Peptides Derived from (RRWQWRMKKLG)2-K-Ahx Induce Selective Cellular Death in Breast Cancer Cell Lines through Apoptotic Pathway

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
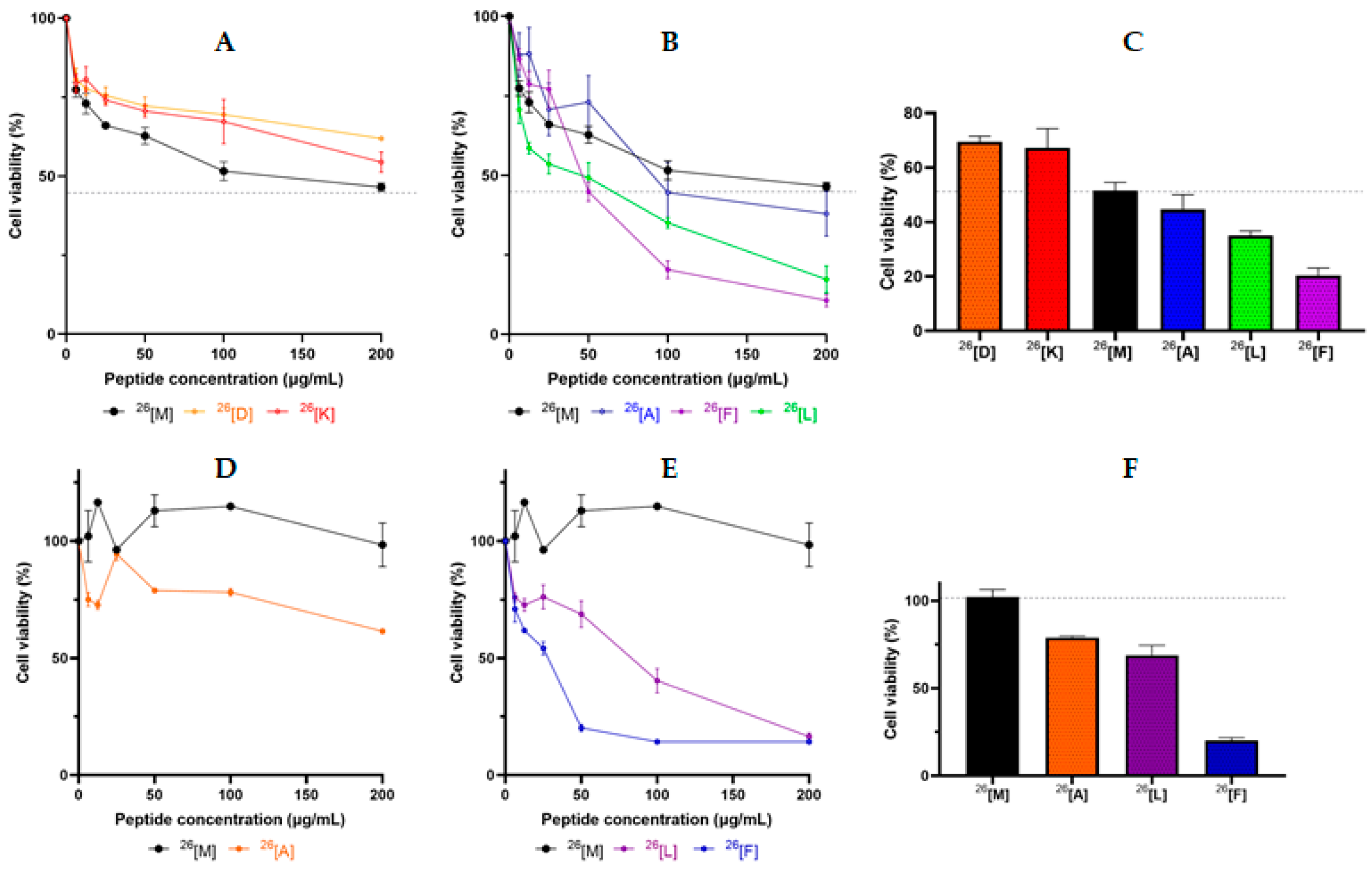
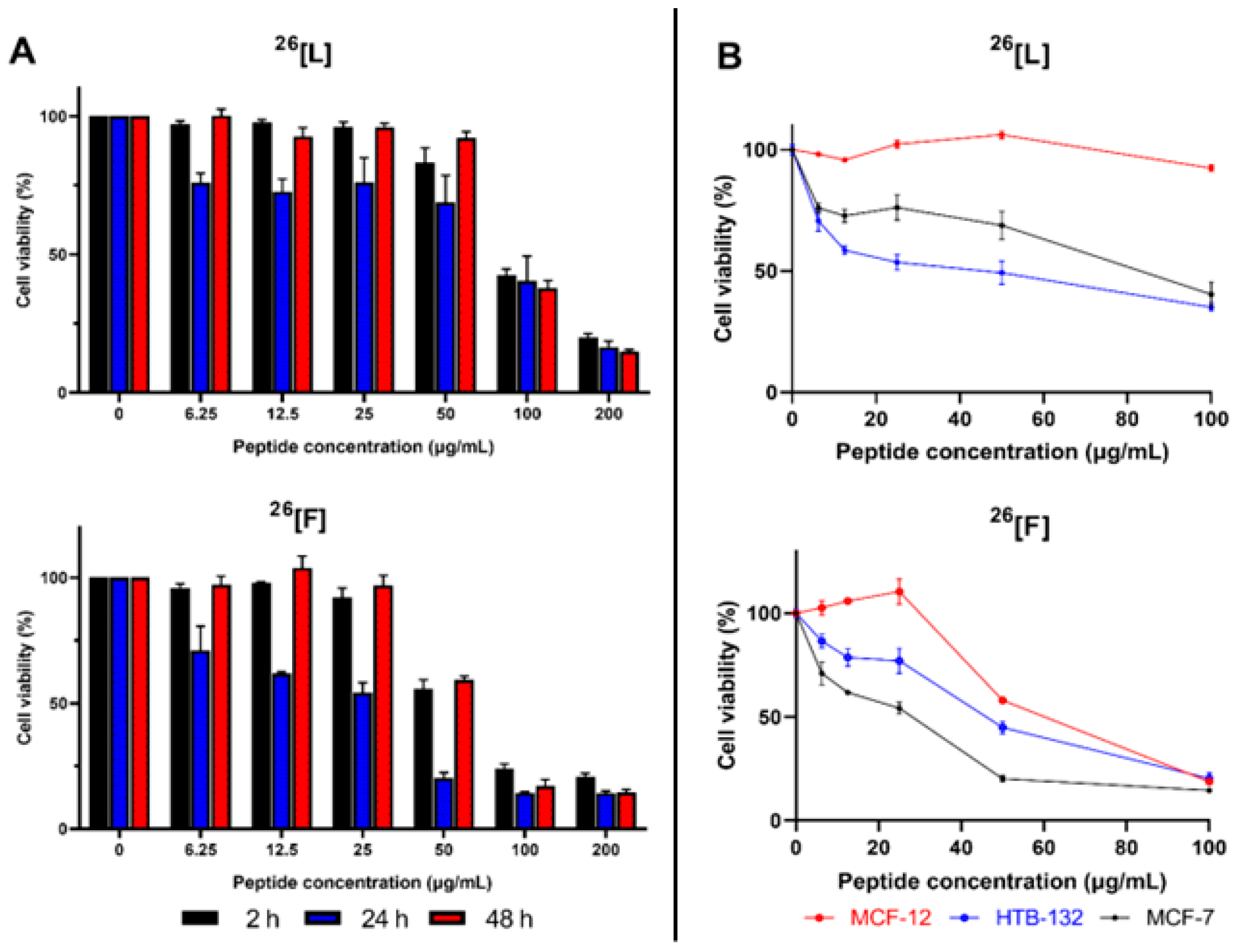
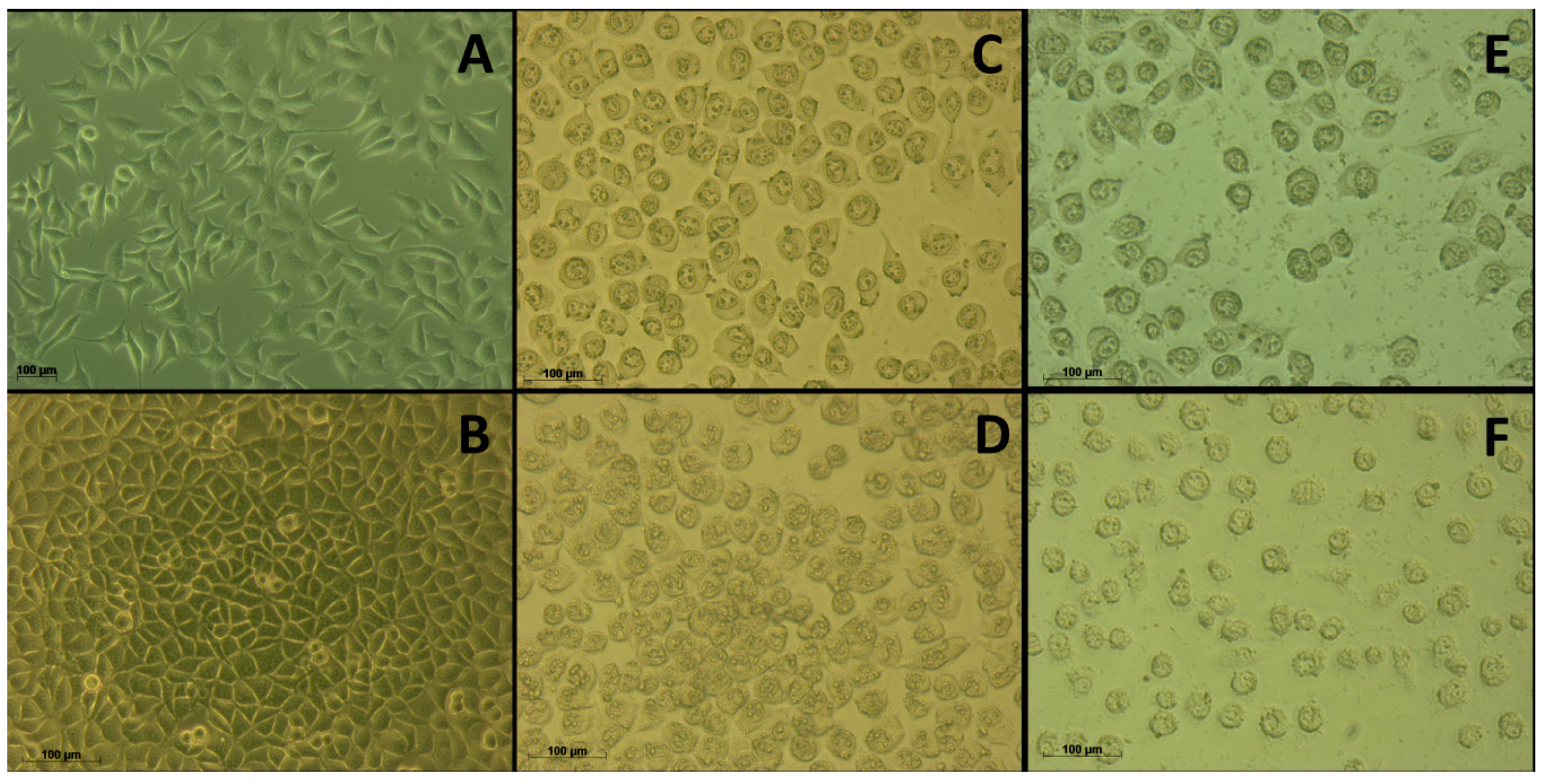
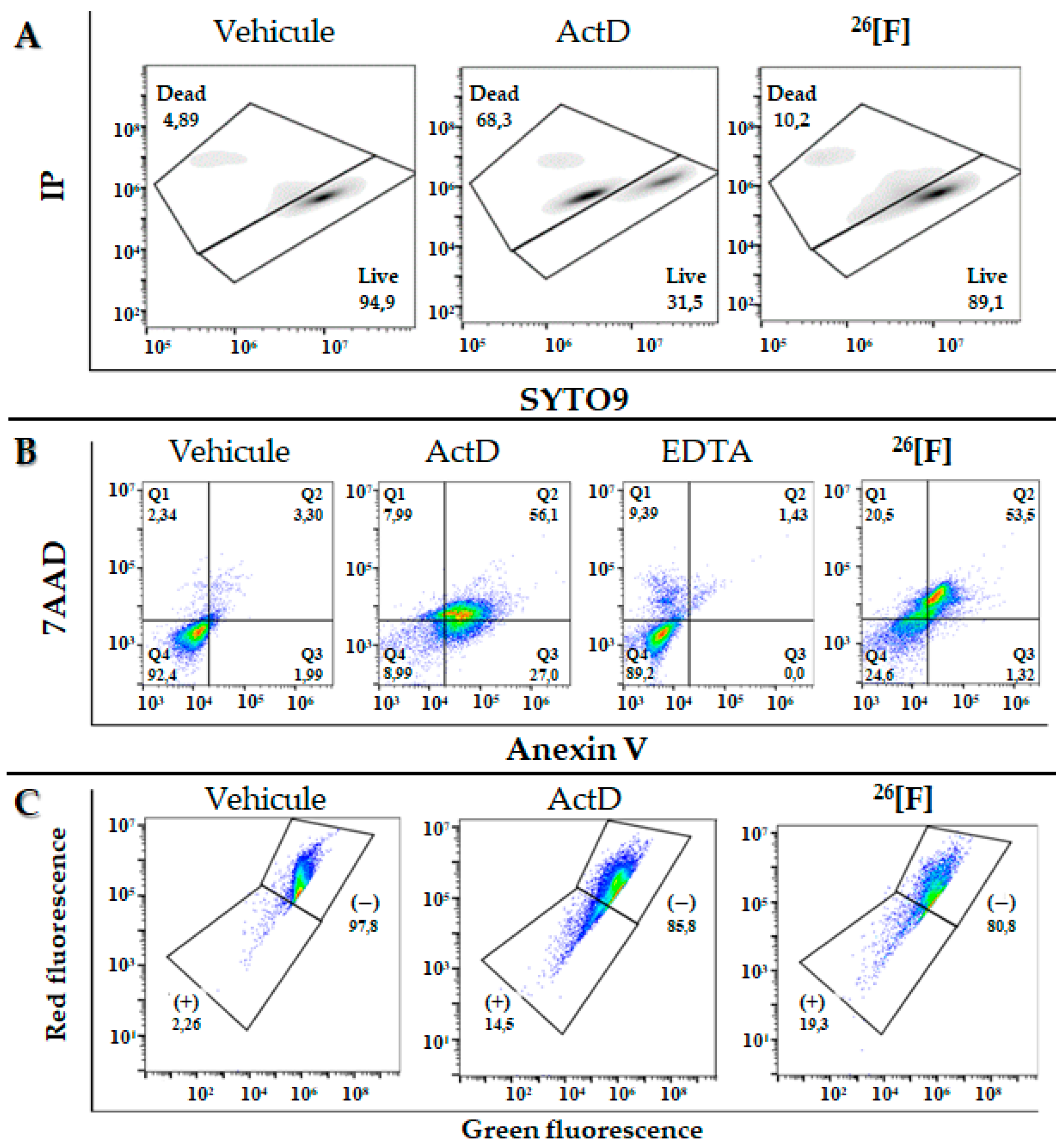
2. Results
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Solid Phase Synthesis SPPS
4.3. RP-HPLC Characterization
4.4. Peptide Purification by Solid Phase Extraction (SPE)
4.5. MALDI-TOF Mass Spectrometry Analysis
4.6. Cell Culture
4.7. Viability Test by MTT
4.8. Evaluation of the Integrity of the Cytoplasmic Membrane Using SYTO9/PI
4.9. Determination of the Type of cell Death (Apoptosis/Necrosis)
4.10. Determination of Mitochondrial Membrane Depolarization
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ahx | 6-aminohexanoic residue |
| LFB | Lactoferrin |
| LfcinB | Bovine Lactoferricin |
| MALDI | Matrix assisted Laser Desortion ionization |
| MS | Mass spectrometry |
| RP-HPLC | Reverse phase High Performance Liquid Chromatography |
| RP-SPE | Reverse Phase Solid phase extraction |
| SPPS | Solid Phase Peptide Synthesis |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Human Organization (WHO). Available online: http://www.who.int/cancer/en/ (accessed on 22 May 2018).
- Globocan. Available online: http://globocan.iarc.fr/Default.aspx (accessed on 5 May 2019).
- DeSantis, C.E.; Ma, J.; Goding Sauer, A.; Newman, L.A.; Jemal, A. Breast cancer statistics, 2017, racial disparity in mortality by state. CA Cancer J. Clin. 2017, 67, 439–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naksuriya, O.; Okonogi, S.; Schiffelers, R.M.; Hennink, W.E. Curcumin nanoformulations: A review of pharmaceutical properties and preclinical studies and clinical data related to cancer treatment. Biomaterials 2014, 35, 3365–3383. [Google Scholar] [CrossRef]
- Delaney, G.; Jacob, S.; Featherstone, C.; Barton, M. The role of radiotherapy in cancer treatment: Estimating optimal utilization from a review of evidence-based clinical guidelines. Cancer 2005, 104, 1129–1137. [Google Scholar] [CrossRef]
- Kerlikowske, K.; Zhu, W.; Hubbard, R.A.; Geller, B.; Dittus, K.; Braithwaite, D.; Wernli, K.J.; Miglioretti, D.L.; O’Meara, E.S. Outcomes of screening mammography by frequency, breast density, postmenopausal hormone therapy. JAMa Intern. Med. 2013, 173, 807–816. [Google Scholar] [CrossRef] [Green Version]
- Clough, K.B.; Kaufman, G.J.; Nos, C.; Buccimazza, I.; Sarfati, I.M. Improving breast cancer surgery: A classification and quadrant per quadrant atlas for oncoplastic surgery. Ann. Surg. Oncol. 2010, 17, 1375–1391. [Google Scholar] [CrossRef]
- Hassett, M.J.; O’Malley, A.J.; Pakes, J.R.; Newhouse, J.P.; Earle, C.C. Frequency and cost of chemotherapy-related serious adverse effects in a population sample of women with breast cancer. J. Natl. Cancer Inst. 2006, 98, 1108–1117. [Google Scholar] [CrossRef]
- Mader, J.S.; Richardson, A.; Salsman, J.; Top, D.; De Antueno, R.; Duncan, R.; Hoskin, D.W. Bovine lactoferricin causes apoptosis in Jurkat T-leukemia cells by sequential permeabilization of the cell membrane and targeting of mitochondria. Exp. Cell Res. 2007, 3. [Google Scholar] [CrossRef]
- Mader, J.S.; Salsman, J.; Conrad, D.M.; Hoskin, D.W. Bovine lactoferricin selectively induces apoptosis in human leukemia and carcinoma cell lines. Mol. Cancer 2005, 4, 612–624. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Tu, J.; Zhou, C.; Li, J.; Huang, L.; Tao, L.; Zhao, L. The effect of Lfcin-B on non-small cell lung cancer H460 cells is mediated by inhibiting VEGF expression and inducing apoptosis. Arch. Pharm. Res. 2015, 38, 261–271. [Google Scholar] [CrossRef]
- Alexander, D.B.; Vogel, H.J.; Tsuda, H. Lactoferrin researchers descend on Nagoya Castle1. Biochem. Cell Biol. 2017, 95, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Solarte, V.A.; Rosas, J.E.; Rivera, Z.J.; Arango-rodríguez, M.L.; García, J.E.; Vernot, J. A Tetrameric Peptide Derived from Bovine Lactoferricin Exhibits Specific Cytotoxic Effects against Oral Squamous-Cell Carcinoma Cell Lines. BioMed Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Vargas Casanova, Y.; Rodríguez Guerra, J.A.; Umaña Pérez, Y.A.; Leal Castro, A.L.; Almanzar Reina, G.; García Castañeda, J.E.; Rivera Monroy, Z.J. Antibacterial synthetic peptides derived from bovine lactoferricin exhibit cytotoxic effect against MDA-MB-468 and MDA-MB-231 breast cancer cell lines. Molecules 2017, 22, 1641. [Google Scholar] [CrossRef] [PubMed]
- Guerra, J.R.; Cárdenas, A.B.; Ochoa-Zarzosa, A.; Meza, J.L.; Umaña Pérez, A.; Fierro-Medina, R.; Rivera Monroy, Z.J.; García Castañeda, J.E. The tetrameric peptide LfcinB (20-25)4 derived from bovine lactoferricin induces apoptosis in the MCF-7 breast cancer cell line. RSC Adv. 2019, 9, 20497–20504. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, A.; Stroup, A.M.; Paddock, L.E.; Hill, S.; Plascak, J.; Llanos, A. Breast cancer incidence and mortality by molecular subtype: Statewide age and racial/ethnic disparities in new jerney. Cancer Health Disparities 2019, 3, e1–e17. [Google Scholar]
- Howlader, N.; Noone, A.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J. SEER cancer statistics review. Natl. Cancer Inst. 2020. [Google Scholar]
- Yao, L.; Zhang, Y.; Chen, K.; Hu, X.; Xu, L.X. Discovery of IL-18 as a novel secreted protein contributing to doxorubicin resistance by comparative secretome analysis of MCF-7 and MCF-7/Dox. PLoS ONE 2011, 6, e24684. [Google Scholar] [CrossRef]
- Cos, P.; Vlietinck, A.J.; Vanden, D.; Maes, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘ proof-of-concept. J. Ethnofarmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef]
- Hakimuddin, F.; Paliyath, G.; Meckling, K. Selective cytotoxicity of a red grape wine flavonoid fraction against MCF-7 cells. Breast Cancer Res. Treat. 2004, 85, 65–79. [Google Scholar] [CrossRef]
- Van Zijl, C.; Lottering, M.L.; Steffens, F.; Joubert, A. In vitro effects of 2-methoxyestradiol on MCF-12A and MCF-7 cell growth, morphology and mitotic spindle formation. Cell Biochem. Funct. 2008, 26, 632–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solarte, A.; Conget, P.; Vernot, J.; Rosas, J.E.; Arango-rodrı, M.L.; Rivera, J.; Garcı, J.E. A tetrameric peptide derived from bovine lactoferricin as a potential therapeutic tool for oral squamous cell carcinoma: A preclinical model. PLoS ONE 2017, 12, e0174707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiosses, W.B.; Hahn, K.M.; Giannelli, G.; Quaranta, V. Characterization of morphological and cytoskeletal changes in MCF10A breast epithelial cells plated on laminin-5: Comparison with breast cancer cell line MCF7. Cell Commun. Adhes. 2001, 8, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, X.; Gueydan, C.; Han, J. Plasma membrane changes during programmed cell deaths. Nat. Publ. Gr. 2017, 28, 9–21. [Google Scholar] [CrossRef]
- Lee, S.; Won, S.; Pyo, C.; Yoo, N.; Kim, J.; Choi, S. Requirement of the JNK-associated Bcl-2 pathway for human lactoferrin-induced apoptosis in the Jurkat leukemia T cell line. Biochimie 2009, 91, 102–108. [Google Scholar] [CrossRef]
- Pan, W.; Chen, P.; Chen, Y.S.; Hsu, H.; Lin, C.; Chen, W. Bovine lactoferricin B induces apoptosis of human gastric cancer cell line AGS by inhibition of autophagy at a late stage. J. Dairy Sci. 2013, 96, 7511–7520. [Google Scholar] [CrossRef]
- Roy, M.K.; Kuwabara, Y.; Hara, K.; Watanabe, Y.; Tamai, Y. Peptides From the N-terminal End of Bovine Lactoferrin Induce Apoptosis in Human Leukemic (HL-60) Cells. J. Dairy Sci. 2002, 85, 2065–2074. [Google Scholar] [CrossRef] [Green Version]
- Kanwar, R.K.; Kanwar, J.R. Immunomodulatory lactoferrin in the regulation of apoptosis modulatory proteins in cancer. Protein Pept. Lett. 2013, 20, 450–458. [Google Scholar]
- Furlong, S.J.; Mader, J.S.; Hoskin, D.W. Bovine lactoferricin induces caspase-independent apoptosis in human B-lymphoma cells and extends the survival of immune-de fi cient mice bearing B-lymphoma xenografts. Exp. Mol. Pathol. 2010, 88, 371–375. [Google Scholar] [CrossRef]
- Xu, X.X.; Jiang, H.R.; Li, H.B.; Zhang, T.N.; Zhou, Q.; Liu, N. Apoptosis of stomach cancer cell SGC-7901 and regulation of Akt signaling way induced by bovine lactoferrin. J. Dairy Sci. 2010, 93, 2344–2350. [Google Scholar] [CrossRef]
- Yoo, Y.; Watanabe, R.; Koike, Y.; Mitobe, M.; Shimazaki, K.; Watanabe, S.; Azuma, I. Apoptosis in human leukemic cells induced by lactoferricin, a bovine milk protein-derived peptide: Involvement of reactive oxygen species. Biochem. Biophys. Res. Commun. 1997, 637, 624–628. [Google Scholar] [CrossRef]
- Zhang, Y.; Lima, C.F.; Rodrigues, L.R. In vitro evaluation of bovine lactoferrin potential as an anticancer agent. Int. Dairy J. 2015, 40, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Zaldivar, M.A.; Andres, M.T.; Rego, A.; Pereira, C.S.; Fierro, J.F.; Real, M.C. Human lactoferrin triggers a mitochondrial- and caspase-dependent regulated cell death in Saccharomyces cerevisiae. Apoptosis. 2016, 21, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Amiri, F.; Moradian, F.; Rafiei, A. Research in molecular medicine anticancer effect of lactoferrin on gastric cancer cell line AGS. Res. Mol. Med. 2015, 3, 11–16. [Google Scholar]
- Richardson, A.; De Antueno, R.; Duncan, R.; Hoskin, D.W. Biochemical and Biophysical Research Communications Intracellular delivery of bovine lactoferricin’s antimicrobial core (RRWQWR) kills T-leukemia cells. Biochem. Biophys. Res. Commun. 2009, 388, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Hilchie, A.L.; Haney, E.F.; Bolscher, J.G.M.; Eric, M.H.; Hancock, R.E.W.; Vogel, H.J. Anticancer activities of bovine and human lactoferricin-derived peptides. Biochem. Cell Biol. 2017, 95, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, D.C.; Nicolau, A.; Teixeira, J.A.; Rodrigues, L.R. The effect of bovine milk lactoferrin on human breast cancer cell lines. J. Dairy Sci. 2011, 94, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, S.J.; Mader, J.S.; Hoskin, D.W. Lactoferricin-induced apoptosis in estrogen-nonresponsive MDA-MB-435 breast cancer cells is enhanced by C 6 ceramide or tamoxifen. Oncol. Rep. 2006, 15, 1385–1390. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.A.; Kanwar, J.R.; Kanwar, R.K. Iron-free and iron-saturated bovine lactoferrin inhibit survivin expression and differentially modulate apoptosis in breast cancer. BMC Cancer 2015, 15, 425. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Lin, Y.; Xiao, Y.; Crommen, J.; Jiang, Z. Recent developments in cyclodextrin functionalized monolithic columns for the enantioseparation of chiral drugs. J. Pharm. Biomed. Anal. 2016, 130, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and arginine-rich antimicrobial peptides: Structures and mechanisms of action. Biochim. Biophys. Acta 2006, 1758, 1184–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umeyama, M.; Kira, A.; Nishimura, K.; Naito, A. Interactions of bovine lactoferricin with acidic phospholipid bilayers and its antimicrobial activity as studied by solid-state NMR. Biochim. Biophys. Acta 2006, 1758, 1523–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Carmela, M.; Valenti, P.; Musci, G. Lactoferrin’s anti-cancer properties: Safety, selectivity, and wide range of action. Biomolecules 2020, 10, 456. [Google Scholar] [CrossRef] [Green Version]
- Sadiq, I.Z.; Babagana, K.; Danlami, D. Peptides: A closer look at bovine lactoferricin. Mol. Cancer 2018, 1, 1–9. [Google Scholar] [CrossRef]
- Islam, Z.; Sharmin, S.; Dohra, H.; Yamazaki, M. Entry of 6-residue antimicrobial peptide derived from lactoferricin B into single vesicles and E. coli cells without damaging their membranes. Biochemistry 2017, 56, 4419–4431. [Google Scholar] [CrossRef]
- Vergel Galeano, C.F.; Rivera Monroy, Z.J.; Rosas Pérez, J.E.; García Castañeda, J.E. Efficient synthesis of peptides with 4-methylpiperidine as Fmoc removal reagent by solid phase synthesis. J. Mex. Chem. Soc. 2014, 58, 386–392. [Google Scholar]
- de Jesús Huertas, N.; Rivera Monroy, Z.J.; Fierro Medina, R.; García Castañeda, J.E. Antimicrobial Activity of Truncated and Polyvalent Peptides Derived from the FKCRRQWQWRMKKGLA Sequence against Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 25923. Molecules 2017, 22, 987. [Google Scholar] [CrossRef] [Green Version]
- Insuasty-Cepeda, D.S.; Rodríguez-Mayor, A.V.; Pineda-Castañeda, H.M.; García-Castañeda, J.E.; Maldonado-Villamil, M.; Fierro-Medina, R.; Rivera-Monroy, Z.J. Synthetic peptide purification via solid-phase economical, fast, and efficient methodology. Molecules 2019, 24, 1215. [Google Scholar] [CrossRef] [Green Version]
- Langan, T.; Rodgers, K.; Chou, R. Synchromization of mammalian cell cultures by serum deprivation. Methods Cell Sci. 2017, 1524, 97–105. [Google Scholar] [CrossRef]
- Robertson, T.; McGoverin, C.; Vanholsbeek, D.; Swift, S. Optimisation of the Protocol for the LIVE/DEAD® BacLightTM Bacterial Viability Kit for Rapid Determination of Bacterial Load. Front. Microbiol. 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | ||||||
|---|---|---|---|---|---|---|
| Code | Sequence | Characterization | Cytotoxic Effect | |||
| RP-HPLC | m/z [M+H]+ | IC50 (µM)/(µg/mL) | ||||
| tR (min) | Purity a (%) | HTB-132 | MCF-7 | |||
| 26[M] | (RRWQWRMKKLG)2-K-Ahx | 5.3 | 94 | 3312.3 | 30/96 | >60/>200 |
| 26[K] | (RRWQWRKKKLG)2-K-Ahx | 4.8 | 95 | 3302.7 | >60/>200 | N/D |
| 26[D] | (RRWQWRDKKLG)2-K-Ahx | 4.8 | 97 | 3276.7 | >60/>200 | N/D |
| 26[A] | (RRWQWRAKKLG)2-K-Ahx | 4.8 | 95 | 3187.9 | 26/96 | >60/>200 |
| 26[F] | (RRWQWRFKKLG)2-K-Ahx | 5.6 | 91 | 3342.5 | 13/43 | 6/19 |
| 26[L] | (RRWQWRLKKLG)2-K-Ahx | 5.5 | 92 | 3272.4 | 10/32 | 20/66 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Insuasty-Cepeda, D.S.; Barragán-Cárdenas, A.C.; Ochoa-Zarzosa, A.; López-Meza, J.E.; Fierro-Medina, R.; García-Castañeda, J.E.; Rivera-Monroy, Z.J. Peptides Derived from (RRWQWRMKKLG)2-K-Ahx Induce Selective Cellular Death in Breast Cancer Cell Lines through Apoptotic Pathway. Int. J. Mol. Sci. 2020, 21, 4550. https://doi.org/10.3390/ijms21124550
Insuasty-Cepeda DS, Barragán-Cárdenas AC, Ochoa-Zarzosa A, López-Meza JE, Fierro-Medina R, García-Castañeda JE, Rivera-Monroy ZJ. Peptides Derived from (RRWQWRMKKLG)2-K-Ahx Induce Selective Cellular Death in Breast Cancer Cell Lines through Apoptotic Pathway. International Journal of Molecular Sciences. 2020; 21(12):4550. https://doi.org/10.3390/ijms21124550
Chicago/Turabian StyleInsuasty-Cepeda, Diego Sebastián, Andrea Carolina Barragán-Cárdenas, Alejandra Ochoa-Zarzosa, Joel E. López-Meza, Ricardo Fierro-Medina, Javier Eduardo García-Castañeda, and Zuly Jenny Rivera-Monroy. 2020. "Peptides Derived from (RRWQWRMKKLG)2-K-Ahx Induce Selective Cellular Death in Breast Cancer Cell Lines through Apoptotic Pathway" International Journal of Molecular Sciences 21, no. 12: 4550. https://doi.org/10.3390/ijms21124550