Effects of Helioxanthin Derivative-Treated Human Dental Pulp Stem Cells on Fracture Healing


Abstract
:1. Introduction
2. Results
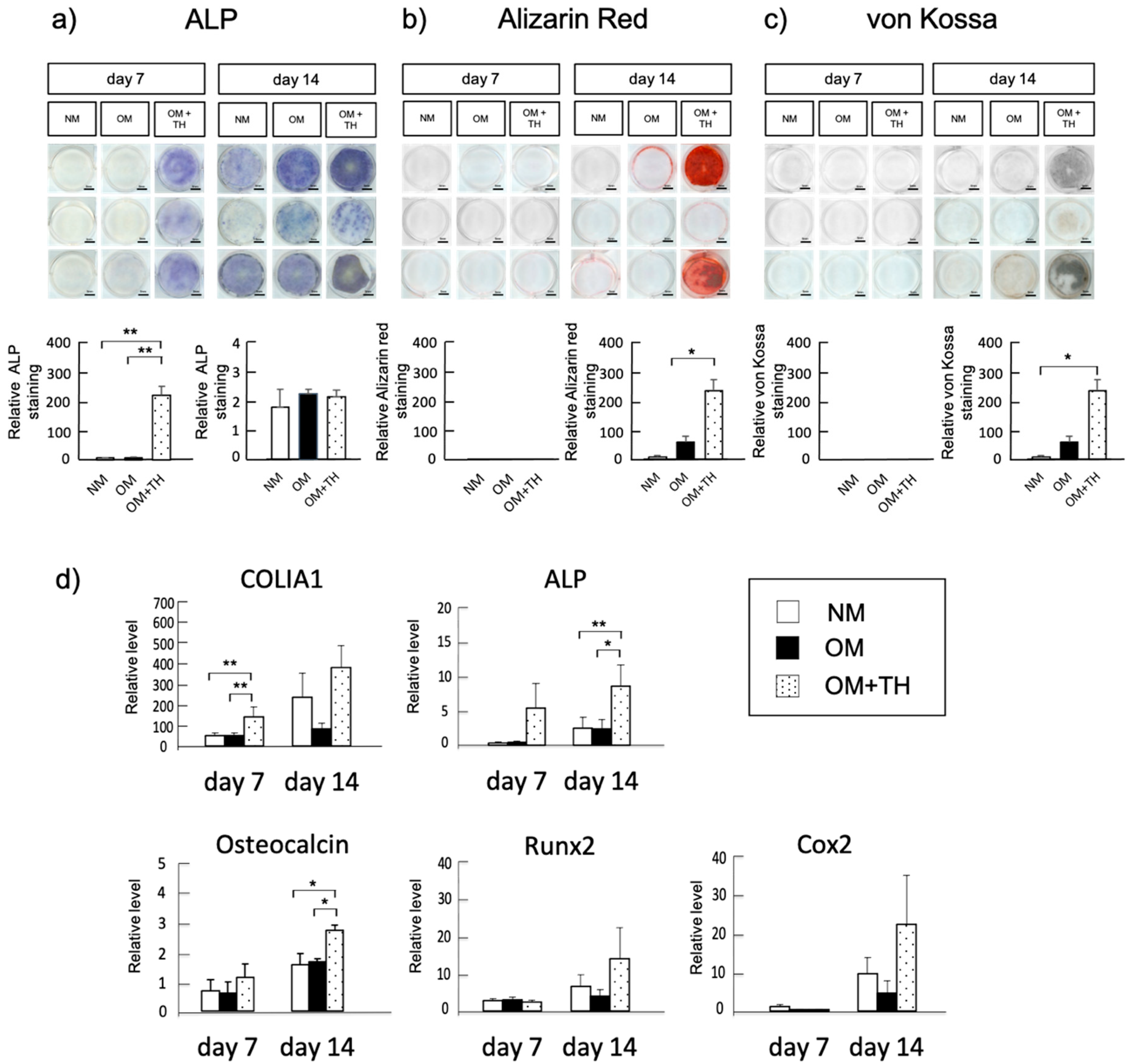
2.1. Osteogenic Differentiation of DPSCs Treated with TH in Short-Term Culture
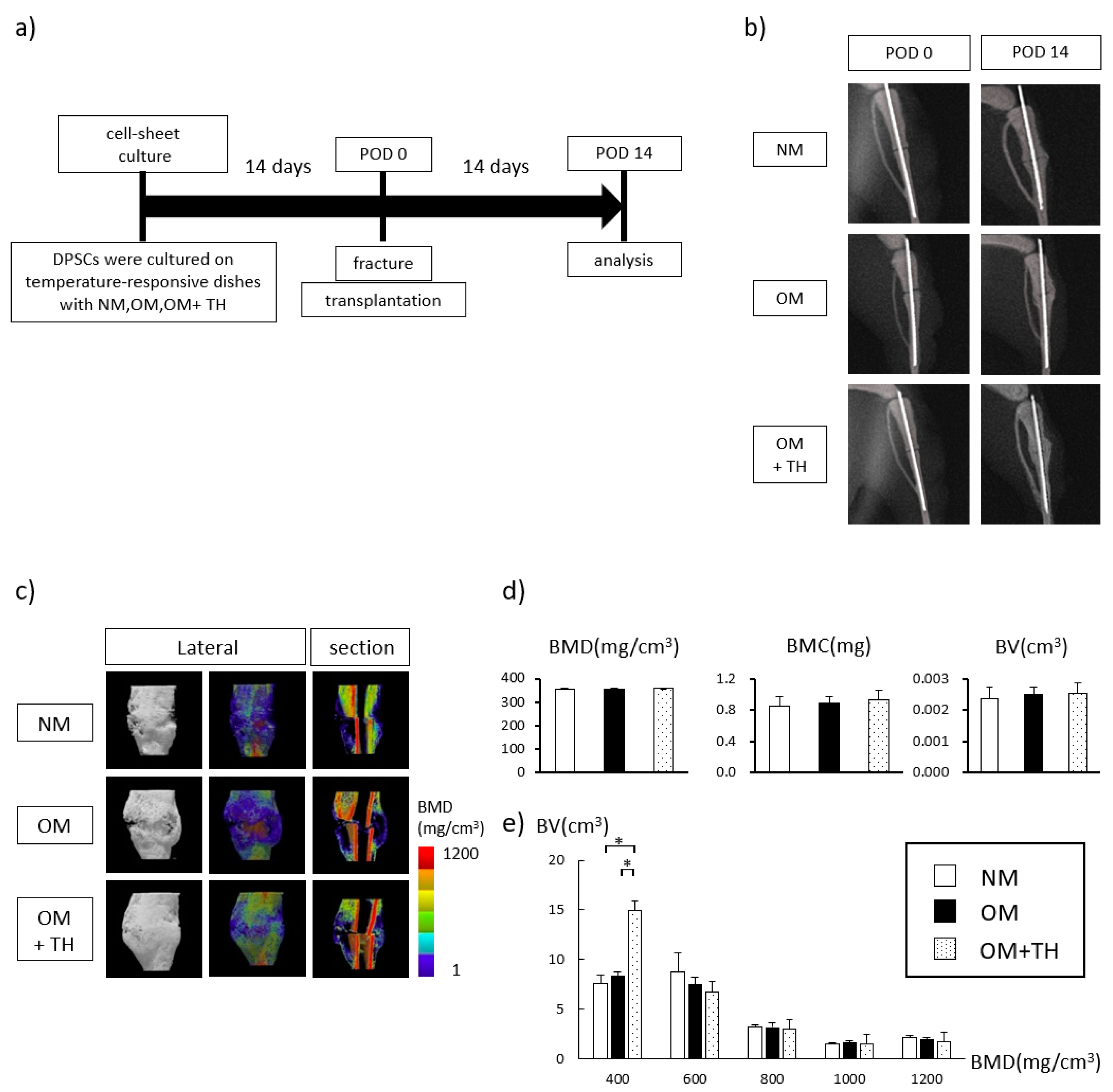
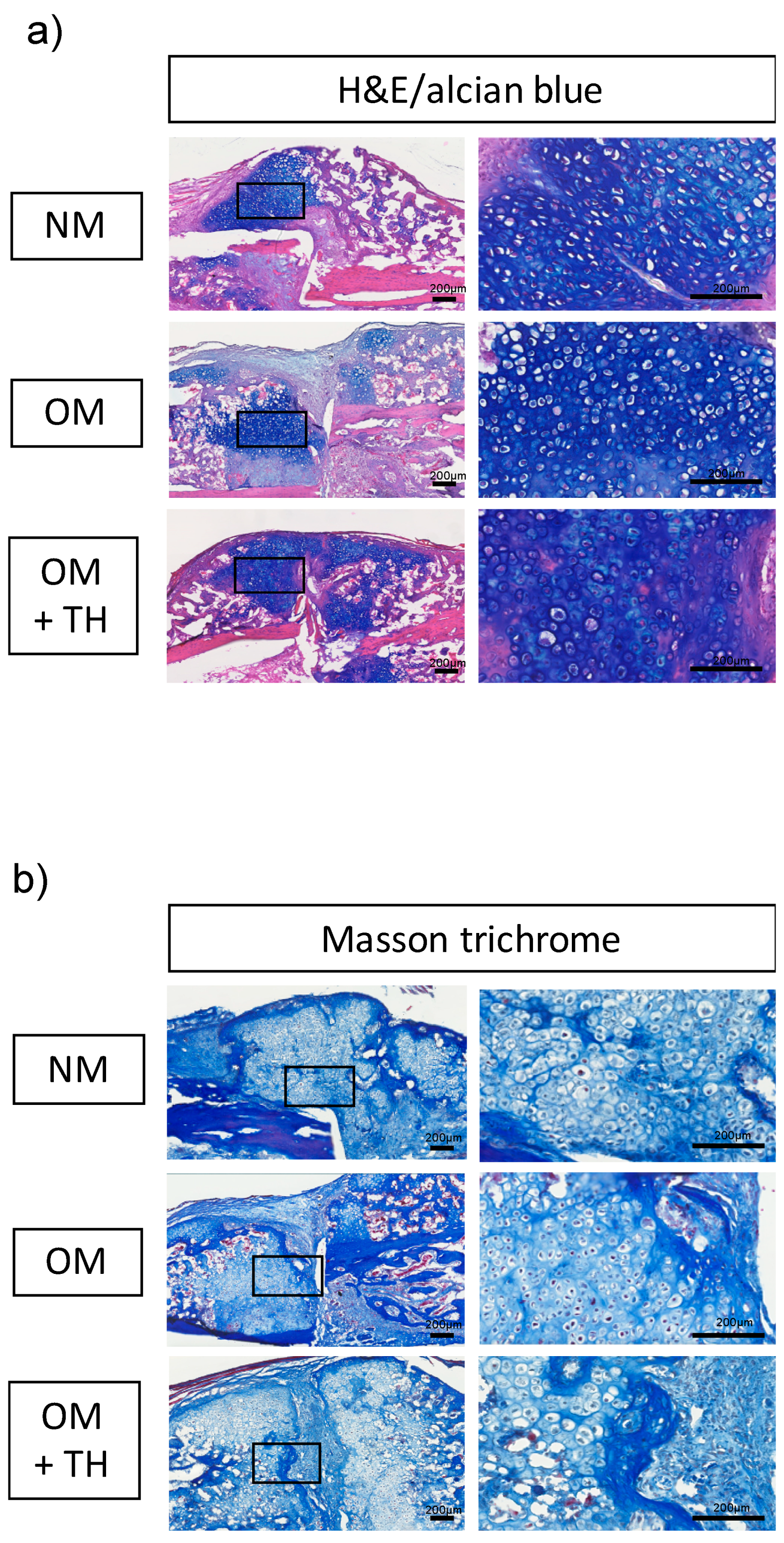
2.2. Fracture Healing by Transplantation of OM+TH-Treated DPSCs In Vivo
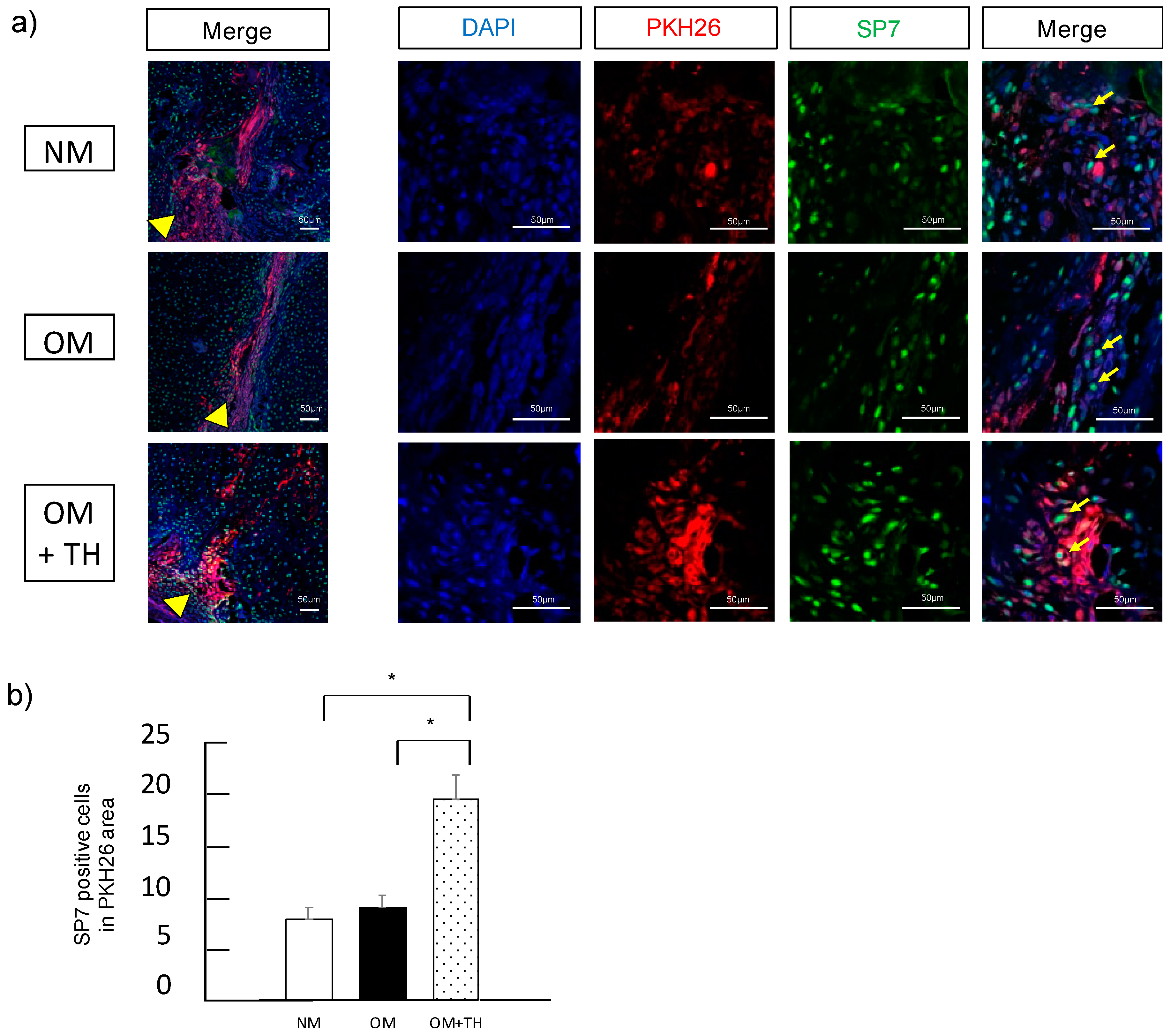
2.3. Functions of Transplanted DPSCs
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Alkaline Phosphatase Staining
4.3. Alizarin Red S Staining
4.4. Von Kossa Staining
4.5. Real-Time Reverse Transcription-Polymerase Chain Reaction Analysis
4.6. Transplantation of a DPSC Sheet into a Mouse Tibia Fracture Model
4.7. Radiological Analysis
4.8. Histological Analysis
4.9. Immunofluorescence Staining
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase |
| BMC | bone mineral content |
| BMD | bone mineral density |
| BMP | bone morphogenic protein |
| BMSCs | bone marrow stem cells |
| BV | Bone volume |
| COLIA1 | Type I collagen alpha 1 |
| DPSCs | human dental pulp stem cells |
| FBS | fetal bovine serum |
| MSCs | mesenchymal stem cells |
| NM | normal medium |
| OM | osteogenic medium |
| PSA | penicillin/streptomycin/amphotericin B |
| PBS | phosphate-buffered saline |
| SEM | standard error of the mean |
| TH | 4-(4-methoxyphenyl) pyrido[40,30:4,5]thieno[2,3-b]pyridine-2-carboxamide |
| αMEM | alpha-modified Eagle’s medium |
References
- Ferguson, C.; Alpern, E.; Miclau, T.; Helms, J.A. Does adult fracture repair reca- pitulate embryonic skeletal formation? Mech. Dev. 1999, 87, 57–66. [Google Scholar] [CrossRef]
- Hall, B.K.; Miyake, T. All for one and one for all: Condensations and the initiation of skeletal development. Bioessays 2000, 22, 138–147. [Google Scholar] [CrossRef]
- Dunlop, L.L.; Hall, B.K. Relationships between cellular condensation, preosteoblast formation and epithelial-mesenchymal interactions in initiation of osteogenesis. Int. J. Dev. Biol. 1995, 39, 357–371. [Google Scholar] [PubMed]
- Shen, X.; Wan, C.; Ramaswamy, G.; Mavalli, M.; Wang, Y.; Duvall, C.L.; Deng, L.F.; Guldberg, R.E.; Eberhart, A.; Clemens, T.L.; et al. Prolyl hydroxylase inhibitors increase neoangiogenesis and callus formation following femur fracture in mice. J. Orthop. Res. 2009, 27, 1298–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem Cell Properties of Human Dental Pulp Stem Cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaki, Y.; Nakahara, T.; Ishikawa, H.; Sato, S. In vitro analysis of mesenchymal stem cells derived from human teeth and bone marrow. Odontology 2012, 101, 121–132. [Google Scholar] [CrossRef]
- Kawamura, M.; Miyagawa, S.; Fukushima, S.; Saito, A.; Miki, K.; Ito, E.; Sougawa, N.; Daimon, T.; Shimizu, T.; Okano, T.; et al. Enhanced Survival of Transplanted Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes by the Combination of Cell Sheets with the Pedicled Omental Flap Technique in a Porcine Heart. Circulation 2013, 128, S87–S94. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Yamato, M.; Hayashida, Y.; Watanabe, K.; Yamamoto, K.; Adachi, E.; Nagai, S.; Kikuchi, A.; Naoyuki, M.; Watanabe, H.; et al. Corneal Reconstruction with Tissue-Engineered Cell Sheets Composed of Autologous Oral Mucosal Epithelium. N. Engl. J. Med. 2004, 351, 1187–1196. [Google Scholar] [CrossRef] [Green Version]
- Ohki, T.; Yamato, M.; Ota, M.; Takagi, R.; Murakami, D.; Kondo, M.; Sasaki, R.; Namiki, H.; Okano, T.; Yamamoto, M. Prevention of Esophageal Stricture After Endoscopic Submucosal Dissection Using Tissue-Engineered Cell Sheets. Gastroenterology 2012, 143, 582–588.e2. [Google Scholar] [CrossRef]
- Iwata, T.; Yamato, M.; Tsuchioka, H.; Takagi, R.; Mukobata, S.; Washio, K.; Okano, T.; Ishikawa, I. Periodontal regeneration with multi-layered periodontal ligament-derived cell sheets in a canine model. Biomaterials 2009, 30, 2716–2723. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Jung, T.; Shahid, M.A.; Khan, I.U.; Kim, W.H.; Kweon, O.-K. Frozen-thawed gelatin-induced osteogenic cell sheets of canine adipose-derived mesenchymal stromal cells improved fracture healing in canine model. J. Veter Sci. 2019, 20, e63. [Google Scholar] [CrossRef] [PubMed]
- Hirose, M.; Kwon, O.H.; Yamato, M.; Kikuchi, A.; Okano, T. Creation of Designed Shape Cell Sheets That Are Noninvasively Harvested and Moved onto Another Surface. Biomacromolecules 2000, 1, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Lavery, K.; Swain, P.; Falb, D.; Alaoui-Ismaili, M.H. BMP-2/4 and BMP-6/7 differentially utilize cell surface receptors to induce osteoblastic differentiation of human bone marrow-derived mesenchymal stem cells. J. Biol. Chem. 2008, 283, 20948–20958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, Y.; Kawase-Koga, Y.; Hojo, H.; Yano, F.; Sato, M.; Chung, U.-I.; Ohba, S.; Chikazu, D. Bone regeneration by human dental pulp stem cells using a helioxanthin derivative and cell-sheet technology. Stem Cell Res. Ther. 2018, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Suen, P.K.; He, Y.-X.; Chow, D.H.K.; Huang, L.; Li, C.; Ke, H.Z.; Ominsky, M.S.; Qin, L. Sclerostin monoclonal antibody enhanced bone fracture healing in an open osteotomy model in rats. J. Orthop. Res. 2014, 32, 997–1005. [Google Scholar] [CrossRef]
- Murray, P.E.; Stanley, H.R.; Matthews, J.B.; Sloan, A.J.; Smith, A.J. Age-related odontometric changes of human teeth. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2002, 93, 474–482. [Google Scholar] [CrossRef]
- Alipour, M.; Aghazadeh, M.; Akbarzadeh, A.; Vafajoo, Z.; Aghazadeh, Z.; Hokmabad, V.R. Towards osteogenic differentiation of human dental pulp stem cells on PCL-PEGPCL/zeolite nanofibrous scaffolds. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3431–3437. [Google Scholar] [CrossRef] [Green Version]
- Notoya, K.; Nagai, H.; Oda, T.; Gotoh, M.; Hoshino, T.; Muranishi, H.; Taketomi, S.; Sohda, T.; Makino, H. Enhancement of osteogenesis in vitro and in vivo by a novel osteoblast differentiation promoting compound, TAK-778. J. Pharmacol. Exp. Ther. 1999, 290, a008334. [Google Scholar]
- Long, F.; Ornitz, D.M. Development of the Endochondral Skeleton. Cold Spring Harb. Perspect. Boil. 2013, 5, a008334. [Google Scholar] [CrossRef] [Green Version]
- Yasui, K.; Komiyama, A. New clinical applications of xanthine derivatives: Modulatory actions on leukocyte survival and function. Int. J. Hematol. 2001, 73, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ohba, S.; Nakajima, K.; Komiyama, Y.; Kugimiya, F.; Igawa, K.; Itaka, K.; Moro, T.; Nakamura, K.; Kawaguchi, H.; Takato, T.; et al. A novel osteogenic helioxanthin-derivative acts in a BMP-dependent manner. Biochem. Biophys. Res. Commun. 2007, 357, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Komiyama, Y.; Hojo, H.; Ohba, S.; Yano, F.; Nishikawa, N.; Aburatani, H.; Takato, T.; Chung, U.-I. Enhancement of bone formation ex vivo and in vivo by a helioxanthin-derivative. Biochem. Biophys. Res. Commun. 2010, 395, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Amano, H.; Iwaki, F.; Oki, M.; Aoki, K.; Ohba, S. An osteogenic helioxanthin derivative suppresses the formation of bone-resorbing osteoclasts. Regen. Ther. 2019, 11, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Chikazu, D.; Li, X.; Kawaguchi, H.; Sakuma, Y.; Vozesensky, O.S.; Adams, D.J.; Xu, M.; Hoshi, K.; Katavic, V.; Herschman, H.R.; et al. Bone Morphogenetic Protein 2 Induces Cyclo-oxygenase 2in Osteoblasts via a Cbfa1 Binding Site: Role in Effects ofBone Morphogenetic Protein 2 In Vitro and In Vivo. J. Bone Miner. Res. 2002, 8, 1430–1439. [Google Scholar] [CrossRef]
- Hinton, R.; Jing, Y.; Jing, J.; Feng, J. Roles of Chondrocytes in Endochondral Bone Formation and Fracture Repair. J. Dent. Res. 2016, 96, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular biology of fracture healing. J. Orthop. Res. 2019, 37, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Undale, A.; Fraser, D.; Hefferan, T.; Kopher, R.A.; Herrick, J.; Evans, G.L.; Li, X.; Kakar, S.; Hayes, M.; Atkinson, E.; et al. Induction of fracture repair by mesenchymal cells derived from human embryonic stem cells or bone marrow. J. Orthop. Res. 2011, 29, 1804–1811. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, C.; Gou, W.-L.; Xu, X.-L.; Wang, Y.; Wang, A.-Y.; Xu, W.; Hao, C.; Liu, S.; Lu, Q.; et al. The optimal time to inject bone mesenchymal stem cells for fracture healing in a murine model. Stem Cell Res. Ther. 2018, 9, 272. [Google Scholar] [CrossRef]
- Jiang, X.; Xu, C.; Shi, H.; Cheng, Q. PTH1-34 improves bone healing by promoting angiogenesis and facilitating MSCs migration and differentiation in a stabilized fracture mouse model. PLoS ONE 2019, 14, e0226163. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, B.; Feng, Y.; Shu, L.; Cao, X.; Karaplis, A.; Goltzman, D.; Miao, D. Endogenous PTH Deficiency Impairs Fracture Healing and Impedes the Fracture-Healing Efficacy of Exogenous PTH(1-34). PLoS ONE 2011, 6, e23060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierdomenico, L.; Bonsi, L.; Calvitti, M.; Rondelli, D.; Arpinati, M.; Chirumbolo, G.; Becchetti, E.; Marchionni, C.; Alviano, F.; Fossati, V.; et al. Multipotent Mesenchymal Stem Cells with Immunosuppressive Activity Can Be Easily Isolated from Dental Pulp. Transplantion 2005, 80, 836–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Ito, K.; Nakamura, S.; Ueda, M.; Nagasaka, T. Promising Cell-Based Therapy for Bone Regeneration Using Stem Cells from Deciduous Teeth, Dental Pulp, and Bone Marrow. Cell Transplant. 2011, 20, 1003–1013. [Google Scholar] [CrossRef]
- Ogasawara, T.; Kawaguchi, H.; Jinno, S.; Hoshi, K.; Itaka, K.; Takato, T.; Nakamura, K.; Okayama, H. Bone Morphogenetic Protein 2-Induced Osteoblast Differentiation Requires Smad-Mediated Down-Regulation of Cdk6. Mol. Cell. Biol. 2004, 24, 6560–6568. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, M.; Hojo, H.; Kitaura, Y.; Maeda, Y.; Aini, H.; Takato, T.; Chung, U.; Ohba, S. Local administration of a hedgehog agonist accelerates fracture healing in a mouse model. Biochem. Biophys. Res. Commun. 2016, 479, 772–778. [Google Scholar] [CrossRef]
- Kitaura, Y.; Hojo, H.; Komiyama, Y.; Takato, T.; Chung, U.-I.; Ohba, S. Gli1 Haploinsufficiency Leads to Decreased Bone Mass with an Uncoupling of Bone Metabolism in Adult Mice. PLoS ONE 2014, 9, e109597. [Google Scholar] [CrossRef]
- Ogi, H.; Suzuki, K.; Ogino, Y.; Kamimura, M.; Miyado, M.; Ying, X.; Zhang, Z.; Shinohara, M.; Chen, Y.; Yamada, G. Ventral abdominal wall dysmorphogenesis ofMsx1/Msx2 double-mutant mice. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2005, 284, 424–430. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (Forward and Reverse, 5′-3′) | Accession |
|---|---|---|
| GAPDH | GAAGGTGAAGGTCGGAGTCA | BC023632 |
| GAAGATGGTGATGGGATTTC | ||
| COLIA1 | GTGCTAAAGGTGCCAATGGT | NM_000088 |
| CTCCTCGCTTTCCTTCCTCT | ||
| ALP | ATGAAGGAAAAGCCAAGCAG | NM_000478 |
| ATGGAGACATTCTCTCGTTC | ||
| Osteocalcin | GGCAGCGAGGTAGTGAAGAG | NM_199173 |
| AGCAGAGCGACACCCTAGAC | ||
| Runx2 | CAGACCAGCAGCACTCCATA | NM_004348 |
| CAGCGTCAACACCATCATTC | ||
| Cox2 | CCTCCTGTGCCTGATGATTGC | M90100 |
| TGGCCCTCGCTTATGATCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamakawa, D.; Kawase-Koga, Y.; Fujii, Y.; Kanno, Y.; Sato, M.; Ohba, S.; Kitaura, Y.; Kashiwagi, M.; Chikazu, D. Effects of Helioxanthin Derivative-Treated Human Dental Pulp Stem Cells on Fracture Healing. Int. J. Mol. Sci. 2020, 21, 9158. https://doi.org/10.3390/ijms21239158
Yamakawa D, Kawase-Koga Y, Fujii Y, Kanno Y, Sato M, Ohba S, Kitaura Y, Kashiwagi M, Chikazu D. Effects of Helioxanthin Derivative-Treated Human Dental Pulp Stem Cells on Fracture Healing. International Journal of Molecular Sciences. 2020; 21(23):9158. https://doi.org/10.3390/ijms21239158
Chicago/Turabian StyleYamakawa, Daiki, Yoko Kawase-Koga, Yasuyuki Fujii, Yuki Kanno, Marika Sato, Shinsuke Ohba, Yoshiaki Kitaura, Miki Kashiwagi, and Daichi Chikazu. 2020. "Effects of Helioxanthin Derivative-Treated Human Dental Pulp Stem Cells on Fracture Healing" International Journal of Molecular Sciences 21, no. 23: 9158. https://doi.org/10.3390/ijms21239158