Sphingosine-1-Phosphate Facilitates Skin Wound Healing by Increasing Angiogenesis and Inflammatory Cell Recruitment with Less Scar Formation

 , and
, and 
Abstract
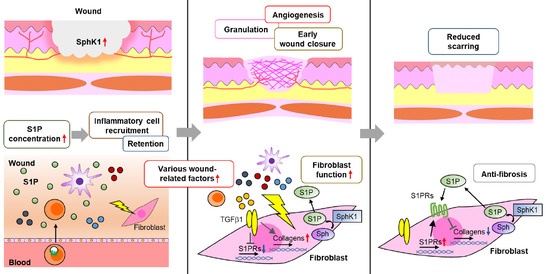
:
1. Introduction
2. Results
2.1. Longitudinal SphKs and S1PRs Expression during Mouse Wound Healing
2.2. Effect of SphK1 Gene Knockout on Wound Healing, Vasculogenesis, and Cell Proliferation
2.3. Effect of SphK1 Gene Knockout on Inflammatory Cell Recruitment during Wound Healing
2.4. Effect of Nanoparticle-Mediated Topical SphK1 Gene Delivery on Wound Healing, Vasculogenesis, and Cell Proliferation
2.5. Effect of Nanoparticle-Mediated Topical SphK1 Gene Delivery on Inflammatory Cell Recruitment during Wound Healing
2.6. Effect of SphK1 Overexpression on Granuloma Formation
2.7. Effect of SphK1 and S1PR2 Gene Expression on Scar Thickness, the Interaction between Transforming Growth Factor (TGF)-β1 and S1P
3. Discussion
4. Material and Methods
4.1. Animal Models and Wound Area Analysis
4.2. Cells and S1P Preparation
4.3. Plasmid Construction, In Vitro Transfection with sCA-Encapsulated Plasmids, and Preparation of Plasmid-sCA Ointment
4.4. Sponge Granulomas in Mice and Their Injection with Plasmid-sCA
4.5. Flow Cytometry
4.6. Quantitative RT-PCR
4.7. Immunohistochemistry, Western Blot Analysis, and Scar Thickness Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| S1P | Sphingosine-1-phosphate |
| S1PRs | Sphingosine-1-phosphate receptors |
| SphKs | Sphingosine kinases |
| TGF-β1 | Transforming growth factor-β1 |
References
- Waldorf, H.; Fewkes, J. Wound healing. Adv. Dermatol. 1995, 10, 77–96. [Google Scholar]
- Boyce, D.E.; Jones, W.D.; Ruge, F.; Harding, K.G.; Moore, K. The role of lymphocytes in human dermal wound healing. Br. J. Dermatol. 2000, 143, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ploeger, D.T.; Hosper, N.A.; Schipper, M.; Koerts, J.A.; de Rond, S.; Bank, R.A. Cell plasticity in wound healing: paracrine factors of M1/M2 polarized macrophages influence the phenotypical state of dermal fibroblasts. Cell Commun. Signal. 2013, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.; Küchler, S. Targeting inflammation and wound healing by opioids. Trends Pharmacol. Sci. 2013, 34, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, U.; Beyersdorf, N.; Weirather, J.; Podolskaya, A.; Bauersachs, J.; Ertl, G.; Kerkau, T.; Frantz, S. Activation of CD4 + T lymphocytes improves wound healing and survival after experimental myocardial infarction in mice. Circulation 2012, 125, 1652–1663. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Saeki, K.; Matsunobu, T.; Okuno, T.; Koga, T.; Sugimoto, Y.; Yokoyama, C.; Nakamizo, S.; Kabashima, K.; Narumiya, S.; et al. 12-Hydroxyheptadecatrienoic acid promotes epidermal wound healing by accelerating keratinocyte migration via the BLT2 receptor. J. Exp. Med. 2014, 211, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Lee, S.J.; Jung, Y.H.; Lee, H.J.; Han, H.J. Arachidonic acid promotes skin wound healing through induction of human MSC migration by MT3-MMP-mediated fibronectin degradation. Cell Death Dis. 2015, 6, e1750. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.Y.; Lii, C.K.; Ye, S.Y.; Li, C.C.; Lu, C.Y.; Lin, A.H.; Liu, K.L.; Chen, H.W. Docosahexaenoic acid inhibits vascular endothelial growth factor (VEGF)-induced cell migration via the GPR120/PP2A/ERK1/2/eNOS signaling pathway in human umbilical vein endothelial cells. J. Agric. Food Chem. 2014, 62, 4152–4158. [Google Scholar] [CrossRef]
- Majtan, J. Honey: An immunomodulator in wound healing. Wound Repair Regen. 2014, 22, 187–192. [Google Scholar] [CrossRef]
- ‘Izzah Ibrahim, N.; Wong, S.K.; Mohamed, I.N.; Mohamed, N.; Chin, K.Y.; Ima-Nirwana, S.; Shuid, A.N. Wound healing properties of selected natural products. Int. J. Environ. Res. Public Health 2018, 15, 2360. [Google Scholar] [CrossRef]
- Takabe, K.; Kim, R.H.; Allegood, J.C.; Mitra, P.; Ramachandran, S.; Nagahashi, M.; Harikumar, K.B.; Hait, N.C.; Milstien, S.; Spiegel, S. Estradiol induces export of sphingosine 1-phosphate from breast cancer cells via ABCC1 and ABCG2. J. Biol. Chem. 2010, 285, 10477–10486. [Google Scholar] [CrossRef] [PubMed]
- Takabe, K.; Paugh, S.W.; Milstien, S.; Spiegel, S. “Inside-out” signaling of sphingosine-1-phosphate: Therapeutic targets. Pharmacol. Rev. 2008, 60, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Aoki, H.; Ramanathan, R.; Hait, N.C.; Takabe, K. Sphingosine-1-phosphate signaling in immune cells and inflammation: roles and therapeutic potential. Mediat. Inflamm. 2016, 2016, 8606878. [Google Scholar] [CrossRef]
- Matloubian, M.; Lo, C.G.; Cinamon, G.; Lesneski, M.J.; Xu, Y.; Brinkmann, V.; Allende, M.L.; Proia, R.L.; Cyster, J.G. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 2004, 427, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Schwab, S.R.; Cyster, J.G. Finding a way out: lymphocyte egress from lymphoid organs. Nat. Immunol. 2007, 8, 1295–1301. [Google Scholar] [CrossRef]
- Schwab, S.R.; Pereira, J.P.; Matloubian, M.; Xu, Y.; Huang, Y.; Cyster, J.G. Lymphocyte sequestration through S1P lyase inhibition and disruption of S1P gradients. Science 2005, 309, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Pappu, R.; Schwab, S.R.; Cornelissen, I.; Pereira, J.P.; Regard, J.B.; Xu, Y.; Camerer, E.; Zheng, Y.-W.; Huang, Y.; Cyster, J.G.; et al. Promotion of lymphocyte egress into blood and lymph by distinct sources of sphingosine-1-phosphate. Science 2007, 316, 295–298. [Google Scholar] [CrossRef]
- Lee, M.J.; Thangada, S.; Claffey, K.P.; Ancellin, N.; Liu, C.H.; Kluk, M.; Volpi, M.; Sha’afi, R.I.; Hla, T. Vascular endothelial cell adherens junction assembly and morphogenesis induced by sphingosine-1-phosphate. Cell 1999, 99, 301–312. [Google Scholar] [CrossRef]
- Allende, M.L.; Proia, R.L. Sphingosine-1-phosphate receptors and the development of the vascular system. Biochim. Biophys. Acta 2002, 1582, 222–227. [Google Scholar] [CrossRef]
- Ledgerwood, L.G.; Lal, G.; Zhang, N.; Garin, A.; Esses, S.J.; Ginhoux, F.; Merad, M.; Peche, H.; Lira, S.A.; Ding, Y.; et al. The sphingosine 1-phosphate receptor 1 causes tissue retention by inhibiting the entry of peripheral tissue T lymphocytes into afferent lymphatics. Nat. Immunol. 2008, 9, 42–53. [Google Scholar] [CrossRef]
- Sanchez, T.; Estrada-Hernandez, T.; Paik, J.; Wu, M.; Venkataraman, K.; Brinkmann, V.; Claffey, K.; Hla, T. Phosphorylation and action of the immunomodulator FTY720 inhibits vascular endothelial cell growth factor-induced vascular permeability. J. Biol. Chem. 2003, 278, 47281–47290. [Google Scholar] [CrossRef] [PubMed]
- Marsolais, D.; Rosen, H. Chemical modulators of sphingosine-1-phosphate receptors as barrier-oriented therapeutic molecules. Nat. Rev. Drug Discov. 2009, 8, 297–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, T.; Skoura, A.; Wu, M.T.; Casserly, B.; Harrington, E.O.; Hla, T. Induction of vascular permeability by the sphingosine-1-phosphate receptor-2 (S1P2R) and its downstream effectors ROCK and PTEN. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, T.; Hla, T. Structural and functional characteristics of S1P receptors. J. Cell. Biochem. 2004, 92, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ge, J.; Tredget, E.E.; Wu, Y. The mouse excisional wound splinting model, including applications for stem cell transplantation. Nat. Protoc. 2013, 8, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Ferreras, C.; Cole, C.L.; Urban, K.; Jayson, G.C.; Avizienyte, E. Segregation of late outgrowth endothelial cells into functional endothelial CD34− and progenitor-like CD34+ cell populations. Angiogenesis 2015, 18, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yamamoto, H.; Nakanishi, H.; Yamamoto, Y.; Inoue, A.; Tei, M.; Hirose, H.; Uemura, M.; Nishimura, J.; Hata, T.; et al. Innovative Delivery of siRNA to Solid Tumors by Super Carbonate Apatite. PLoS ONE 2015, 10, e0116022. [Google Scholar] [CrossRef] [PubMed]
- Allende, M.L.; Sasaki, T.; Kawai, H.; Olivera, A.; Mi, Y.; Van Echten-Deckert, G.; Hajdu, R.; Rosenbach, M.; Keohane, C.A.; Mandala, S.; et al. Mice deficient in sphingosine kinase 1 are rendered lymphopenic by FTY720. J. Biol. Chem. 2004, 279, 52487–52492. [Google Scholar] [CrossRef] [PubMed]
- Canesso, M.C.C.; Vieira, A.T.; Castro, T.B.R.; Schirmer, B.G.A.; Cisalpino, D.; Martins, F.S.; Rachid, M.A.; Nicoli, J.R.; Teixeira, M.M.; Barcelos, L.S. Skin Wound Healing Is Accelerated and Scarless in the Absence of Commensal Microbiota. J. Immunol. 2014, 193, 5171–5180. [Google Scholar] [CrossRef]
- Tang, Q.L.; Han, S.S.; Feng, J.; Di, J.Q.; Qin, W.X.; Fu, J.; Jiang, Q.Y. Moist exposed burn ointment promotes cutaneous excisional wound healing in rats involving VEGF and bFGF. Mol. Med. Rep. 2014, 9, 1277–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghdam, S.Y.; Eming, S.A.; Willenborg, S.; Neuhaus, B.; Niessen, C.M.; Partridge, L.; Krieg, T.; Bruning, J.C. Vascular endothelial insulin/IGF-1 signaling controls skin wound vascularization. Biochem. Biophys. Res. Commun. 2012, 421, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Kyriakides, T.R.; Zhu, Y.H.; Yang, Z.; Huynh, G.; Bornstein, P. Altered extracellular matrix remodeling and angiogenesis in sponge granulomas of thrombospondin 2-null mice. Am. J. Pathol. 2001, 159, 1255–1262. [Google Scholar] [CrossRef]
- Schmid, P.; Kunz, S.; Cerletti, N.; Mcmaster, G.; Cox, D. Injury induced expression of TGF-β1 mRNA is enhanced by exogenously applied TGF-βS. Biochem. Biophys. Res. Commun. 1993, 194, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Haroon, Z.A.; Raleigh, J.A.; Greenberg, C.S.; Dewhirst, M.W. Early wound healing exhibits cytokine surge without evidence of hypoxia. Ann. Surg. 2000, 231, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Agaiby, A.D.; Dyson, M. Immuno-inflammatory cell dynamics during cutaneous wound healing. J. Anat. 1999, 195, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Efron, J.; Frankel, H.; Lazarou, S.; Wasserkrug, H.; Barbul, A. Wound healing and T-lymphocytes. J. Surg. Res. 1990, 48, 460–463. [Google Scholar] [CrossRef]
- Rani, M.; Zhang, Q.; Scherer, M.R.; Cap, A.P.; Schwacha, M.G. Activated skin γδ T-cells regulate T-cell infiltration of the wound site after burn. Innate Immun. 2015, 21, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Havran, W.L.; Jameson, J.M. Epidermal T Cells and Wound Healing. J. Immunol. 2010, 184, 5423–5428. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.; Kim, K.; Lau, L.F. The matricellular protein CCN1 mediates neutrophil efferocytosis in cutaneous wound healing. Nat. Commun. 2015, 6, 7386. [Google Scholar] [CrossRef]
- Dreymueller, D.; Denecke, B.; Ludwig, A.; Jahnen-Dechent, W. Embryonic stem cell-derived M2-like macrophages delay cutaneous wound healing. Wound Repair Regen. 2013, 21, 44–54. [Google Scholar] [CrossRef]
- Zheng, X.F.; Hong, Y.X.; Feng, G.J.; Zhang, G.F.; Rogers, H.; Lewis, M.A.O.; Williams, D.W.; Xia, Z.F.; Song, B.; Wei, X.Q. Lipopolysaccharide-induced M2 to M1 macrophage transformation for IL-12p70 production is blocked by candida albicans mediated up-regulation of EBI3 expression. PLoS ONE 2013, 8, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, H.; Bräutigam, L.; Geisslinger, G.; Pfeilschifter, J.; Frank, S. Cyclooxygenase-1-coupled prostaglandin biosynthesis constitutes an essential prerequisite for skin repair. J. Investig. Dermatol. 2003, 120, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Toyota, T.; Hirata, Y.; Ikeda, Y.; Matsuoka, K.; Sakuma, A.; Mizushima, Y. Lipo-PGE1, a new lipid-encapsulated preparation of prostaglandin E1: placebo-and prostaglandin E1-controlled multicenter trials in patients with diabetic neuropathy and leg ulcers. Prostaglandis 1993, 46, 453–468. [Google Scholar] [CrossRef]
- Barrientos, S.; Brem, H.; Stojadinovic, O.; Tomic-Canic, M. Clinical application of growth factors and cytokines in wound healing. Wound Repair Regen. 2014, 22, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef]
- He, R.; Yin, H.; Yuan, B.; Liu, T.; Luo, L.; Huang, P.; Dai, L.; Zeng, K. IL-33 improves wound healing through enhanced M2 macrophage polarization in diabetic mice. Mol. Immunol. 2017, 90, 42–49. [Google Scholar] [CrossRef]
- Chowdhury, E.; Akaike, T. High performance DNA nano-carriers of carbonate apatite: Multiple factors in regulation of particle synthesis and transfection efficiency. Int. J. Nanomed. 2007, 2, 101–106. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Bio-Rad Assay ID | Forward (5′->3′) | Reverse (5′->3′) |
|---|---|---|---|
| Col1a1 | CGATGGATTCCCGTTCGAGTA | CATTAGGCGCAGGAAGGTCA | |
| Col3a1 | GAAGTCTCTGAAGCTGATGGG | TTGCCTTGCGTGTTTGATATTC | |
| GAPDH | qMmuCEP0039581 | ||
| S1PR1 | qMmuCID0020925 | ||
| S1PR2 | qMmuCED0004722 | ||
| S1PR3 | qMmuCIP0028162 | ||
| SphK1 | qMmuCED0040475 | ||
| SphK2 | qMmuCED0039969 |
| Antigen | Company | Application | Concentration |
|---|---|---|---|
| F4/80 | Abcam | Immunohistochemistry | 1:100 |
| Ki67 | Abcam | Immunohistochemistry | 1:100 |
| CD34 | Abcam | Immunohistochemistry | 1:100 |
| V5 | Invitrogen | Western blot | 1:5000 |
| VEGF | Santa Cruz | Western blot | 1:200 |
| FGF-2 | Santa Cruz | Western blot | 1:200 |
| IGF-1 | Abcam | Western blot | 1:500 |
| GAPDH | Cell Signaling Technology | Western blot | 1:1000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, M.; Aoki, H.; Mukhopadhyay, P.; Tsuge, T.; Yamamoto, H.; Matsumoto, N.M.; Toyohara, E.; Okubo, Y.; Ogawa, R.; Takabe, K. Sphingosine-1-Phosphate Facilitates Skin Wound Healing by Increasing Angiogenesis and Inflammatory Cell Recruitment with Less Scar Formation. Int. J. Mol. Sci. 2019, 20, 3381. https://doi.org/10.3390/ijms20143381
Aoki M, Aoki H, Mukhopadhyay P, Tsuge T, Yamamoto H, Matsumoto NM, Toyohara E, Okubo Y, Ogawa R, Takabe K. Sphingosine-1-Phosphate Facilitates Skin Wound Healing by Increasing Angiogenesis and Inflammatory Cell Recruitment with Less Scar Formation. International Journal of Molecular Sciences. 2019; 20(14):3381. https://doi.org/10.3390/ijms20143381
Chicago/Turabian StyleAoki, Masayo, Hiroaki Aoki, Partha Mukhopadhyay, Takuya Tsuge, Hirofumi Yamamoto, Noriko M. Matsumoto, Eri Toyohara, Yuri Okubo, Rei Ogawa, and Kazuaki Takabe. 2019. "Sphingosine-1-Phosphate Facilitates Skin Wound Healing by Increasing Angiogenesis and Inflammatory Cell Recruitment with Less Scar Formation" International Journal of Molecular Sciences 20, no. 14: 3381. https://doi.org/10.3390/ijms20143381