Skin Mycobiome of Psoriasis Patients is Retained during Treatment with TNF and IL-17 Inhibitors

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Patient Background and Sequences of Fungi Detected in Skin Samples
2.2. Taxonomic Analysis of Fungi (Upper Rank)
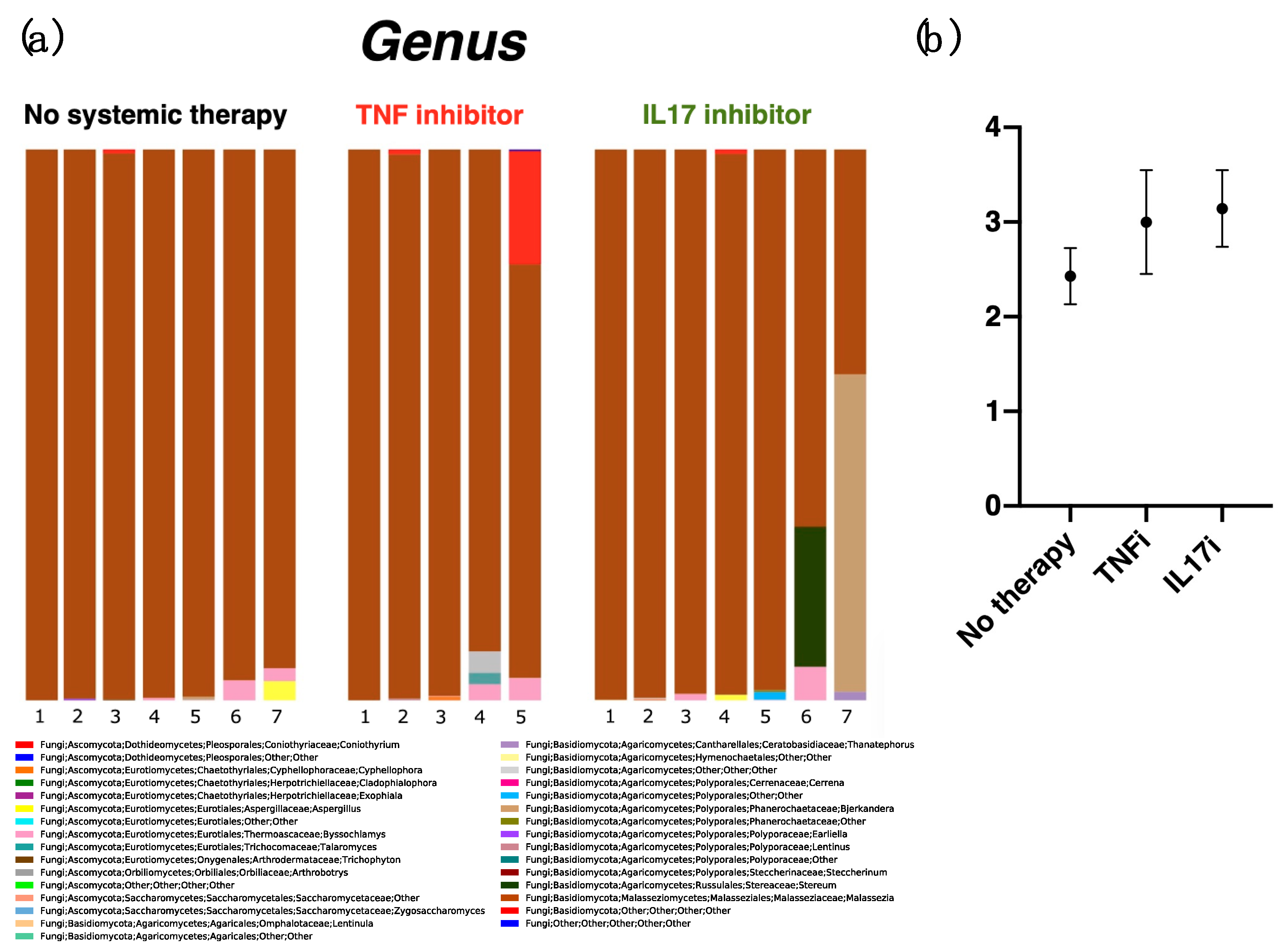
2.3. Diversity at the Genus Level
2.4. Assessment of Malassezia Species
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Sample Collection
4.3. Fungal ITS1 Deep Sequencing, Bioinformatic Analysis, and Taxonomic Assignment
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TNF | Tumor necrosis factor |
| IL | Interleukin |
| ITS | Internal transcribed spacer |
| M. | Malassezia |
| PASI | Psoriasis area and severity index |
| TNFi | Tumor necrosis factor inhibitors |
| IL-17i | Interleukin-17 inhibitors |
| OTU | Operational taxonomic units |
References
- Schon, M.P.; Boehncke, W.H. Psoriasis. N. Engl. J. Med. 2005, 352, 1899–1912. [Google Scholar]
- Hawkes, J.E.; Yan, B.Y.; Chan, T.C.; Krueger, J.G. Discovery of the IL-23/IL-17 Signaling Pathway and the Treatment of Psoriasis. J. Immunol. 2018, 201, 1605–1613. [Google Scholar] [CrossRef]
- Kuwatsuka, S.; Koike, Y.; Asai, M.; Sato, Y.; Murota, H. Numerous plasmacytoid dendritic cell infiltration in HIV-associated psoriasis relieved only with antiretroviral therapy. J. Dermatol. 2018, 45, 1126–1129. [Google Scholar]
- Kaneko, S.; Tsuruta, N.; Yamaguchi, K.; Miyagi, T.; Takahashi, K.; Higashi, Y.; Morizane, S.; Nomura, H.; Yamaguchi, M.; Hino, R.; et al. Mycobacterium tuberculosis infection in psoriatic patients treated with biologics: Real-world data from 18 Japanese facilities. J. Dermatol. 2019, 47, 128–132. [Google Scholar]
- Koike, Y.; Fujiki, Y.; Higuchi, M.; Fukuchi, R.; Kuwatsuka, S.; Murota, H. An interleukin-17 inhibitor successfully treated a complicated psoriasis and psoriatic arthritis patient with hepatitis B virus infection and end-stage kidney disease on hemodialysis. JAAD Case Rep. 2019, 5, 150–152. [Google Scholar]
- Kamata, M.; Tada, Y. Safety of biologics in psoriasis. J. Dermatol. 2018, 45, 279–286. [Google Scholar]
- Lebwohl, M.; Strober, B.; Menter, A.; Gordon, K.; Weglowska, J.; Puig, L.; Papp, K.; Spelman, L.; Toth, D.; Kerdel, F.; et al. Phase 3 Studies Comparing Brodalumab with Ustekinumab in Psoriasis. N. Engl. J. Med. 2015, 373, 1318–1328. [Google Scholar]
- Gordon, K.B.; Blauvelt, A.; Papp, K.A.; Langley, R.G.; Luger, T.; Ohtsuki, M.; Reich, K.; Amato, D.; Ball, S.G.; Braun, D.K.; et al. Phase 3 Trials of Ixekizumab in Moderate-to-Severe Plaque Psoriasis. N. Engl. J. Med. 2016, 375, 345–356. [Google Scholar] [CrossRef]
- van de Kerkhof, P.C.; Griffiths, C.E.; Reich, K.; Leonardi, C.L.; Blauvelt, A.; Tsai, T.F.; Gong, Y.; Huang, J.; Papavassilis, C.; Fox, T. Secukinumab long-term safety experience: A pooled analysis of 10 phase II and III clinical studies in patients with moderate to severe plaque psoriasis. J. Am. Acad. Dermatol. 2016, 75, 83–98.e4. [Google Scholar] [CrossRef]
- Puel, A.; Cypowyj, S.; Bustamante, J.; Wright, J.F.; Liu, L.; Lim, H.K.; Migaud, M.; Israel, L.; Chrabieh, M.; Audry, M.; et al. Chronic mucocutaneous candidiasis in humans with inborn errors of interleukin-17 immunity. Science 2011, 332, 65–68. [Google Scholar]
- Belkaid, Y.; Segre, J.A. Dialogue between skin microbiota and immunity. Science 2014, 346, 954–959. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar]
- Lande, R.; Botti, E.; Jandus, C.; Dojcinovic, D.; Fanelli, G.; Conrad, C.; Chamilos, G.; Feldmeyer, L.; Marinari, B.; Chon, S.; et al. The antimicrobial peptide LL37 is a T-cell autoantigen in psoriasis. Nat. Commun. 2014, 5, 5621. [Google Scholar] [CrossRef]
- Lowes, M.A.; Suarez-Farinas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef] [Green Version]
- Tomi, N.S.; Kranke, B.; Aberer, E. Staphylococcal toxins in patients with psoriasis, atopic dermatitis, and erythroderma, and in healthy control subjects. J. Am. Acad. Dermatol. 2005, 53, 67–72. [Google Scholar] [CrossRef]
- Raza, N.; Usman, M.; Hameed, A. Chronic plaque psoriasis: Streptococcus pyogenes throat carriage rate and therapeutic response to oral antibiotics in comparison with oral methotrexate. J. Coll. Physicians Surg. Pak. 2007, 17, 717–720. [Google Scholar]
- Gao, Z.; Tseng, C.H.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial alterations of the cutaneous bacterial biota in psoriatic lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef]
- Fahlen, A.; Engstrand, L.; Baker, B.S.; Powles, A.; Fry, L. Comparison of bacterial microbiota in skin biopsies from normal and psoriatic skin. Arch. Dermatol. Res. 2012, 304, 15–22. [Google Scholar] [CrossRef]
- Alekseyenko, A.V.; Perez-Perez, G.I.; De Souza, A.; Strober, B.; Gao, Z.; Bihan, M.; Li, K.; Methe, B.A.; Blaser, M.J. Community differentiation of the cutaneous microbiota in psoriasis. Microbiome 2013, 1, 31. [Google Scholar] [CrossRef] [Green Version]
- Tett, A.; Pasolli, E.; Farina, S.; Truong, D.T.; Asnicar, F.; Zolfo, M.; Beghini, F.; Armanini, F.; Jousson, O.; De Sanctis, V.; et al. Unexplored diversity and strain-level structure of the skin microbiome associated with psoriasis. NPJ Biofilms Microbiomes 2017, 3, 14. [Google Scholar] [CrossRef]
- Assarsson, M.; Duvetorp, A.; Dienus, O.; Soderman, J.; Seifert, O. Significant Changes in the Skin Microbiome in Patients with Chronic Plaque Psoriasis after Treatment with Narrowband Ultraviolet B. Acta Derm. Venereol. 2018, 98, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Loesche, M.A.; Farahi, K.; Capone, K.; Fakharzadeh, S.; Blauvelt, A.; Duffin, K.C.; DePrimo, S.E.; Munoz-Elias, E.J.; Brodmerkel, C.; Dasgupta, B.; et al. Longitudinal Study of the Psoriasis-Associated Skin Microbiome during Therapy with Ustekinumab in a Randomized Phase 3b Clinical Trial. J. Investig. Dermatol. 2018, 138, 1973–1981. [Google Scholar] [CrossRef] [Green Version]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Sequencing, N.I.S.C.C.; et al. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Honnavar, P.; Chakrabarti, A.; Dogra, S.; Singh, P.; Handa, S. Association of Malassezia species with psoriatic lesions. Mycoses 2014, 57, 483–488. [Google Scholar] [CrossRef]
- Takemoto, A.; Cho, O.; Morohoshi, Y.; Sugita, T.; Muto, M. Molecular characterization of the skin fungal microbiome in patients with psoriasis. J. Dermatol. 2015, 42, 166–170. [Google Scholar] [CrossRef]
- Conti, H.R.; Gaffen, S.L. IL-17-Mediated Immunity to the Opportunistic Fungal Pathogen Candida albicans. J. Immunol. 2015, 195, 780–788. [Google Scholar]
- Mengesha, B.G.; Conti, H.R. The Role of IL-17 in Protection against Mucosal Candida Infections. J. Fungi (Basel) 2017, 3, 52. [Google Scholar] [CrossRef]
- Sparber, F.; De Gregorio, C.; Steckholzer, S.; Ferreira, F.M.; Dolowschiak, T.; Ruchti, F.; Kirchner, F.R.; Mertens, S.; Prinz, I.; Joller, N.; et al. The Skin Commensal Yeast Malassezia Triggers a Type 17 Response that Coordinates Anti-fungal Immunity and Exacerbates Skin Inflammation. Cell Host Microbe 2019, 25, 389–403. [Google Scholar]
- Jo, J.H.; Kennedy, E.A.; Kong, H.H. Topographical and physiological differences of the skin mycobiome in health and disease. Virulence 2017, 8, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Sugita, T.; Suzuki, M.; Goto, S.; Nishikawa, A.; Hiruma, M.; Yamazaki, T.; Makimura, K. Quantitative analysis of the cutaneous Malassezia microbiota in 770 healthy Japanese by age and gender using a real-time PCR assay. Med. Mycol. 2010, 48, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Motooka, D.; Fujimoto, K.; Tanaka, R.; Yaguchi, T.; Gotoh, K.; Maeda, Y.; Furuta, Y.; Kurakawa, T.; Goto, N.; Yasunaga, T.; et al. Fungal ITS1 Deep-Sequencing Strategies to Reconstruct the Composition of a 26-Species Community and Evaluation of the Gut Mycobiota of Healthy Japanese Individuals. Front. Microbiol. 2017, 8, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Group | Male /Female | Age 1 | Onset Age 1 | PASI Before Treatment 1 | PASI Current 1 |
|---|---|---|---|---|---|
| No-therapy group (n = 7) | 5/2 | 51.6 ± 21.4 (16–82) | 35.0 ± 19.2 (12–61) | N/A | 5.8 ± 3.6 (0.4–11.3) |
| TNFi group (n = 5) | 2/3 | 50.2 ± 16.2 (33–82) | 33.6 ± 12.6 (21–57) | 10.2 ± 5.7 (0.4–18.2) | 1.8 ± 2.1 (0–5.6) |
| IL-17i group (n = 7) | 3/4 | 56.9 ± 22.8 (16–83) | 38.1 ± 20.3 (12–65) | 27.2 ± 16.6 (4.3–59.1) | 0.2 ± 0.5 (0–1.5) |
| Group | Average Reads (±S.D.) |
|---|---|
| No-therapy group (n = 7) | 34,523 (±17,986) |
| TNFi group (n = 5) | 40,301 (±19,250) |
| IL-17i group (n = 7) | 38,349 (±22,162) |
| Gender | M. restricta | M. globosa |
|---|---|---|
| Male (n = 10) | 10 | 0 |
| Female (n = 9) | 5 | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koike, Y.; Kuwatsuka, S.; Nishimoto, K.; Motooka, D.; Murota, H. Skin Mycobiome of Psoriasis Patients is Retained during Treatment with TNF and IL-17 Inhibitors. Int. J. Mol. Sci. 2020, 21, 3892. https://doi.org/10.3390/ijms21113892
Koike Y, Kuwatsuka S, Nishimoto K, Motooka D, Murota H. Skin Mycobiome of Psoriasis Patients is Retained during Treatment with TNF and IL-17 Inhibitors. International Journal of Molecular Sciences. 2020; 21(11):3892. https://doi.org/10.3390/ijms21113892
Chicago/Turabian StyleKoike, Yuta, Sayaka Kuwatsuka, Katsutaro Nishimoto, Daisuke Motooka, and Hiroyuki Murota. 2020. "Skin Mycobiome of Psoriasis Patients is Retained during Treatment with TNF and IL-17 Inhibitors" International Journal of Molecular Sciences 21, no. 11: 3892. https://doi.org/10.3390/ijms21113892