cells. Int. J. Mol. Med. 2010, 25, 531–536. [Google Scholar] [PubMed]
- Wang, H.; Sim, M.K.; Loke, W.K.; Chinnathambi, A.; Alharbi, S.A.; Tang, F.R.; Sethi, G. Potential Protective Effects of Ursolic Acid against Gamma Irradiation-Induced Damage Are Mediated through the Modulation of Diverse Inflammatory Mediators. Front. Pharmacol. 2017, 8, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuntic, V.S.; Stankovic, M.B.; Vujic, Z.B.; Brboric, J.S.; Uskokovic-Markovic, S.M. Radioprotectors—The evergreen topic. Chem. Biodivers. 2013, 10, 1791–1803. [Google Scholar] [CrossRef] [PubMed]
- Sandeep, D.; Nair, C.K. Radioprotection by alpha-asarone: Prevention of genotoxicity and hematopoietic injury in mammalian organism. Mutat. Res. 2011, 722, 62–68. [Google Scholar] [CrossRef]
- Ran, Y.; Wang, R.; Hasan, M.; Jia, Q.; Tang, B.; Shan, S.; Deng, Y.; Qing, H. Radioprotective effects of dragon’s blood and its extracts on radiation-induced myelosuppressive mice. J. Ethnopharmacol. 2014, 154, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wang, S.; Zhou, P.; Liu, W.; Ni, F. Effects of Ligustrazine on DNA damage and apoptosis induced by irradiation. Environ. Toxicol. Pharmacol. 2013, 36, 1197–1206. [Google Scholar] [CrossRef]
- Luo, Q.; Cui, X.; Yan, J.; Yang, M.; Liu, J.; Jiang, Y.; Li, J.; Zhou, Y. Antagonistic effects of Lycium barbarum polysaccharides on the impaired reproductive system of male rats induced by local subchronic exposure to 60Co-gamma irradiation. Phytother. Res. 2011, 25, 694–701. [Google Scholar] [CrossRef]
- Lata, M.; Prasad, J.; Singh, S.; Kumar, R.; Singh, L.; Chaudhary, P.; Arora, R.; Chawla, R.; Tyagi, S.; Soni, N.L.; et al. Whole body protection against lethal ionizing radiation in mice by REC-2001: A semi-purified fraction of Podophyllum hexandrum. Phytomedicine 2009, 16, 47–55. [Google Scholar] [CrossRef]
- Sankhwar, S.; Gupta, M.L.; Gupta, V.; Verma, S.; Suri, K.A.; Devi, M.; Sharma, P.; Khan, E.A.; Alam, M.S. Podophyllum hexandrum-Mediated Survival Protection and Restoration of Other Cellular Injuries in Lethally Irradiated Mice. Evid. Based Complement. Altern. Med. 2011, 2011, 175140. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Verma, S.; Sankhwar, S.; Flora, S.J.; Gupta, M.L. Bioavailability, antioxidant and non toxic properties of a radioprotective formulation prepared from isolated compounds of Podophyllum hexandrum: A study in mouse model. Cell. Mol. Biol. 2012, 58, Ol1646–Ol1653. [Google Scholar]
- Saini, R.; Verma, S.; Singh, A.; Lata Gupta, M. Role of Active Principles of Podophyllum hexandrum in Amelioration of Radiation Mediated Lung Injuries by Reactive Oxygen/Nitrogen Species Reduction. CellBio 2013, 02, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, P.; Shukla, S.K.; Sharma, R.K. REC-2006-A Fractionated Extract of Podophyllum hexandrum Protects Cellular DNA from Radiation-Induced Damage by Reducing the Initial Damage and Enhancing Its Repair In Vivo. Evid. Based Complement. Altern. Med. 2011, 2011, 473953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandeep, D.; Nair, C.K. Protection of DNA and membrane from gamma-radiation induced damage by the extract of Acorus calamus Linn.: An in vitro study. Environ. Toxicol. Pharmacol. 2010, 29, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Sandeep, D.; Nair, C.K. Protection from lethal and sub-lethal whole body exposures of mice to gamma-radiation by Acorus calamus L.: Studies on tissue antioxidant status and cellular DNA damage. Exp. Toxicol. Pathol. 2012, 64, 57–64. [Google Scholar] [CrossRef] [PubMed]
- El-Desouky, W.; Hanafi, A.; Abbas, M.M. Radioprotective effect of green tea and grape seed extracts mixture on gamma irradiation induced immune suppression in male albino rats. Int. J. Radiat. Biol. 2016, 93, 433–439. [Google Scholar] [CrossRef]
- Ismail, A.F.; Salem, A.A.; Eassawy, M.M. Modulation of gamma-irradiation and carbon tetrachloride induced oxidative stress in the brain of female rats by flaxseed oil. J. Photochem. Photobiol. 2016, 161, 91–99. [Google Scholar] [CrossRef]
- Dai, Y.; DeSano, J.T.; Meng, Y.; Ji, Q.; Ljungman, M.; Lawrence, T.S.; Xu, L. Celastrol potentiates radiotherapy by impairment of DNA damage processing in human prostate cancer. Int. J. Radiat. Oncol. Biol. Phys. 2009, 74, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Stankova, K.; Ivanova, K.; Nikolov, V.; Aneva, N.; Georgieva, R.; Boteva, R. Proteasome inhibition protects human peripheral blood mononuclear cells from radiation-induced oxidative stress. Int. J. Radiat. Biol. 2013, 89, 493–500. [Google Scholar] [CrossRef]
- Han, X.B.; Tan, Y.; Fang, Y.Q.; Li, F. Protective effects of celastrol against gamma irradiation-induced oxidative stress in human umbilical vein endothelial cells. Exp. Ther. Med. 2018, 16, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Aravindan, N.; Xu, J.; Natarajan, M. Inter- and Intra-cellular Mechanism of NF-kB-dependent Survival Advantage and Clonal Expansion of Radio-resistant Cancer Cells. Cell. Signal. 2017, 31, 105–111. [Google Scholar] [CrossRef]
- Lee, Y.H.; Wang, E.; Kumar, N.; Glickman, R.D. Ursolic acid differentially modulates apoptosis in skin melanoma and retinal pigment epithelial cells exposed to UV-VIS broadband radiation. Apoptosis 2014, 19, 816–828. [Google Scholar] [CrossRef] [PubMed]
- Das, U.; Manna, K.; Sinha, M.; Datta, S.; Das, D.K.; Chakraborty, A.; Ghosh, M.; Saha, K.D.; Dey, S. Role of ferulic acid in the amelioration of ionizing radiation induced inflammation: A murine model. PLoS ONE 2014, 9, e97599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Adhikari, J.S.; Rizvi, M.A.; Chaudhury, N.K. Radioprotective potential of melatonin against 60Co gamma-ray-induced testicular injury in male C57BL/6 mice. J. Biomed. Sci. 2015, 22, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindekov, I.; Mileva, M.; Krastev, D.; Vassilieva, V.; Raynova, Y.; Doumanova, L.; Aljakov, M.; Idakieva, K. Radioprotective effect of Rapana thomasiana hemocyanin in gamma induced acute radiation syndrome. Biotechnol. Biotechnol. Equip. 2014, 28, 533–539. [Google Scholar] [CrossRef]
- Hu, J.; Yang, Z.; Wang, J.; Tang, Y.; Liu, H.; Zhang, B.; Chen, H. Infusion of Trx-1-overexpressing hucMSC prolongs the survival of acutely irradiated NOD/SCID mice by decreasing excessive inflammatory injury. PLoS ONE 2013, 8, e78227. [Google Scholar] [CrossRef]
- Mortazavi, S.M.; Rahimi, S.; Mosleh-Shirazi, M.A.; Arjomandi, M.; Soleimani, A.; Koohi Hossein-Abadi, O.; Haghani, M.; Alavi, M. A Comparative Study on the Life-Saving Radioprotective Effects of Vitamins A, E, C and Over-the-Counter Multivitamins. J. Biomed. Phys. Eng. 2015, 5, 59–66. [Google Scholar]
- Booth, C.; Tudor, G.; Tudor, J.; Katz, B.P.; MacVittie, T.J. Acute gastrointestinal syndrome in high-dose irradiated mice. Health Phys. 2012, 103, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.M.; Day, R.; Singh, V.K. New approaches to radiation protection. Front. Oncol. 2014, 4, 381. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.P.; Brown, S.L.; Georges, G.E.; Hauer-Jensen, M.; Hill, R.P.; Huser, A.K.; Kirsch, D.G.; Macvittie, T.J.; Mason, K.A.; Medhora, M.M.; et al. Animal models for medical countermeasures to radiation exposure. Radiat. Res. 2010, 173, 557–578. [Google Scholar] [CrossRef] [Green Version]
- Kusy, S.; Ghosn, E.E.; Herzenberg, L.A.; Contag, C.H. Development of B cells and erythrocytes is specifically impaired by the drug celastrol in mice. PLoS ONE 2012, 7, e35733. [Google Scholar] [CrossRef]
- Venkatesha, S.H.; Dudics, S.; Astry, B.; Moudgil, K.D. Control of autoimmune inflammation by celastrol, a natural triterpenoid. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Shan, J.; Chen, X.; Li, S.; Long, D.; Li, Y. Celastrol mediates Th17 and Treg cell generation via metabolic signaling. Biochem. Biophys. Res. Commun. 2018, 497, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Astry, B.; Venkatesha, S.H.; Laurence, A.; Christensen-Quick, A.; Garzino-Demo, A.; Frieman, M.B.; O’Shea, J.J.; Moudgil, K.D. Celastrol, a Chinese herbal compound, controls autoimmune inflammation by altering the balance of pathogenic and regulatory T cells in the target organ. Clin. Immunol. 2015, 157, 228–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdin, A.A.; Hasby, E.A. Modulatory effect of celastrol on Th1/Th2 cytokines profile, TLR2 and CD3+ T-lymphocyte expression in a relapsing-remitting model of multiple sclerosis in rats. Eur. J. Pharmacol. 2014, 742, 102–112. [Google Scholar] [CrossRef]
- Sinha, M.; Das, D.K.; Manna, K.; Datta, S.; Ray, T.; Sil, A.K.; Dey, S. Epicatechin ameliorates ionising radiation-induced oxidative stress in mouse liver. Free Radic. Res. 2012, 46, 842–849. [Google Scholar] [CrossRef]
- Brach, M.A.; Hass, R.; Sherman, M.L.; Gunji, H.; Weichselbaum, R.; Kufe, D. Ionizing radiation induces expression and binding activity of the nuclear factor kappa B. J. Clin. Investig. 1991, 88, 691–695. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Ahn, K.S.; Alharbi, S.A.; Shair, O.H.M.; Arfuso, F.; Sethi, G.; Chinnathambi, A.; Tang, F.R. Celastrol Alleviates Gamma Irradiation-Induced Damage by Modulating Diverse Inflammatory Mediators. Int. J. Mol. Sci. 2020, 21, 1084. https://doi.org/10.3390/ijms21031084
Wang H, Ahn KS, Alharbi SA, Shair OHM, Arfuso F, Sethi G, Chinnathambi A, Tang FR. Celastrol Alleviates Gamma Irradiation-Induced Damage by Modulating Diverse Inflammatory Mediators. International Journal of Molecular Sciences. 2020; 21(3):1084. https://doi.org/10.3390/ijms21031084
Chicago/Turabian StyleWang, Hong, Kwang Seok Ahn, Sulaiman Ali Alharbi, Omar H. M. Shair, Frank Arfuso, Gautam Sethi, Arunachalam Chinnathambi, and Feng Ru Tang. 2020. "Celastrol Alleviates Gamma Irradiation-Induced Damage by Modulating Diverse Inflammatory Mediators" International Journal of Molecular Sciences 21, no. 3: 1084. https://doi.org/10.3390/ijms21031084