Ultrastructural and Photosynthetic Responses of Pod Walls in Alfalfa to Drought Stress

Abstract
:1. Introduction
2. Results
2.1. Changes of the Surface Characteristics and Ultrastructure of Pod Wall under Drought Stress
2.2. Changes of Chlorophyll Concentration in Pod Wall under Drought Stress
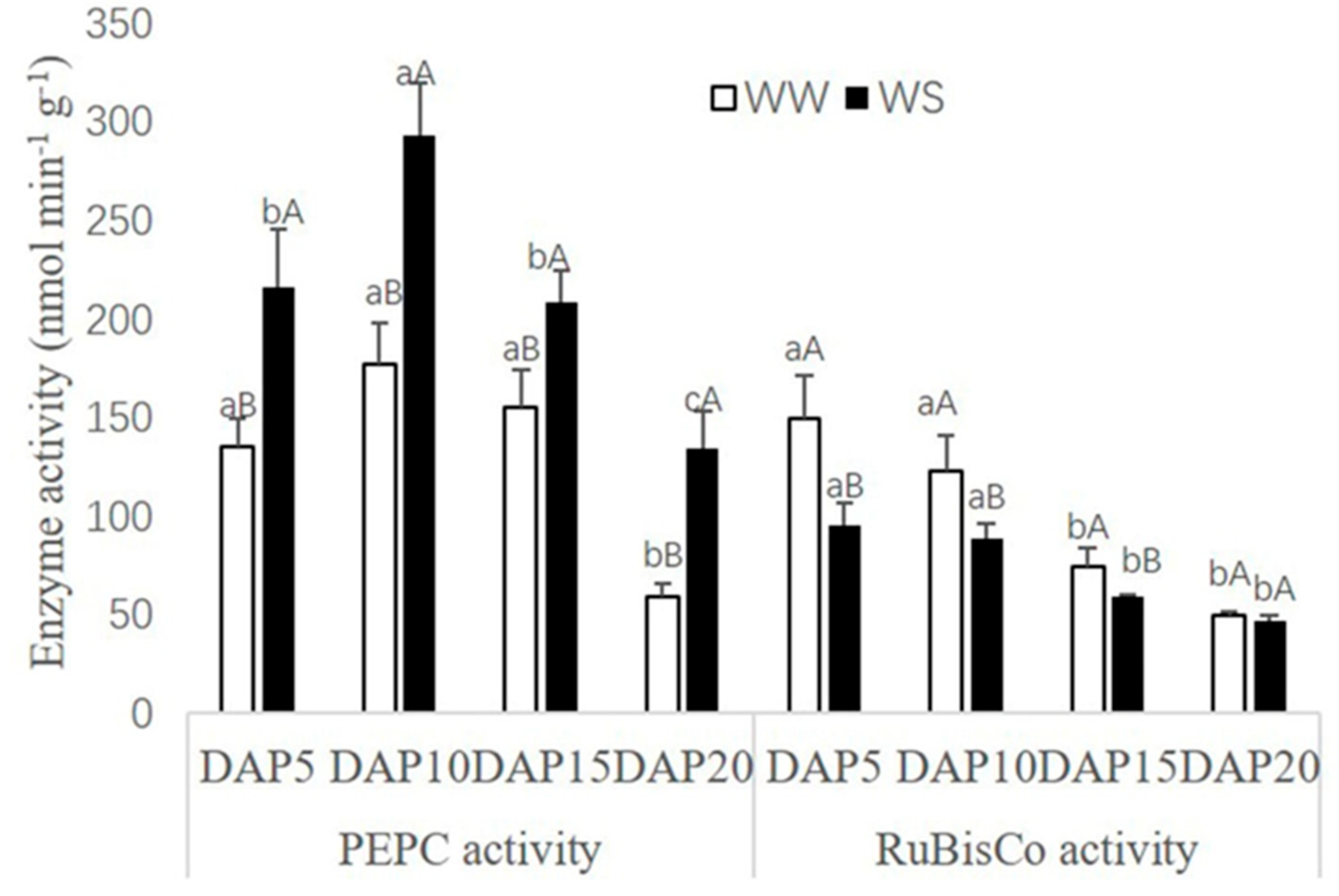
2.3. Changes of Photosynthetic Enzyme Activities in Pod Wall under Drought Stress
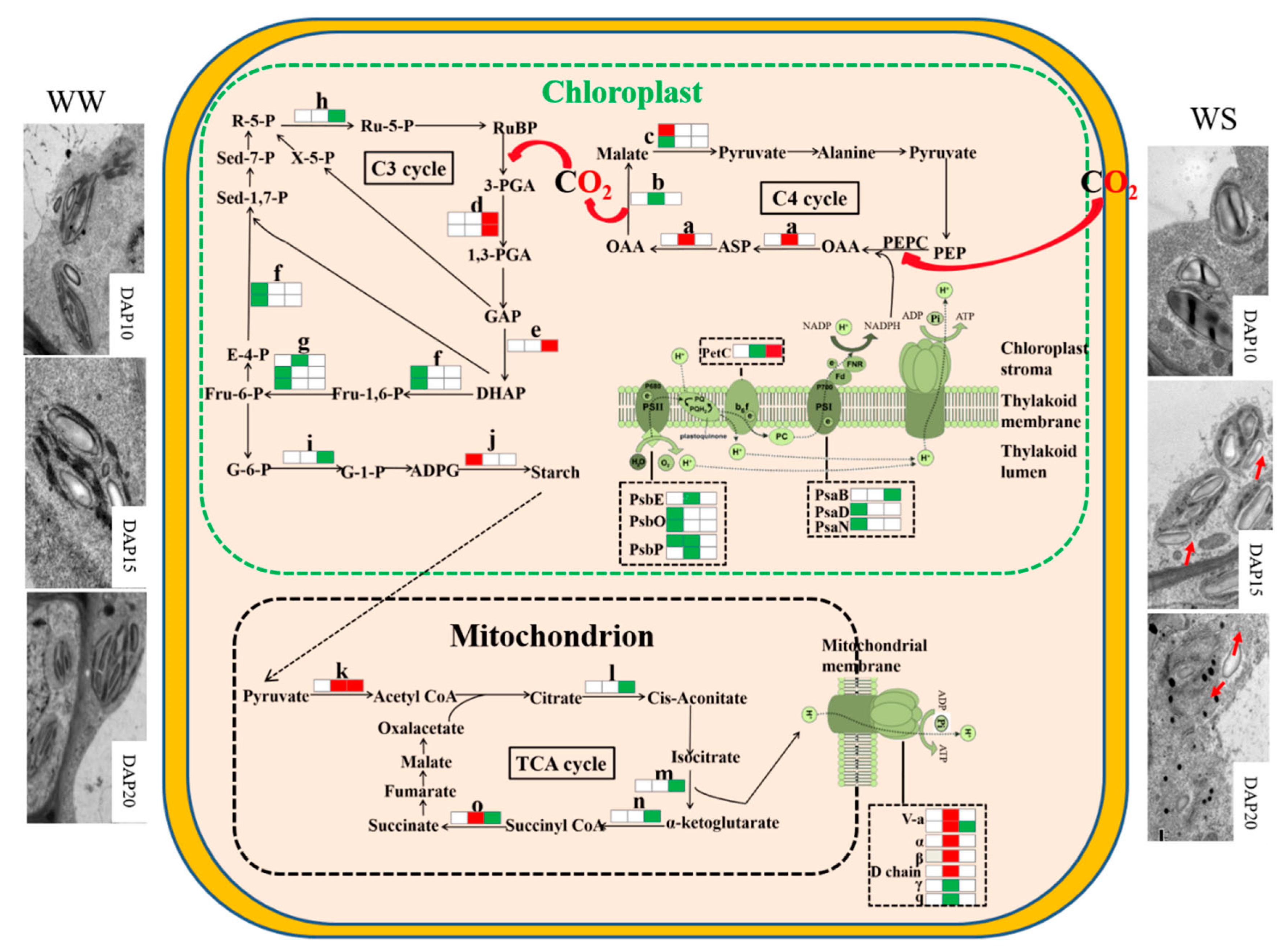
2.4. Proteomic Analysis on the Response of Pod Wall to Drought Stress
3. Discussion
3.1. Observation of Surface and Ultrastructure in the Pod Wall
3.2. Response of Chlorophyll Concentration and Photosynthetic Enzyme Activities
3.3. The Differential Expression of Proteins under Drought Stress
3.3.1. Photosynthesis-Antenna Proteins
3.3.2. Photosynthesis
3.3.3. Carbon Fixation in Photosynthetic Organisms
3.3.4. Carbohydrate Metabolism
3.3.5. Energy Metabolism
3.3.6. Other Metabolism
4. Material and Method
4.1. Material
4.2. Surface and Ultrastructure Characteristics Observation for Pod Wall
4.3. Chlorophyll Concentration of Pod Walls Measurement
4.4. Photosynthetic Enzyme Activities Assays
4.5. Proteomic Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Loomis, R.S.; Williams, W.A. Maximum crop productivity: An extimate. Crop Sci. 1963, 3, 67–72. [Google Scholar] [CrossRef]
- Shen, Y. The Most Improtant Chemistry Reaction on the Earth-Photosynthesis; Tsinghua University Press: Beijing, China, 2000; pp. 2–13. [Google Scholar]
- Long, S.; Marshall-Colon, A.; Zhu, X. Meeting the global food demand of the future by engineering crop photosynthesis and yield potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yang, J.; Hai, J.; Chen, W.; Zhao, X. Photosynthetic features of leaf and silique of ‘Qinyou 7′oilseed rape (Brassica napus L.) at reproductive growth stage. Oil Crop. Sci. 2018, 3, 176. [Google Scholar]
- Zhang, Y.; Mulpuri, S.; Liu, A. Photosynthetic capacity of the capsule wall and its contribution to carbon fixation and seed yield in castor (Ricinus communis L.). Acta Physiol. Plant. 2016, 38, 245. [Google Scholar] [CrossRef]
- Furbank, R.T.; White, R.; Palta, J.A.; Turner, N.C. Internal recycling of respiratory CO2 in pods of chickpea (Cicer arietinum L.): The role of pod wall, seed coat, and embryo. J. Exp. Bot. 2004, 55, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Kariyat, D.; Marriboina, S.; Reddy, A.R. Pod-wall proteomics provide novel insights into soybean seed-filling process under chemical-induced terminal drought stress. J. Sci. Food Agric. 2019, 99, 2481–2493. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hou, L.; Wang, M.; Mao, P. Contribution of the pod wall to seed grain filling in alfalfa. Sci. Rep. 2016, 6, 26586. [Google Scholar] [CrossRef] [Green Version]
- Tambussi, E.A.; Bort, J.; Guiamet, J.J.; Nogués, S.; Araus, J.L. The photosynthetic role of ears in C3 cereals: Metabolism, water use efficiency and contribution to grain yield. Crit. Rev. Plant Sci. 2007, 26, 1–16. [Google Scholar] [CrossRef]
- Jiang, Q.; Roche, D.; Durham, S.; Hole, D. Awn contribution to gas exchanges of barley ears. Photosynthetica 2006, 44, 536–541. [Google Scholar] [CrossRef]
- Baluar, N.; Badicean, D.; Peterhaensel, C.; Mereniuc, L.; Vorontsov, V.; Terteac, D. The peculiarities of carbon metabolism in the ears of C3 cereals CO2 exchange kinetics, chloroplasts structure and ultra-structure in the cells from photosynthetic active components of the ear. J. Tissue Cult. Bioeng. 2018, 1, 1–14. [Google Scholar]
- AuBuchon-Elder, T.; Coneva, V.; Goad, D.M.; Allen, D.K.; Kellogg, E.A. Sterile spikelets assimilate carbon in sorghum and related grasses. BioRxiv 2018. [Google Scholar] [CrossRef]
- Ávila-Lovera, E.; Zerpa, A.J.; Santiago, L.S. Stem photosynthesis and hydraulics are coordinated in desert plant species. New Phytol. 2017, 216, 1119–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaya, Y.; Yabuki, K.; Kiyota, M.; Tani, A.; Hirano, T.; Aiga, I. Gas exchange and oxygen concentration in pneumatophores and prop roots of four mangrove species. Trees-Struct. Funct. 2002, 16, 155–158. [Google Scholar] [CrossRef]
- Zhang, C.; Zhan, D.; Luo, H.; Zhang, Y.; Zhang, W. Photorespiration and photoinhibition in the bracts of cotton under water stress. Photosynthetica 2016, 54, 12–18. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Abebe, T.; Melmaiee, K.; Berg, V.; Wise, R.P. Drought response in the spikes of barley: Gene expression in the lemma, palea, awn, and seed. Funct. Interg. Genomic. 2010, 10, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Wu, Y.; Zhang, X. Stomatal characteristics of different green organs in wheat under different irrigation regimes. Acta Agron. Sin. 2006, 32, 70–75. [Google Scholar]
- Hu, Y.; Zhang, Y.; Yi, X.; Zhan, D.; Luo, H.; Soon, C.W.; Zhang, W. The relative contribution of non-foliar organs of cotton to yield and related physiological characteristics under water deficit. J. Integr. Agric. 2014, 13, 975–989. [Google Scholar] [CrossRef]
- Lou, L.; Li, X.; Chen, J.; Li, Y.; Tang, Y.; Lv, J. Photosynthetic and ascorbate-glutathione metabolism in the flag leaves as compared to spikes under drought stress of winter wheat (Triticum aestivum L.). PLoS ONE 2018, 13, e0194625. [Google Scholar] [CrossRef] [Green Version]
- Wei, A.; Wang, Z.; Zhai, Z.; Cong, Y. Effect of soil drought on C4 photosynthetic enzyme activities of flag leaf and ear in wheat. Agric. Sci. China 2003, 36, 508–512. [Google Scholar]
- Kong, L.; Wang, F.; Feng, B.; Li, S.; Si, J.; Zhang, B. The structural and photosynthetic characteristics of the exposed peduncle of wheat (Triticum aestivum L.): An important photosynthate source for grain-filling. BMC Plant Biol. 2010, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Lv, J.; Jiang, S.; Liang, T.; Liu, C.; Jing, Z. Response of wheat ear photosynthesis and photosynthate carbon distribution to water deficit. Photosynthetica 2015, 53, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Rangan, P.; Furtado, A.; Henry, R.J. New evidence for grain specific C4 photosynthesis in wheat. Sci. Rep. 2016, 6, 31721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singal, H.R.; Sheoran, I.S.; Singh, R. In vitro enzyme activities and products of 14CO2 assimilation in flag leaf and ear parts of wheat (Triticum aestivum L.). Photosynth. Res. 1986, 8, 113–122. [Google Scholar] [CrossRef]
- Gu, S.; Yin, L.; Wang, Q. Phosphoenolpyruvate carboxylase in the stem of the submersed species Egeria densa may be involved in an inducible C 4-like mechanism. Aquat. Bot. 2015, 125, 1–8. [Google Scholar] [CrossRef]
- Atkins, C.A.; Kuo, J.; Pate, J.S.; Flinn, A.M.; Steele, T.W. Photosynthetic pod wall of pea (Pisum sativum L.) distribution of carbon dioxide-fixing enzymes in relation to pod structure. Plant Physiol. 1977, 60, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhang, Y.; Luo, H.; Li, W.; Oguchi, R.; Fan, D.; Soon, C.W.; Zhang, W. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta 2012, 235, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Xu, Y.; Jiang, S.; Lu, K.; Lu, Y.; Feng, X.; Zhen, W.; Shan, L.; Yu, Y.; Wang, X.; et al. Light-harvesting chlorophyll a/b-binding proteins, positively involved in abscisic acid signalling, require a transcription repressor, WRKY40, to balance their function. J. Exp. Bot. 2013, 64, 5443–5456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, P. Physiological and Molecular Basis of Drought Resistance Enhanced by Si Application in Sugarcane. Ph.D. Thesis, Guangxi University, Nanning, China, 2012. [Google Scholar]
- Cui, Q. Analysis of the Chloroplast Proteome of Cucumber Leaves under Elevated CO2 Concentration and Drought Stress. Masters’ Thesis, Shandong Agricultural University, Taian, China, 2017. [Google Scholar]
- Hossain, M.A.; Wani, S.H.; Bhattacharjee, S.; Burritt, D.J.; Tran, L.S.P. Drought Stress Tolerance in Plants, Physiology and Biochemistry; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, pp. 155–157. [Google Scholar]
- Ifuku, K. Localization and functional characterization of the extrinsic subunits of photosystem II: An update. Biosci. Biotechnol. Biochem. 2015, 79, 1223–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, G.; Cavaliere, C.; Foglia, P.; Gubbiotti, R.; Samperi, R.; Laganà, A. Analysis of drought responsive proteins in wheat (Triticum durum) by 2D-PAGE and MALDI-TOF mass spectrometry. Plant Sci. 2009, 177, 570–576. [Google Scholar] [CrossRef]
- Xiao, X.; Yang, F.; Zhang, S.; Korpelainen, H.; Li, C. Physiological and proteomic responses of two contrasting Populus cathayana populations to drought stress. Physiol. Plant. 2009, 136, 150–168. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Sun, X.; Lin, H.; Chen, J.; Ren, J.; Hu, X.; Yang, Y. Comparative physiological and proteomic analyses of poplar (Populus yunnanensis) plantlets exposed to high temperature and drought. PLoS ONE 2014, 9, e107605. [Google Scholar] [CrossRef]
- Das, A.; Eldakak, M.; Paudel, B.; Kim, D.W.; Hemmati, H.; Basu, C.; Rohila, J.S. Leaf proteome analysis reveals prospective drought and heat stress response mechanisms in soybean. BioMed Res. Int. 2016, 6021047. [Google Scholar] [CrossRef] [Green Version]
- Aranjuelo, I.; Molero, G.; Erice, G.; Avice, J.C.; Nogués, S. Plant physiology and proteomics reveals the leaf response to drought in alfalfa (Medicago sativa L.). J. Exp. Bot. 2010, 62, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Y.; Wang, Z. Photosynthetic diurnal variation characteristics of leaf and non-leaf organs in winter wheat under different irrigation regimes. Acta Ecol. Sin. 2011, 31, 1312–1322. [Google Scholar]
- Feng, B. Physiological Basis and Response Mechanisms of Different Green Organs under High Temperature after Anthesis in Winter Wheat. Ph.D. Thesis, Shandong Agricultural University, Taian, China, 2014. [Google Scholar]
- Pei, C.; Zhang, Z.; Ma, J. Differentially expressed proteins analysis of seedling leaf of southern type alfalfa (Medicago sativa ‘Millenium’) under salt stress. J. Agric. Biotechnol. 2016, 24, 1629–1642. [Google Scholar]
- Li, H. Phosphoproteomic Differentiation of Maize Seedlings in Response to Drought Stress and Abscisic and Induction. Masters’ Thesis, Sichuan Agricultural University, Chengdu, China, 2016. [Google Scholar]
- Umeda, M.; Hara, C.; Matsubayashi, Y.; Li, H.; Liu, Q.; Tadokoro, F.; Aotsuka, S.; Uchimiya, H. Expressed sequence tags from cultured cells of rice (Oryza sativa L.) under stressed conditions: Analysis of transcripts of genes engaged in ATP-generating pathways. Plant Mol. Biol. 1994, 25, 469–478. [Google Scholar] [CrossRef]
- Riccardi, F.; Gazeau, P.; de Vienne, D.; Zivy, M. Protein changes in response to progressive water deficit in maize: Quantitative variation and polypeptide identification. Plant Physiol. 1998, 117, 1253–1263. [Google Scholar] [CrossRef] [Green Version]
- Silveira, R.D.D.; Abreu, F.R.M.; Mamidi, S.; McClean, P.E.; Vianello, R.P.; Lanna, A.C.; Carneiro, N.P.; Brondani, C. Expression of drought tolerance genes in tropical upland rice cultivars (Oryza sativa). Genet. Mol. Res. 2015, 14, 8181–8200. [Google Scholar] [CrossRef]
- Phan, T.T.; Li, J.; Sun, B.; Jia, Y.; Wen, H.; Chan, H.; Li, T.; Li, Y. ATP-citrate lyase gene (SOACLA-1), a novel ACLA gene in sugarcane, and its overexpression enhance drought tolerance of transgenic tobacco. Sugar Technol. 2017, 19, 258–269. [Google Scholar] [CrossRef]
- Stitt, M.; Gibon, Y.; Lunn, J.E.; Piques, M. Multilevel genomics analysis of carbon signalling during low carbon availability: Coordinating the supply and utilisation of carbon in a fluctuating environment. Funct. Plant Biol. 2007, 34, 526–549. [Google Scholar] [CrossRef]
- Budak, H.; Akpinar, B.A.; Unver, T.; Turktas, M. Proteome changes in wild and modern wheat leaves upon drought stress by two-dimensional electrophoresis and nanoLC-ESI–MS/MS. Plant Mol. Biol. 2013, 83, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-responsive mechanisms in plant leaves revealed by proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregorova, Z.; Kovacik, J.; Klejdus, B.; Maglovski, M.; Kuna, R.; Hauptvogel, P.; Matušíková, I. Drought-induced responses of physiology, metabolites, and PR proteins in Triticum aestivum. J. Agric. Food Chem. 2015, 63, 8125–8133. [Google Scholar] [CrossRef]
- Li, P.; Zhang, Y.; Wu, X.; Liu, Y. Drought stress impact on leaf proteome variations of faba bean (Vicia faba L.) in the Qinghai–Tibet Plateau of China. 3 Biotech 2018, 8, 110. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Li, Z.; Chen, Y.; Chen, M.; Li, L.; Ma, Y. Heat shock protein 90 in plants: Molecular mechanisms and roles in stress responses. Int. J. Mol. Sci. 2012, 13, 15706–15723. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Seki, M.; Mochizuki, Y.; Heida, N.; Hirosawa, K.; Okamoto, N.; Sakurai, T.; Satou, M.; Akiyama, K.; Iida, K.; et al. A flexible representation of omic knowledge for thorough analysis of microarray data. Plant Methods 2006, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Ashoub, A.; Beckhaus, T.; Berberich, T.; Karas, M.; Brüggemann, W. Comparative analysis of barley leaf proteome as affected by drought stress. Planta 2013, 237, 771–781. [Google Scholar] [CrossRef]
- Lim, C.W.; Han, S.W.; Hwang, I.S.; Kim, D.S.; Hwang, B.K.; Lee, S.C. The pepper lipoxygenase CaLOX1 plays a role in osmotic, drought and high salinity stress response. Plant Cell Physiol. 2015, 56, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Kang, L. Biological Electron Microscopy Techniques; China Science and Technology University Press: Hefei, China, 2004. [Google Scholar]
- Blanke, M.M.; Ebert, G. Phosphoenolpyruvate carboxylase and carbon economy of apple seedlings. J. Exp. Bot. 1992, 43, 965–968. [Google Scholar] [CrossRef]
- Lilley, R.M.C.; Walker, D.A. An improved spectrophotometric assay for ribulosebisphosphate carboxylase. Biochim. Biophys. Acta 1974, 358, 226–229. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days after Pollination | Chlorophyll a (mg g−1) | Chlorophyll b (mg g−1) | Total Chlorophyll (mg g−1) | |||
|---|---|---|---|---|---|---|
| WW | WS | WW | WS | WW | WS | |
| DAP5 | 0.314 aA | 0.141 aB | 0.137 aA | 0.055 aB | 0.451 aA | 0.196 aB |
| DAP10 | 0.138 bA | 0.085 abB | 0.064 bA | 0.044 abA | 0.202 bA | 0.128 bB |
| DAP15 | 0.094 bA | 0.041 bcB | 0.055 bA | 0.025 bcB | 0.149 bA | 0.066 cB |
| DAP20 | 0.024 cA | 0.012 cB | 0.019 cA | 0.010 cB | 0.043 cA | 0.022 dB |
| KEGG | Accession | Proteins | Fold | ||
|---|---|---|---|---|---|
| WS10 vs. WW10 | WS15 vs. WW15 | WS20 vs. WW20 | |||
| Amino sugar and nucleotide sugar metabolism | A0A072UKS2 | PfkB family carbohydrate kinase | NS | NS | 0.4 |
| A0A072VQZ5 | UDP-D-apiose/UDP-D-xylose synthase | NS | NS | 0.2 | |
| G7JUS9 | UDP-glucuronic acid decarboxylase | NS | 3.3 | 0.4 | |
| G7ID31 | Chitinase | NS | NS | 5.0 | |
| G7LA76 | Chitinase (Class Ib)/Hevein | NS | NS | 3.3 | |
| Ascorbate and aldarate metabolism | A0A072TLF4 | Myo-inositol oxygenase | 0.4 | NS | NS |
| A0A072U2G7 | NAD-dependent aldehyde dehydrogenase family protein | 0.2 | 3.3 | NS | |
| A0A072UQP6 | UDP-glucose 6-dehydrogenase | 2.5 | NS | NS | |
| G7L571 | UDP-glucose 6-dehydrogenase | NS | 2.5 | NS | |
| A0A072V120 | UTP-glucose-1-phosphate uridylyltransferase | NS | 2.5 | NS | |
| A0A072V151 | L-ascorbate oxidase | 5.0 | NS | NS | |
| A0A072VNM9 | GME GDP-D-mannose-3, 5-epimerase | NS | 3.3 | 0.1 | |
| G7L1 × 0 | GME GDP-D-mannose-3, 5-epimerase | 2.5 | NS | 0.3 | |
| G7JTZ5 | Aldo/keto reductase family oxidoreductase | 0.5 | NS | NS | |
| G7KAG7 | Thylakoid lumenal 29 kDa protein | 0.3 | NS | NS | |
| beta-Alanine metabolism | A0A072UCM6 | Glutamate decarboxylase | NS | NS | 0.2 |
| Carbon metabolism | G7IT85 | Phosphoglycerate kinase | NS | NS | 3.3 |
| G7IT86 | Phosphoglycerate kinase | NS | NS | 3.3 | |
| G7KJZ8 | Glucose-6-phosphate isomerase | NS | NS | 0.4 | |
| G7L1U4 | Ribose-5-phosphate isomerase A | NS | NS | 0.4 | |
| I3S3S0 | Cytosolic triosephosphate isomerase | NS | NS | 2.0 | |
| A0A072VS77 | Methylenetetrahydrofolate reductase | NS | 3.3 | 0.3 | |
| Starch and sucrose metabolism | A0A072UCM8 | Phosphotransferase | 0.3 | NS | NS |
| A0A072UKS2 | PfkB family carbohydrate kinase | NS | NS | 0.4 | |
| A0A072UU47 | Glycoside hydrolase family 1 protein | 3.3 | NS | NS | |
| A0A072VLQ9 | Starch synthase | 2.5 | NS | NS | |
| G7IJV7 | Glycoside hydrolase family 3 protein | 0.3 | NS | NS | |
| G7KJZ8 | Glucose-6-phosphate isomerase | NS | NS | 0.4 | |
| Citrate cycle (TCA cycle) | G7KVS0 | E1 subunit-like 2-oxoglutarate dehydrogenase | NS | NS | 0.3 |
| G7JYQ8 | Aconitate hydratase | NS | NS | 0.3 | |
| B7FJJ4 | Pyruvate dehydrogenase E1 beta subunit | NS | 2.0 | 2.5 | |
| G7KHI5 | Isocitrate dehydrogenase [NADP] | NS | NS | 0.2 | |
| A2Q2V1 | ATP-citrate lyase/succinyl-CoA ligase | NS | 2.5 | 0.1 | |
| Glycine, serine and threonine metabolism | A0A072URB1 | Amine oxidase | NS | NS | 5.0 |
| A0A072V290 | Amine oxidase | NS | NS | 3.3 | |
| G7J7B0 | Amine oxidase | NS | NS | 5.0 | |
| A9YWS0 | Serine hydroxymethyltransferase | NS | NS | 0.2 | |
| G7I9Z0 | Glycine dehydrogenase [decarboxylating] protein | NS | NS | 0.4 | |
| G7JJ96 | Aminomethyltransferase | NS | NS | 0.3 | |
| G7JNS2 | NAD-dependent aldehyde dehydrogenase family protein | NS | NS | 0.4 | |
| G7L9H1 | Phosphoserine aminotransferase | NS | NS | 0.3 | |
| Linoleic acid metabolism | A0A072UMH4 | Lipoxygenase | 0.5 | NS | NS |
| G7J629 | Lipoxygenase | 0.4 | NS | NS | |
| G7LIX7 | Lipoxygenase | 5.0 | NS | NS | |
| G7LIY0 | Lipoxygenase | 5.0 | NS | 0.4 | |
| G7LIY2 | Lipoxygenase | 10.0 | NS | NS | |
| G7J632 | Lipoxygenase | 2.5 | 2.5 | NS | |
| Oxidative phosphorylation | A0A072URM9 | Archaeal/vacuolar-type H+-ATPase subunit A | NS | 2.0 | NS |
| A0A072V4G2 | F0F1 ATP synthase subunit gamma | NS | 0.5 | NS | |
| A0A072W1H5 | ATP synthase subunit beta | NS | 2.5 | NS | |
| A0A126TGR5 | ATP synthase subunit alpha | NS | 2.0 | NS | |
| B7FN64 | NADH dehydrogenase | NS | 0.4 | NS | |
| G7JIL4 | V-type proton ATPase subunit a | NS | 2.5 | 0.3 | |
| G7I9M9 | ATP synthase D chain | NS | 5.0 | NS | |
| Phagosome | A0A072VSL4 | Archaeal/vacuolar-type H+-ATPase subunit B | NS | NS | 0.4 |
| B7FMK2 | Archaeal/vacuolar-type H+-ATPase subunit E | NS | NS | 3.3 | |
| G7KSI7 | Archaeal/vacuolar-type H+-ATPase subunit B | NS | NS | 0.3 | |
| G7LIN7 | Tubulin beta-1 chain | NS | NS | 5.0 | |
| Plant–pathogen interaction | B7FNA2 | EF hand calcium-binding family protein | NS | NS | 2.5 |
| G7I7Q4 | Heat shock protein 81-2 | NS | 2.5 | 0.2 | |
| G7IDZ4 | Heat shock protein 81-2 | NS | NS | 0.1 | |
| A0A072U9J1 | Heat shock protein 81-2 | 2.5 | 5.0 | 0.3 | |
| Proteasome | A0A072TQB8 | Glyceraldehyde-3-phosphate dehydrogenase | NS | NS | 10 |
| B7FGZ8 | Proteasome subunit beta type | NS | NS | 2.5 | |
| G7JTX3 | 6S proteasome regulatory subunit S2 1B | NS | 5.0 | 0.3 | |
| I3RZQ6 | Proteasome subunit alpha type | NS | NS | 3.3 | |
| I3SSX1 | Proteasome subunit alpha type | NS | NS | 2.5 | |
| alpha-linolenic acid metabolism | G7J5N1 | Uncharacterized protein | 3.3 | NS | NS |
| Q711Q9 | Allene oxide cyclase | 2.5 | 0.1 | NS | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Zhou, Q.; Mao, P. Ultrastructural and Photosynthetic Responses of Pod Walls in Alfalfa to Drought Stress. Int. J. Mol. Sci. 2020, 21, 4457. https://doi.org/10.3390/ijms21124457
Wang H, Zhou Q, Mao P. Ultrastructural and Photosynthetic Responses of Pod Walls in Alfalfa to Drought Stress. International Journal of Molecular Sciences. 2020; 21(12):4457. https://doi.org/10.3390/ijms21124457
Chicago/Turabian StyleWang, Hui, Qingping Zhou, and Peisheng Mao. 2020. "Ultrastructural and Photosynthetic Responses of Pod Walls in Alfalfa to Drought Stress" International Journal of Molecular Sciences 21, no. 12: 4457. https://doi.org/10.3390/ijms21124457