Genome-Wide Identification and Characterization of the UBP Gene Family in Moso Bamboo (Phyllostachys edulis)

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of PeUBP Genes in Moso Bamboo
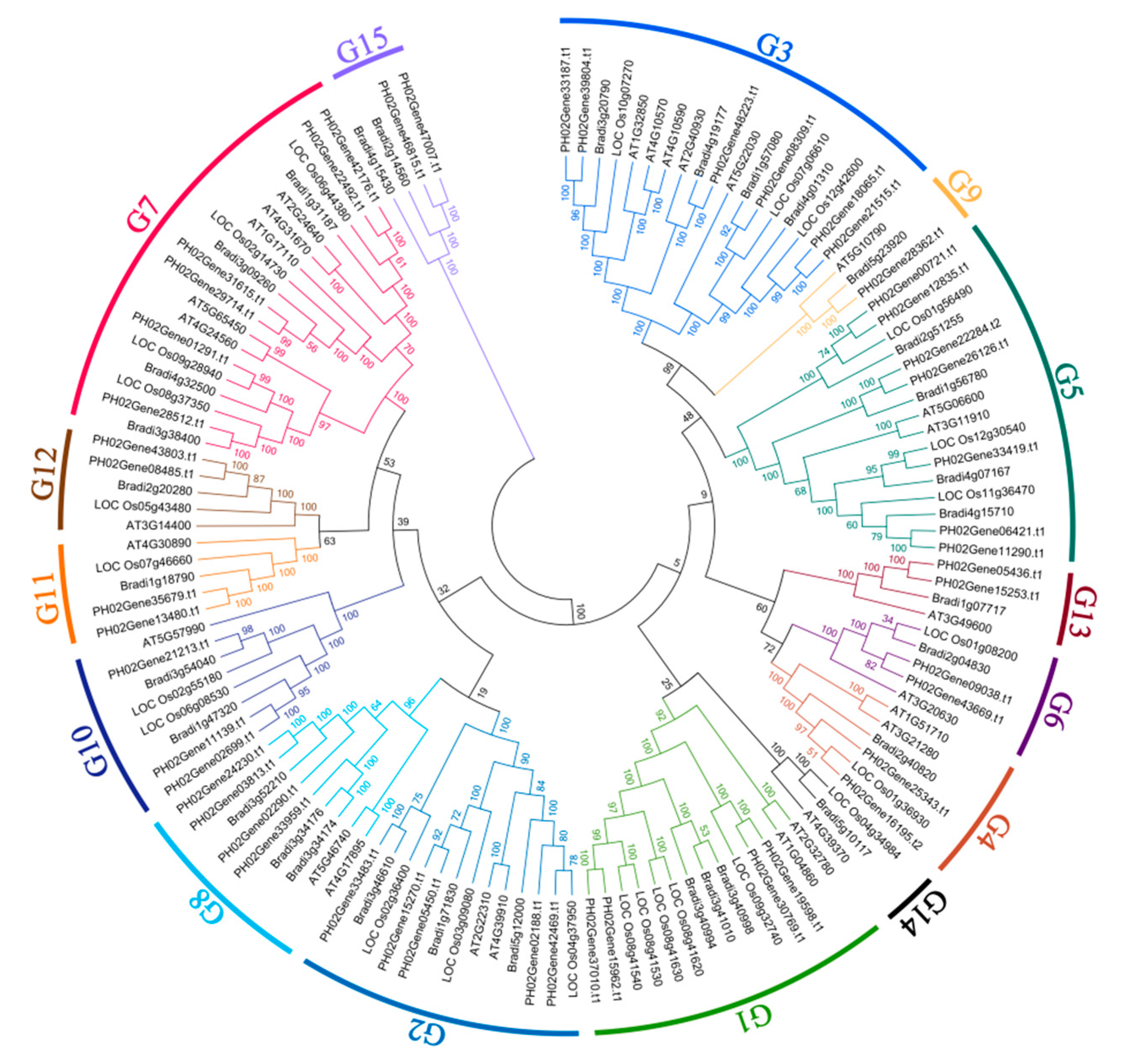
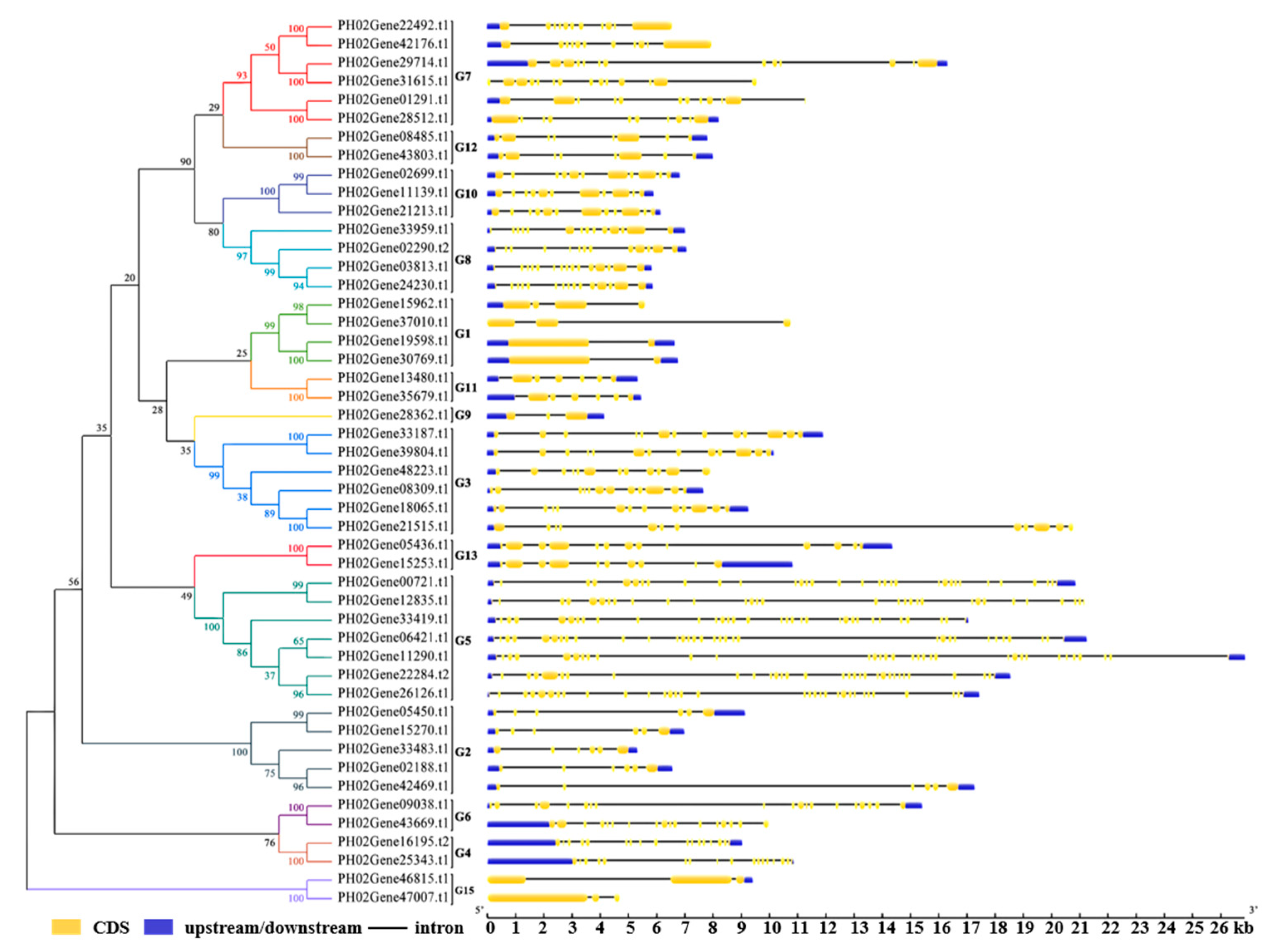
2.2. Phylogenetic Analysis of the PeUBP Genes and Identification of Exon-Intron Structure
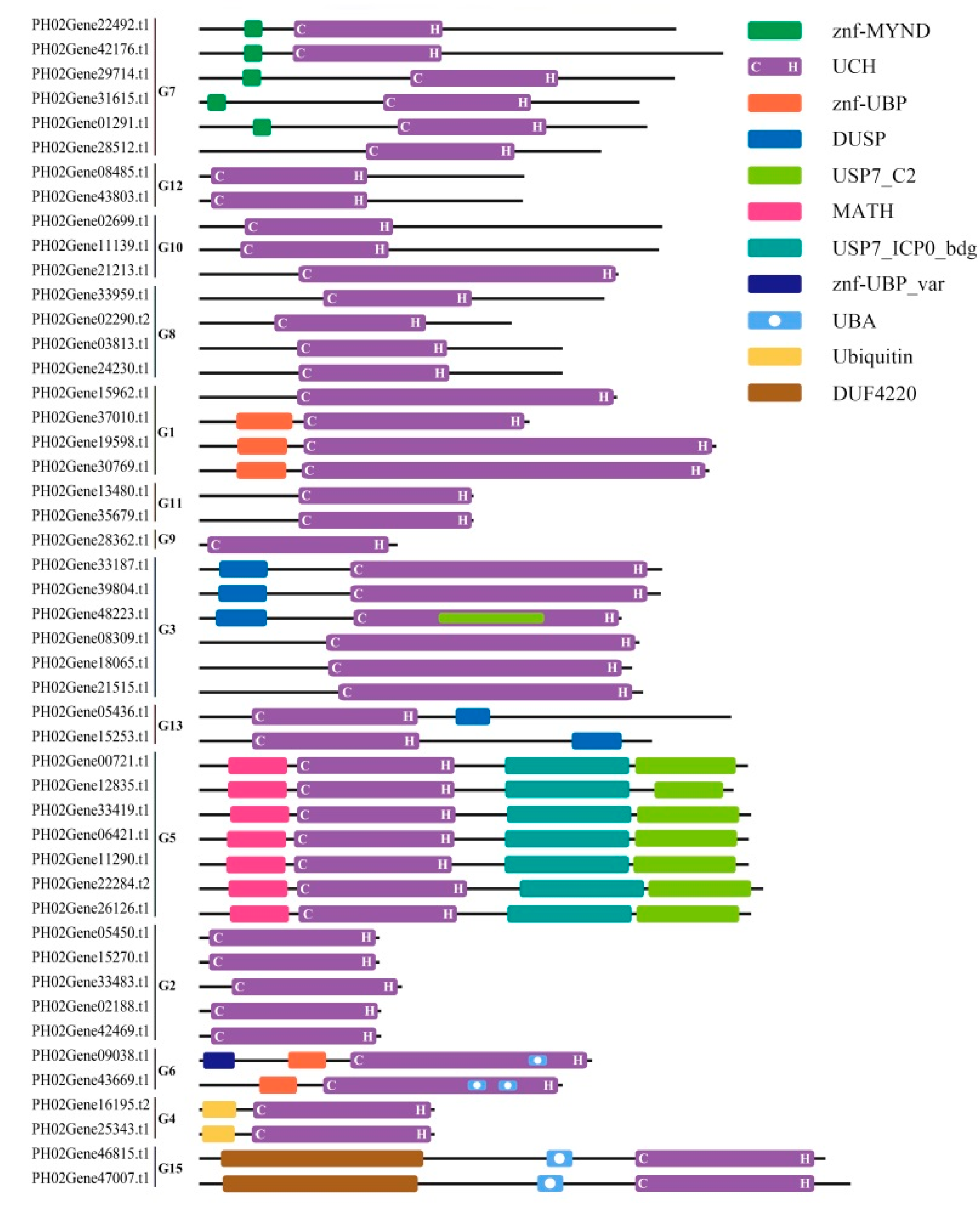
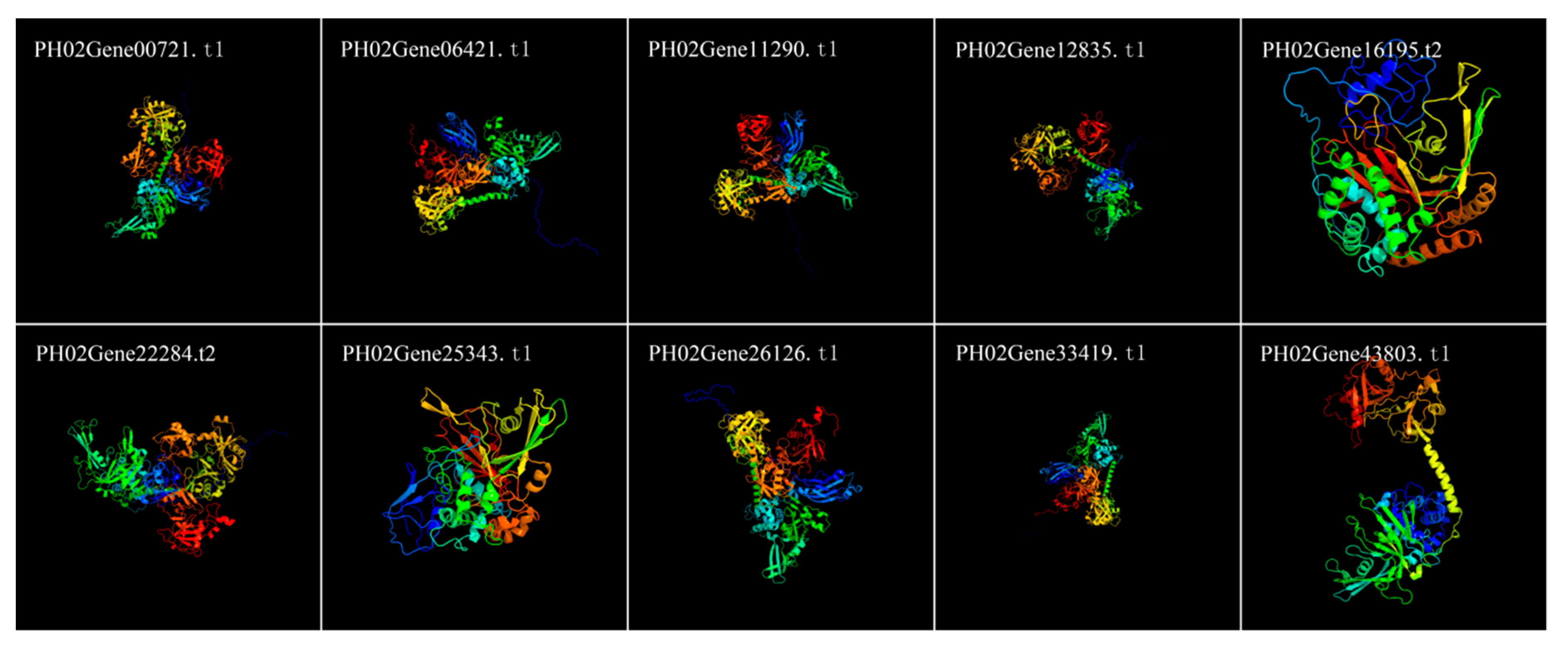
2.3. Identification of Conserved Sequence Motifs and Determination of Homology Modeling in Moso Bamboo UBP Genes
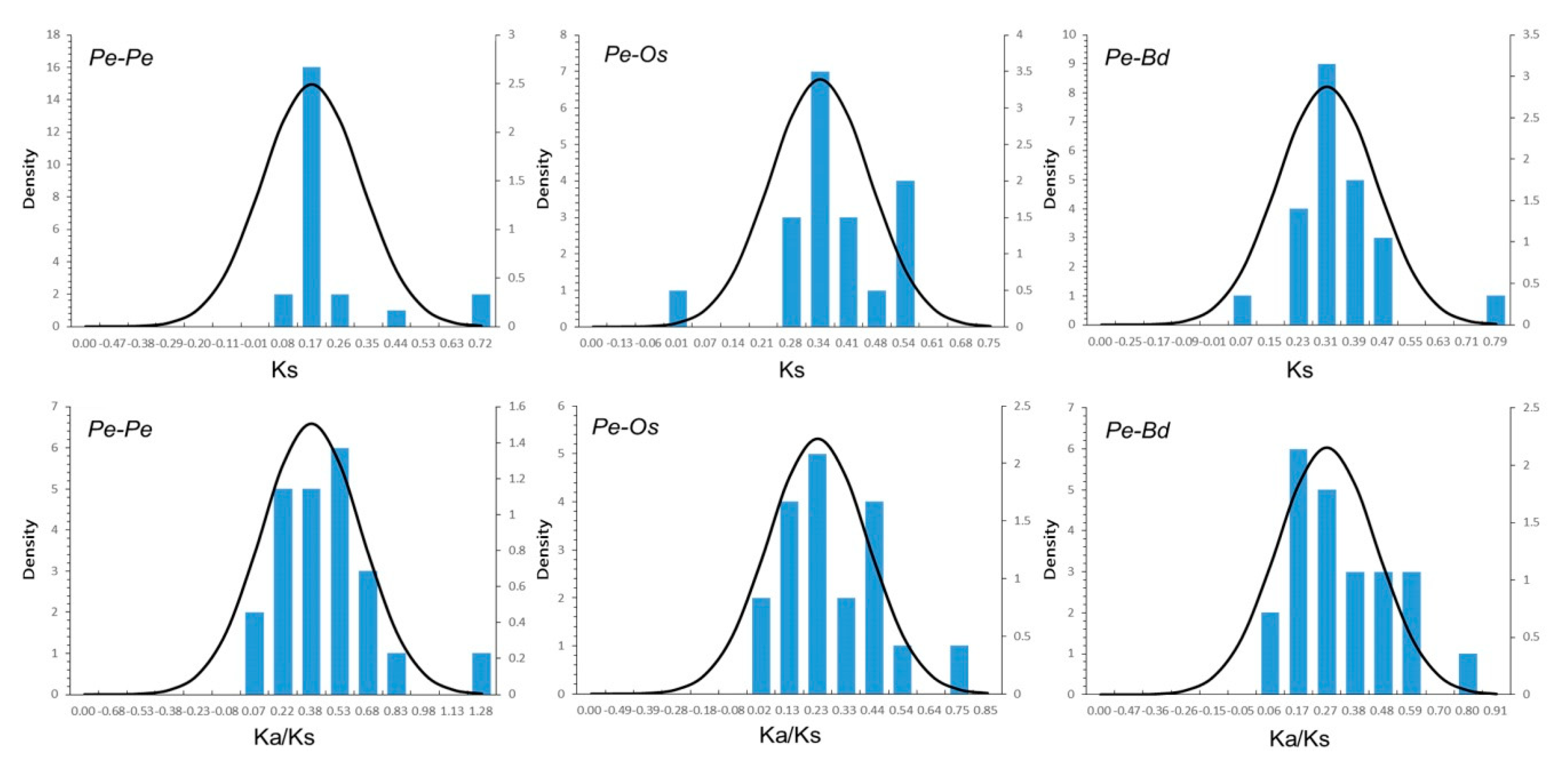
2.4. Evolutionary and Divergence Patterns of the UBP Genes in Moso Bamboo, Rice, and B. distachyon
2.5. PeUBP Expression Levels in Different Tissues
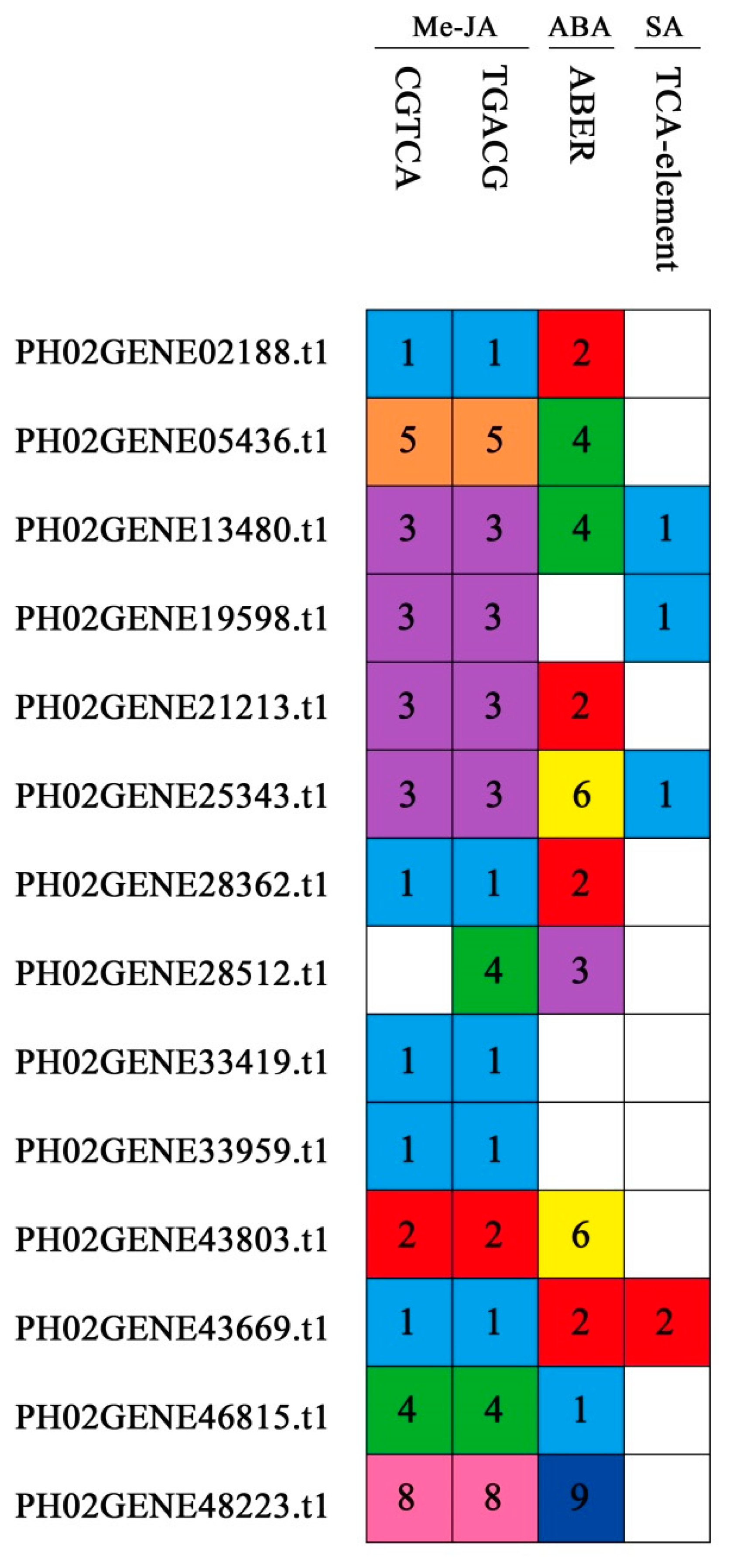
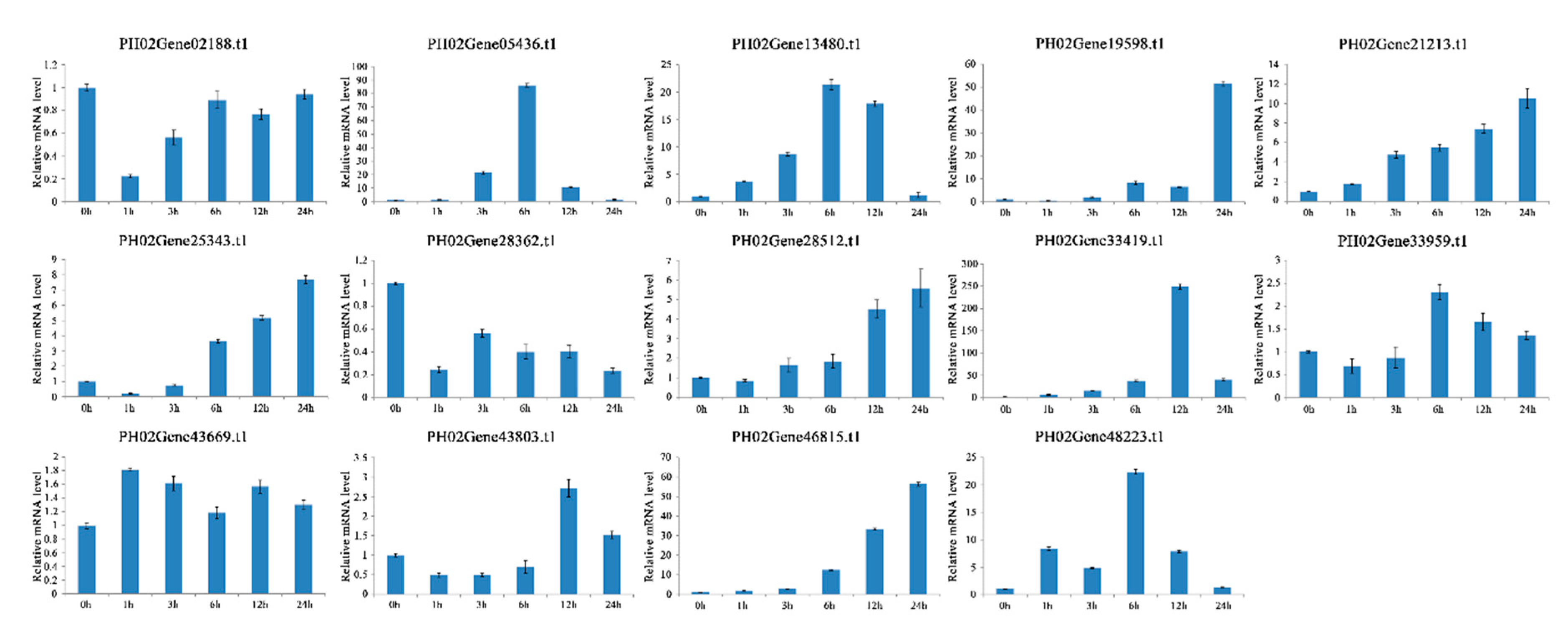
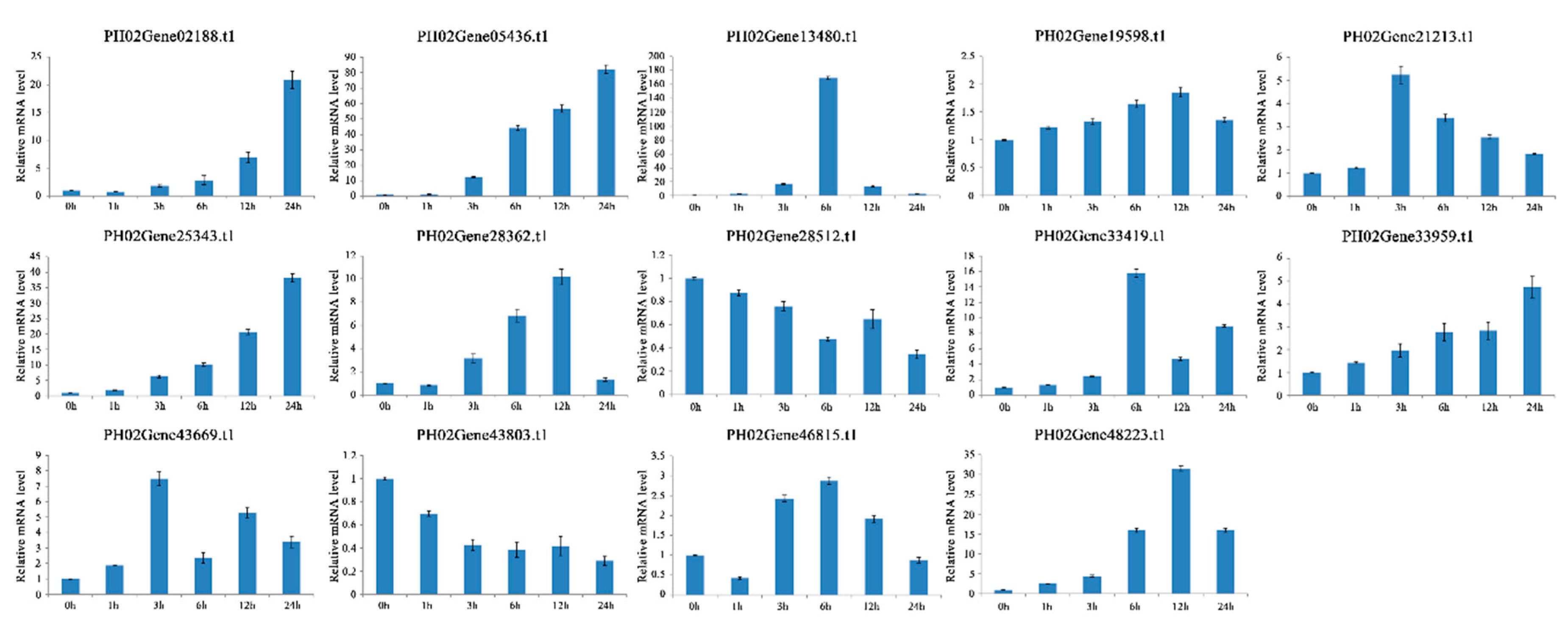
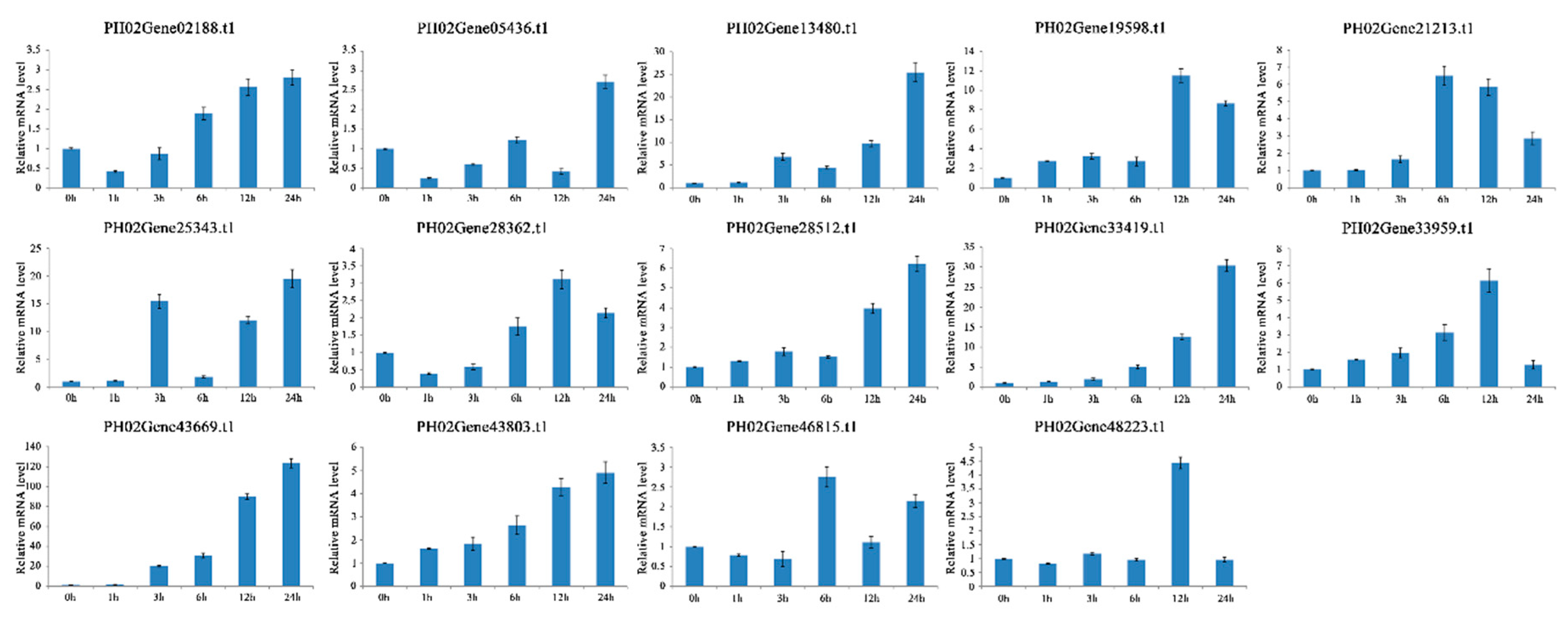
2.6. Expression Profiles of PeUBP Genes under Abscisic Acid (ABA), Methyl Jasmonate (MeJA), and Salicylic Acid (SA) Treatments
3. Discussion
3.1. UBPs in Moso Bamboo
3.2. Divergence of UBPs in Moso Bamboo, Rice, and Brachypodium
3.3. The Potential Functions of UBPs in Development and Stress-Responses in Moso Bamboo
4. Materials and Methods
4.1. Identification of Moso Bamboo PeUBP Genes
4.2. PeUBP Gene Alignments and Phylogenetic Analysis
4.3. Gene Structure Analysis
4.4. Protein Structure Analyses
4.5. Calculation of Ka/Ks Values
4.6. Putative Promoter Region Analysis of PeUBP Genes
4.7. Plant Material and Growth Conditions
4.8. PeUBP Expression Levels in Different Tissues
4.9. RNA Isolation and qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| UBPs | Ubiquitin-specific proteases |
| DUBs | Deubiquitinases/Deubiquitinating enzymes |
| ABA | Abscisic acid |
| MeJA | Methyl jasmonate |
| SA | Salicylic acid |
| E1 | Ubiquitin activating enzyme |
| E2 | Ubiquitin conjugating enzyme |
| E3 | Ubiquitin ligase= |
| UCH | Carboxyl-terminal hydrolase |
| Cys | Cysteine |
| His | Histidine |
| DUSP | Domain present in UBP |
References
- Clague, M.; Coulson, J.; Urbe, S. Cellular functions of the DUBs. J. Cell Sci. 2012, 125, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isono, E.; Nagel, M. Deubiquitylating enzymes and their emerging role in plant biology. Front. Plant Sci. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickart, C. Back to the future with ubiquitin. Cell 2004, 116, 181–190. [Google Scholar] [CrossRef]
- Vierstra, R. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1999, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Atanassov, S.; Koutelou, E.; Dent, Y. The role of deubiquitinating enzymes in chromatin regulation. FEBS Lett. 2011, 585, 2016–2023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhao, J.; Cai, J.; Patil, S. Ubiquitin-specific proteases function in plant development and stress responses. Plant Mol. Biol. 2017, 94, 565–576. [Google Scholar] [CrossRef]
- Wilkinson, K. Ubiquitination and deubiquitination: Targeting of proteins for degradation by the proteasome. Semin Cell Dev. Biol. 2000, 11, 141. [Google Scholar] [CrossRef]
- Katz, E.; Isasa, M.; Crosas, B. A new map to understand deubiquitination. Biochem. Soc. Trans. 2010, 38, 21–28. [Google Scholar] [CrossRef]
- Neutzner, M.; Neutzner, A. Enzymes of ubiquitination and deubiquitination. Essays Biochem. 2012, 52, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Varshavsky, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 22, 383–387. [Google Scholar] [CrossRef]
- Yan, N.; Doelling, J.; Falbel, T.; Durski, A.; Vierstra, R. The ubiquitin-specific proteases family from Arabidopsis. AtUBP1 and 2 are required for the resistance to the amino acid analog canavanine. Plant Physiol. 2000, 124, 1828–1843. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, F.; Zhang, H.; He, H.; Ma, L.; Deng, X. Functional characterization of the Arabidopsis ubiquitin-specific protease gene family reveals specific role and redundancy of individual members in development. Plant J. 2008, 55, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, K. Regulation of ubiquitin-dependent processes by deubiquitinating enzymes. FASEB J. 1997, 11, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.; Hong, J.; Cho, Y.; Yang, S.; An, G.; Kim, W. Structure and expression of OsUBP6, an ubiquitin-specific protease 6 homolog in rice (Oryza sativa L.). Mol. Cells. 2009, 28, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, N.; Chen, L.; Xu, Y.; Li, Y.; Zhang, Y.; Li, C.; Li, Y. The ubiquitin receptor DA1 regulates seed and organ size by modulating the stability of the ubiquitin-specific protease UBP15/SOD2 in Arabidopsis. Plant Cell 2014, 26, 665–677. [Google Scholar] [CrossRef]
- Xu, Y.; Jin, W.; Li, N.; Zhang, W.; Liu, C.; Li, C.; Li, Y. Ubiquiitin-specific protease 14 interacts with ultiraviolet-B insensitive 4 to regulate endoreduplication and cell and organ growth in Arabidopsis. Plant Cell 2016, 28, 1200–1214. [Google Scholar] [CrossRef] [PubMed]
- Doelling, J.; Yan, N.; Kurepa, J.; Walker, J.; Vierstra, R. The ubiquitin-specific protease UBP14 is essential for early embryo development in Arabidopsis thaliana. Plant J. 2001, 27, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, L.; Corke, F.; Smith, C.; Bevan, M. Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev. 2008, 22, 1331–1336. [Google Scholar] [CrossRef]
- Pan, F.; Wu, M.; Hu, W.; Liu, R.; Yan, H.; Xiang, Y. Genome-wide identification and expression analyses of the bZIP transcription factor genes in moso bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2019, 20, 2203. [Google Scholar] [CrossRef]
- Walton, A.; Stes, E.; Cybulski, N.; Van Bel, M.; Iñigo, S.; Durand, A.N.; Timmerman, E.; Heyman, J.; Pauwels, L.; De Veylder, L.; et al. It’s time for some “site”-seeing: novel tools to monitor the ubiquitin landscape in Arabidopsis thaliana. Plant Cell 2016, 28, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Nassrallah, A.; Rougée, M.; Bourbousse, C.; Drevensek, S.; Fonseca, S.; Iniesto, E.; Ait-Mohamed, O.; Deton-Cabanillas, A.F.; Zabulon, G.; Ahmed, I.; et al. DET1-mediated degradation of a SAGA-like deubiquitination module controls H2Bub homeostasis. Elife 2018, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhou, H.; Zhang, M.; Gao, Y.; Li, L.; Gao, Y.; Li, M.; Yang, Y.; Guo, Y.; Li, X. Ubiquitin-specific protease 24 negatively regulates abscisic acid signaling in Arabidopsis thaliana. Plant Cell Environ. 2016, 39, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lv, H.; Li, L.; Liu, J.; Mu, S.; Li, X.; Gao, J. Genome-wide analysis of the AP2/ERF transcription factors family and the expression patterns of DREB genes in moso bamboo (Phyllostachys edulis). PLoS ONE 2015, 10, e0126657. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Zhou, Y.; Wang, Y.; Wang, S.; Wang, S.; Hu, Y.; Bo, S.; Chen, H.; Zhou, C.; Ma, N.; et al. Insights into the bamboo genome: Syntenic relationships to rice and sorghum. Chin. Bull. Bot. 2010, 52, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Amerik, A.; Hochstrasser, M. Mechanism and function of deubiquitinating enzymes. Biochim. Biophys. Acta 2004, 1695, 189–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Song, W.; Wei, S.; Zheng, Y.; Chen, Z.; Han, J.; Zhang, H.; Luo, J.; Qin, Y.; Xu, Z.; et al. Characterization of the ubiquitin C-terminal hydrolase and ubiquiitin-specific protease families in rice (Oryza sativa). Front Plant Sci. 2018, 15, 1636. [Google Scholar] [CrossRef] [PubMed]
- Soding, J. Protein homology detection by HMM-HMM comparison. Bioinformatics 2005, 21, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Mitra, A.; Baumgarten, A.; Young, D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.; Karlowski, W.; Pan, R.; Tzeng, Y.; Mayer, K.; Li, W. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- An, Z.; Liu, Y.; Ou, Y.; Li, J.; Zhang, B.; Sun, D.; Sun, Y.; Tang, W. Regulation of the stability of RGF1 receptor by the ubiquitin-specific proteases UBP12/UBP13 is critical for root meristem maintenance. Proc. Natl. Acad. Sci. USA 2018, 115, 1123–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Lu, F.; Li, Y.; Xue, Y.; Kang, Y.; Zhang, S.; Qiu, Q.; Cui, X.; Zheng, S.; Liu, B.; et al. Ubiquitin-specific proteases UBP12 and UBP13 act in circadian clock and photoperiodic flowering regulation in Arabidopsis. Plant Physiol. 2013, 162, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Derkacheva, M.; Liu, S.; Figueiredo, D.; Gentry, M.; Mozgova, I.; Nanni, P.; Tang, M.; Mannervik, M.; Köhler, C.; Hennig, L. H2A deubiquitinases UBP12/13 are part of the Arabidopsis polycomb group protein system. Nat. Plants 2016, 2, 16126. [Google Scholar] [CrossRef] [PubMed]
- Chico, J.; Fernandez-Barbero, G.; Chini, A.; Fernandez-Calvo, P.; Diez-Diaz, M.; Solano, R. Repression of jasmonate-dependent defenses by shade involves differential regulation of protein stability of MYC transcription factors and their JAZ repressors in Arabidopsis. Plant Cell 2014, 26, 1967–1980. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.; Cutler, S.; Sheen, J.; Rodriguez, P.; Zhu, J. In vitro reconstitution of an abscisic acid signaling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Mehar, F.; Per, S.; Anjum, N.; Khan, N. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front Plant Sci. 2015, 6, 642. [Google Scholar] [CrossRef] [PubMed]
- Le, D.; Nishiyama, R.; Watanabe, Y.; Vankova, R.; Tanaka, M.; Seki, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L. Identification and expression analysis of cytokinin metabolic genes in soybean under normal and drought conditions in relation to cytokinin levels. PLoS ONE 2012, 7, e42411. [Google Scholar] [CrossRef]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, e11. [Google Scholar] [CrossRef]
- Eduardo, M.; Sara, F. Plant deubiquitinases and their role in the control of gene expression through modification of histones. Front Plant Sci. 2014, 8, 2274. [Google Scholar] [CrossRef]
- Opanowicz, M.; Vain, P.; Draper, J.; Parker, D.; Doonan, H. Brachypodium distachyon: Making hay with a wild grass. Trends Plant Sci. 2008, 13, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Bowers, E.; Chapman, A.; Rong, J.; Paterson, H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Steinmetz, L.; Gu, X.; Li, W. Role of duplicate genes in genetic robustness against null mutations. Nature 2003, 421, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Li, W.F.; Perry, P.J.; Prafulla, N.N.; Schmidt, W. Ubiquitin-specific protease 14 547 (UBP14) is involved in root responses to phosphate deficiency in Arabidopsis. Mol. Plant 2010, 3, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, M.; Li, F.; Gao, Y.; Chen, F.; Xiang, Y. TCP transcription factors in moso bamboo (Phyllostachys edulis): Genome-wide identification and expression analysis. Front Plant Sci. 2018, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchishinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Díaz, M.; Tomas, L.; Liliana, C.; Adriano, N.-N.; Jorge, R.; Laura, J.; Miren, A.; Alejandra, R.-F. Methyl jasmonate: An alternative for improving the quality and health properties of fresh fruits. Molecules 2016, 21, 567. [Google Scholar]
- Jeong, J.; Jung, C.; Seo, J.; Kim, J.; Chua, N. The deubiquitinating enzymes UBP12 and UBP13 positively regulate MYC2 levels in jasmonate responses. Plant Cell 2017, 29, 1406–1424. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Gibson, T.; Plewniak, F.; Jeanmougin, F.; Higgins, D. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Mistry, J.; Schuster-Böckler, B.; Griffiths-Jones, S.; Hollich, V.; Lassmann, T.; Moxon, S.; Marshall, M.; Khanna, A.; Durbin, R.; et al. Pfam: Clans, web tools and services. Nucleic Acids Res. 2006, 34, D247–D251. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lou, Y.; Sun, H.; Li, L.; Wang, L.; Dong, L.; Gao, Z. Transcriptome and comparative gene expression analysis of Phyllostachys edulis in response to high light. BMC Plant Biol. 2016, 16, 34. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Cheng, Z.; Xie, L.; Li, X.; Li, J.; Mu, S.; Gao, J. The R2R3 MYB gene family in Phyllostachys edulis: Genome-wide analysis and identification of stress or development-related R2R3MYBs. Front. Plant Sci. 2018, 9, 738. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction, and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van, D.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Mao, S.; Gao, Y.; Wu, D.; Cui, Y.; Li, J.; Qian, W. Genome-wide identification and expression analysis of WRKY transcription factors under multiple stresses in Brassica napus. PLoS ONE 2016, 11, e0157558. [Google Scholar] [CrossRef]
- Wu, M.; Li, Y.; Chen, D.; Liu, H.; Zhu, D.; Xiang, Y. Genome-wide identification and expression analysis of the IQD gene family in moso bamboo (Phyllostachys edulis). Sci. Rep. 2016, 6, 24520. [Google Scholar] [CrossRef]
- Fan, C.; Ma, J.; Guo, Q.; Li, X.; Wang, H.; Lu, M. Selection of reference genes for quantitative real-time PCR in bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e56573. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Location | CDS Length (bp) | Size (aa) | Protein | pl | Exons |
|---|---|---|---|---|---|---|
| MW (Da) | ||||||
| PH02Gene00721.t1 | 16: 30179141-30199136 | 3333 | 1110 | 129890.52 | 5.57 | 32 |
| PH02Gene01291.t1 | 18: 35966647-35977509 | 2721 | 906 | 99762.48 | 6.22 | 12 |
| PH02Gene02188.t1 | 24:40155854-40161467 | 1107 | 368 | 41908.50 | 5.88 | 6 |
| PH02Gene02290.t2 | 23:13745085-13751574 | 1896 | 631 | 69325.70 | 4.85 | 13 |
| PH02Gene02699.t1 | 6:13203375-13209603 | 2823 | 940 | 102597.05 | 9.36 | 11 |
| PH02Gene03813.t1 | 3:84811131-84816494 | 2223 | 740 | 82069.42 | 4.92 | 15 |
| PH02Gene05436.t1 | 15:67480992-67493851 | 3228 | 1075 | 119371.13 | 6.08 | 13 |
| PH02Gene05450.t1 | 15:67882508-67890349 | 1101 | 366 | 42022.55 | 5.86 | 6 |
| PH02Gene06421.t1 | 7:52268122-52288354 | 3354 | 1117 | 131095.16 | 5.62 | 32 |
| PH02Gene08309.t1 | 4:54406565-54413534 | 2673 | 890 | 99416.65 | 5.55 | 12 |
| PH02Gene08485.t1 | 9:53943713-53950726 | 1971 | 656 | 71789.57 | 8.98 | 8 |
| PH02Gene09038.t1 | 16:113303229-113317987 | 2383 | 795 | 88955.33 | 5.27 | 19 |
| PH02Gene11139.t1 | 8:10275805-10281089 | 2805 | 934 | 101955.24 | 9.34 | 11 |
| PH02Gene11290.t1 | 1:21490093-21516076 | 3354 | 1117 | 131044.10 | 5.59 | 32 |
| PH02Gene12835.t1 | 14:60797306-60818319 | 3249 | 1082 | 126562.99 | 5.71 | 30 |
| PH02Gene13480.t1 | 10:1249930-1254111 | 1662 | 553 | 60007.42 | 8.70 | 7 |
| PH02Gene15253.t1 | 21:101647723-101655588 | 2748 | 915 | 101640.25 | 6.34 | 10 |
| PH02Gene15270.t1 | 21:102143034-102149220 | 1101 | 366 | 41960.49 | 6.13 | 6 |
| PH02Gene15962.t1 | 13:51305905-51310945 | 2547 | 848 | 93,234.54 | 4.94 | 4 |
| PH02Gene16195.t2 | 14:31330299-31338713 | 1425 | 474 | 53,211.28 | 5.87 | 17 |
| PH02Gene18065.t1 | 11:42415269-42423636 | 2643 | 880 | 98195.73 | 5.11 | 13 |
| PH02Gene19598.t1 | 3:10292006-10297292 | 3147 | 1048 | 113926.14 | 5.19 | 2 |
| PH02Gene21213.t1 | 3:99405737-99411540 | 2979 | 992 | 109834.65 | 8.62 | 12 |
| PH02Gene21515.t1 | 12:47774473-47795019 | 2694 | 897 | 100426.26 | 5.37 | 12 |
| PH02Gene22284.t2 | 10:28962334-28980176 | 3429 | 1142 | 134013.04 | 5.66 | 31 |
| PH02Gene22492.t1 | 6:34432327-34438439 | 2907 | 968 | 106038.69 | 5.96 | 11 |
| PH02Gene24230.t1 | 2835:25003-30357 | 2217 | 738 | 81849.15 | 5.09 | 15 |
| PH02Gene25343.t1 | 16:69739339-69750041 | 1425 | 474 | 53184.22 | 6.01 | 17 |
| PH02Gene26126.t1 | 4:53402592-53419429 | 3360 | 1119 | 131489.97 | 5.71 | 32 |
| PH02Gene28362.t1 | 23:7647625-7650488 | 1200 | 399 | 45414.32 | 7.91 | 3 |
| PH02Gene28512.t1 | 13:58209169-58217387 | 2439 | 812 | 89586.32 | 6.69 | 10 |
| PH02Gene29714.t1 | 20:29015633-29030162 | 2889 | 962 | 107226.58 | 7.13 | 13 |
| PH02Gene30769.t1 | 18:30083038-30088423 | 3111 | 1036 | 113047.92 | 5.11 | 2 |
| PH02Gene31615.t1 | 17:78162911-78172473 | 2673 | 890 | 98449.22 | 6.41 | 14 |
| PH02Gene33187.t1 | 13:102796122-102807090 | 2808 | 935 | 105880.58 | 5.04 | 13 |
| PH02Gene33419.t1 | 12:32620099-32636788 | 3357 | 1118 | 131431.81 | 5.71 | 32 |
| PH02Gene33483.t1 | 17:40198974-40203749 | 1230 | 409 | 46321.46 | 5.85 | 6 |
| PH02Gene33959.t1 | 5:54814061-54820578 | 2469 | 822 | 92602.24 | 5.49 | 14 |
| PH02Gene35679.t1 | 4:3711703-3715914 | 1662 | 553 | 60216.88 | 8.88 | 7 |
| PH02Gene37010.t1 | 18:22571046-22581812 | 2004 | 667 | 73289.87 | 9.21 | 3 |
| PH02Gene39804.t1 | 5:3099809-3109660 | 2808 | 935 | 105866.37 | 5.02 | 13 |
| PH02Gene42176.t1 | 8:66644923-66652376 | 3180 | 1059 | 115732.16 | 5.90 | 11 |
| PH02Gene42469.t1 | 23:34433041-34449409 | 1107 | 368 | 41893.49 | 5.98 | 6 |
| PH02Gene43669.t1 | 14:97907481-97915260 | 1962 | 733 | 81520.86 | 4.99 | 17 |
| PH02Gene43803.t1 | 7:39435913-39442930 | 1961 | 653 | 71613.43 | 8.84 | 8 |
| PH02Gene46815.t1 | 1:15050591-15059698 | 3810 | 1269 | 144256.52 | 6.22 | 3 |
| PH02Gene47007.t1 | 1:15242214-15246894 | 3693 | 1320 | 151083.45 | 5.89 | 3 |
| PH02Gene48223.t1 | 13:32005753-32013346 | 2565 | 855 | 96768.17 | 5.70 | 12 |
| Pe-Pe | Pe-Os | Pe-Bd |
|---|---|---|
| PH02Gene05450.t1/PH02Gene15270.t1 | PH02Gene09038.t1/Os01g08200 | PH02Gene05436.t1/Bradi1g07717 |
| PH02Gene11139.t1/PH02Gene02699.t1 | PH02Gene16195.t2/Os01g36930 | PH02Gene13480.t1/Bradi1g18790 |
| PH02Gene11290.t1/PH02Gene06421.t1 | PH02Gene00721.t1/Os01g56490 | PH02Gene11139.t1/Bradi1g47320 |
| PH02Gene12835.t1/PH02Gene00721.t1 | PH02Gene29714.t1/Os02g14730 | PH02Gene26126.t1/Bradi1g56780 |
| PH02Gene15253.t1/PH02Gene05436.t1 | PH02Gene33483.t1/Os02g36400 | PH02Gene08309.t1/Bradi1g57080 |
| PH02Gene21213.t1/PH02Gene11139.t1 | PH02Gene05450.t1/Os03g09080 | PH02Gene05450.t1/Bradi1g71830 |
| PH02Gene21515.t1/PH02Gene18065.t1 | PH02Gene42469.t1/Os04g37950 | PH02Gene09038.t1/Bradi2g04830 |
| PH02Gene24230.t1/PH02Gene03813.t1 | PH02Gene43803.t1/Os05g43480 | PH02Gene47007.t1/Bradi2g14560 |
| PH02Gene25343.t1/PH02Gene16195.t2 | PH02Gene42176.t1/Os06g44380 | PH02Gene43803.t1/Bradi2g20280 |
| PH02Gene26126.t1/PH02Gene22284.t2 | PH02Gene08309.t1/Os07g06610 | PH02Gene16195.t2/Bradi2g40820 |
| PH02Gene28512.t1/PH02Gene01291.t1 | PH02Gene28512.t1/Os08g37350 | PH02Gene00721.t1/Bradi2g51255 |
| PH02Gene30769.t1/PH02Gene19598.t1 | PH02Gene15962.t1/Os08g41540 | PH02Gene39804.t1/Bradi3g20790 |
| PH02Gene31615.t1/PH02Gene29714.t1 | PH02Gene01291.t1/Os09g28940 | PH02Gene33959.t1/Bradi3g34176 |
| PH02Gene35679.t1/PH02Gene13480.t1 | PH02Gene30769.t1/Os09g32740 | PH02Gene28512.t1/Bradi3g38400 |
| PH02Gene37010.t1/PH02Gene15962.t1 | PH02Gene33187.t1/Os10g07270 | PH02Gene15962.t1/Bradi3g41010 |
| PH02Gene39804.t1/PH02Gene33187.t1 | PH02Gene06421.t1/Os11g36470 | PH02Gene33483.t1/Bradi3g46610 |
| PH02Gene42176.t1/PH02Gene22492.t1 | PH02Gene33419.t1/Os12g30540 | PH02Gene24230.t1/Bradi3g52210 |
| PH02Gene42469.t1/PH02Gene02188.t1 | PH02Gene18065.t1/Os12g42600 | PH02Gene18065.t1/Bradi4g01310 |
| PH02Gene43669.t1/PH02Gene09038.t1 | PH02Gene11290.t1/Bradi4g15710 | |
| PH02Gene43803.t1/PH02Gene08485.t1 | PH02Gene48223.t1/Bradi4g19177 | |
| PH02Gene47007.t1/PH02Gene46815.t1 | PH02Gene42469.t1/Bradi5g12000 | |
| PH02Gene28362.t1/Bradi5g23920 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, R.; Shi, Y.; Zhang, Q.; Zheng, W.; Chen, S.; Du, L.; Lu, C. Genome-Wide Identification and Characterization of the UBP Gene Family in Moso Bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2019, 20, 4309. https://doi.org/10.3390/ijms20174309
Wu R, Shi Y, Zhang Q, Zheng W, Chen S, Du L, Lu C. Genome-Wide Identification and Characterization of the UBP Gene Family in Moso Bamboo (Phyllostachys edulis). International Journal of Molecular Sciences. 2019; 20(17):4309. https://doi.org/10.3390/ijms20174309
Chicago/Turabian StyleWu, Ruihua, Yanrong Shi, Qian Zhang, Wenqing Zheng, Shaoliang Chen, Liang Du, and Cunfu Lu. 2019. "Genome-Wide Identification and Characterization of the UBP Gene Family in Moso Bamboo (Phyllostachys edulis)" International Journal of Molecular Sciences 20, no. 17: 4309. https://doi.org/10.3390/ijms20174309