Disturbance of Intracerebral Fluid Clearance and Blood–Brain Barrier in Vascular Cognitive Impairment

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
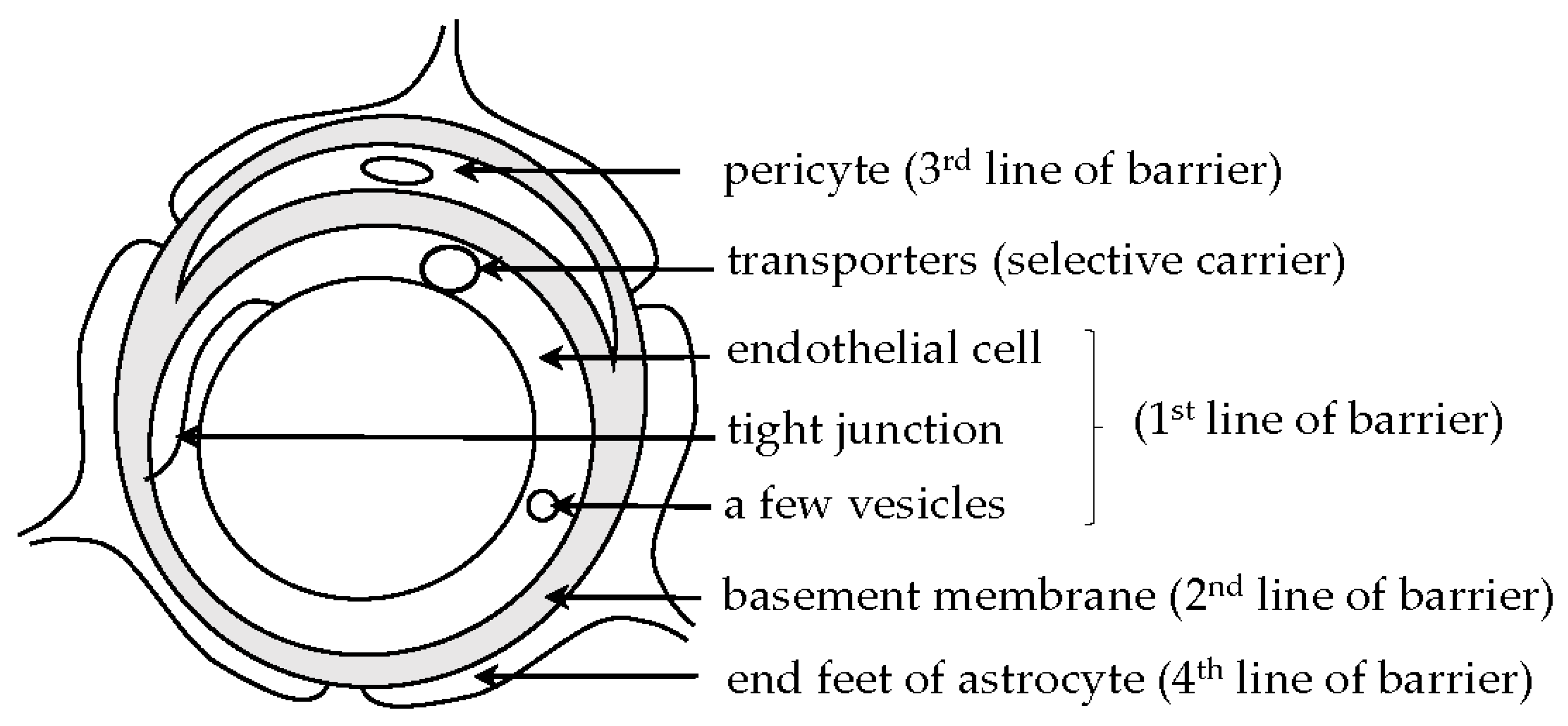
2. BBB and BCSFB
2.1. BBB and BCSFB
2.2. Circumventricular Organs (CVOs)
3. Intramural Peri-Arterial Drainage (IPAD) Pathway and Glymphatic System
4. BBB Damage in Human Brains and Experimental Animal Models for Vascular Cognitive Impairment
4.1. BBB Changes in Aging With or Without Cognitive Dysfunction
4.2. BBB Changes in Hypertension
4.3. BBB Changes in Hyperglycemia
4.4. BBB Changes in Acute Ischemia Followed by Reperfusion
4.5. BBB Changes in Chronic Hypoperfusion
4.6. BBB Changes in Hydrocephalus
5. Target Genes for Vascular Cognitive Impairment
6. Conclusions
Author Contributions
Acknowledgements
Conflicts of Interest
Abbreviation
| Aβ | amyloid-β |
| ABC | ATP-binding cassette |
| AD | Alzheimer’s disease |
| AQP4 | aquaporin 4 |
| BBB | blood–brain barrier |
| BCSFB | blood-cerebrospinal fluid barrier |
| BM | basement membrane |
| CSF | cerebrospinal fluid |
| CVO | circumventricular organ |
| FPRL1 | formylpeptide receptor-like-1 |
| GLUT | glucose transporter |
| HRP | horseradish peroxidase |
| IDE | insulin-degrading enzyme |
| IPAD | intramural peri-arterial drainage |
| ISF | interstitial fluid |
| LDLR | low-density-lipoprotein receptor |
| LRP | LDLR-related protein |
| MMP | matrix metalloproteinase |
| MRI | magnetic resonance imaging |
| 8-OHdG | 8-hydroxy-2’-deoxyguanosine |
| RAGE | receptor for advanced end products |
| SAMP | senescence-accelerated mouse prone |
| SHR | spontaneously hypertensive rat |
| SHRSP | SHR stroke-prone |
| WKY | Wistar Kyoto |
References
- Lee, B.I.; Nam, H.S.; Heu, J.H.; Kim, D.I.; The Yonsei Stroke Team. Yonsei Stroke Registry analysis of 1000 patients with acute cerebral infarction. Cerebrovasc. Dis. 2001, 12, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, J.M.; Doubal, F.; Armitage, P.; Chappell, F.; Carpenter, T.; Muñoz Maniega, S.; Farrall, A.; Sudlow, C.; Dennis, M.; Dhillon, B. Lacunar stroke is associated with diffuse blood–brain barrier dysfunction. Ann. Neurol. 2009, 65, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Topakian, R.; Barrick, T.R.; Howe, F.A.; Markus, H.S. Blood–brain barrier permeability is increased in normal-appearing white matter in patients with lacunar stroke and leukoaraiosis. J. Neurol. Neurosurg. Psychiatry 2010, 81, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, J.M.; Doubal, F.N.; Valdes-Hemandez, M.; Wang, X.; Chappell, F.M.; Shuler, K.; Armitage, P.A.; Carpenter, T.C.; Dennis, M.S. Blood–brain barrier permeability and long-term clinical and imaging outcomes in cerebral small vessel disease. Stroke 2013, 44, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Arba, F.; Leigh, R.; Inzitari, D.; Warach, S.J.; Luby, M.; Lees, K.R.; STIR/VISTA imaging collaboration. Blood–brain barrier leakage increases with small vessel disease in acute ischemic stroke. Neurology 2017, 89, 2143–2150. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood–brain barrier: From physiology to disease and back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.; Chiba, Y.; Matsumoto, K.; Murakami, R.; Fujihara, R.; Kawauchi, M.; Miyanaka, H.; Nakagawa, T. Blood–brain barrier damage in vascular dementia. Neuropathology 2016, 36, 115–124. [Google Scholar] [CrossRef]
- Ueno, M.; Chiba, Y.; Murakami, R.; Matsumoto, K.; Kawauchi, M.; Fujihara, R. Blood–brain barrier and blood-cerebrospinal fluid barrier in normal and pathological conditions. Brain Tumor Pathol. 2016, 33, 89–96. [Google Scholar] [CrossRef]
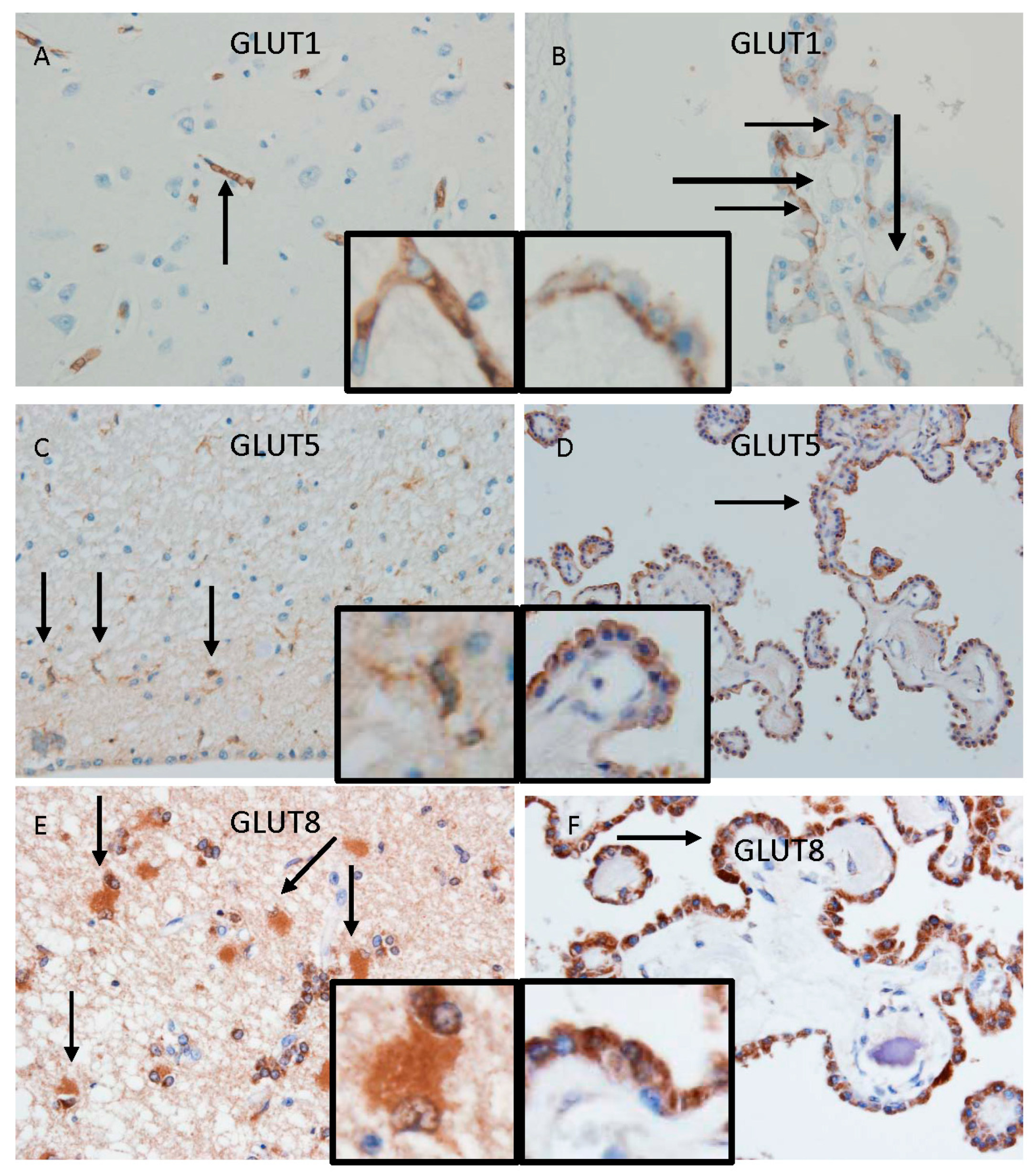
- Ueno, M.; Nishi, N.; Nakagawa, T.; Chiba, Y.; Tsukamoto, I.; Kusaka, T.; Miki, T.; Sakamoto, H.; Yamaguchi, F.; Tokuda, M. Immunoreactivity of glucose transporter 5 is located in epithelial cells of the choroid plexus and ependymal cells. Neuroscience 2014, 260, 149–157. [Google Scholar] [CrossRef]
- Murakami, R.; Chiba, Y.; Tsuboi, K.; Matsumoto, K.; Kawauchi, M.; Fujihara, R.; Mashima, M.; Kanenishi, K.; Yamamoto, T.; Ueno, M. Immunoreactivity of glucose transporter 8 is localized in the epithelial cells of the choroid plexus and in ependymal cells. Histochem. Cell Biol. 2016, 146, 231–236. [Google Scholar] [CrossRef]
- Mashima, M.; Chiba, Y.; Murakami, R.; Uemura, N.; Matsumoto, K.; Kawauchi, M.; Kanenishi, K.; Hata, T.; Ueno, M. Glucose transporter 8 immunoreactivity in astrocytic and microglial cells in subependymal areas of human brains. Neurosci. Lett. 2017, 636, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.; Chiba, Y.; Matsumoto, K.; Nakagawa, T.; Miyanaka, H. Clearance of beta-amyloid in the brain. Curr. Med. Chem. 2014, 21, 4085–4090. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Chiba, Y.; Fujihara, R.; Kubo, H.; Sakamoto, H.; Ueno, M. Immunohistochemical analysis of transporters related to clearance of amyloid-β peptides through blood-cerebrospinal fluid barrier in human brain. Histochem. Cell Biol. 2015, 144, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Daood, M.; Tsai, C.; Ahdab-Barmada, M.; Watchko, J.F. ABC transporter (P-gp/ABCB1, MRP1/ABCC1, BCRP/ABCG2) expression in the developing human CNS. Neuropediatrics 2008, 39, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.; Akiguchi, I.; Hosokawa, M.; Kotani, H.; Kanenishi, K.; Sakamoto, H. Blood–brain barrier permeability in the periventricular areas of the normal mouse brain. Acta Neuropathol. 2000, 99, 385–392. [Google Scholar] [CrossRef]
- Weller, R.O.; Subash, M.; Preston, S.D.; Mazanti, I.; Carare, R.O. Perivascular drainage of amyloid-β peptides from the brain and its failure in cerebral amyloid angiopathy and Alzheimer’s disease. Brain Pathol. 2008, 18, 253–266. [Google Scholar] [CrossRef]
- Hawkes, C.A.; Härtig, W.; Kacza, J.; Schliebs, R.; Weller, R.O.; Nicoll, J.A.; Carare, R.O. Perivascular drainage of solutes is impaired in the ageing mouse brain and in the presence of cerebral amyloid angiopathy. Acta Neuropathol. 2011, 121, 431–443. [Google Scholar] [CrossRef]
- Carare, R.O.; Hawkes, C.A.; Jeffrey, M.; Kalaria, R.N.; Weller, R.O. Cerebral amyloid angiopathy, prion angiopathy, CADASIL and spectrum of protein elimination failure angiopathies (PEFA) in neurodegenerative disease with a focus on therapy. Neuropathol. Appl. Neurobiol. 2013, 39, 593–611. [Google Scholar] [CrossRef]
- Carare, R.O.; Hawkes, C.A.; Weller, R.O. Afferent and efferent immunological pathways of the brain. Anatomy, function and failure. Brain Behav. Immun. 2014, 36, 9–14. [Google Scholar] [CrossRef]
- Diem, A.K.; Tan, M.; Bressloff, N.W.; Hawkes, C.; Morris, A.W.; Weller, R.O.; Carare, R.O. A simulation model of periarterial clearance of amyloid-β from the brain. Front. Aging Neurosci. 2016, 8, 18. [Google Scholar] [CrossRef]
- Morris, A.W.; Sharp, M.M.; Albargothy, N.J.; Fernandes, R.; Hawkes, C.A.; Verma, A.; Weller, R.O.; Carare, R.O. Vascular basement membranes as pathways for the passage of fluid into and out of the brain. Acta Neuropathol. 2016, 131, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, B.; Carare, R.O.; Bechmann, I.; Flügel, A.; Laman, J.D.; Weller, R.O. Vascular, glial, and lymphatic immune gateways of the central nervous system. Acta Neuropathol. 2016, 132, 318–338. [Google Scholar] [CrossRef] [PubMed]
- Diem, A.K.; Carare, R.O.; Weller, R.O.; Bressloff, N.W. A control mechanism for intra-mural peri-arterial drainage via astrocytes: How neuronal activity could improve waste clearance from the brain. PLoS One 2018, 13, e025276. [Google Scholar] [CrossRef] [PubMed]
- Aldea, R.; Weller, R.O.; Wilcock, D.M.; Carare, R.O.; Richardson, G. Cerebrovascular smooth muscle cells as the drivers of intramural periarterial drainage of the brain. Front. Aging Neurol. 2019, 11, 1. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Iliff, J.; Chen, M.J.; Plog, B.A.; Zeppenfeld, D.M.; Soltero, M.; Yang, L.; Singh, I.; Deane, R.; Nedergaard, M. Impairment of glymphatic pathway function promotes tau pathology after traumatic brain injury. J. Neurosci. 2014, 34, 16180–16193. [Google Scholar] [CrossRef]
- Kress, B.T.; Iliff, J.J.; Xia, M.; Wang, M.; Wei, H.S.; Zeppenfeld, D.; Xie, L.; Kang, H.; Xu, Q.; Liew, J.A.; et al. Impairment of paravascular clearance pathways in the aging brain. Ann. Neurol. 2014, 76, 845–861. [Google Scholar] [CrossRef]
- Iliff, J.J.; Goldman, S.A.; Nedergaard, M. Implications of the discovery of brain lymphatic pathways. Lancet Neurol. 2015, 14, 977–979. [Google Scholar] [CrossRef] [Green Version]
- Tarasoff-Conway, J.M.; Carare, R.O.; Osorio, R.S.; Glodzik, L.; Butler, T.; Fieremans, E.; Axel, L.; Rusinek, H.; Nicholson, C.; Zlokovic, B.V.; et al. Clearance systems in the brain – implications for Alzheimer disease. Nat. Rev. Neurol. 2015, 11, 457–470. [Google Scholar] [CrossRef]
- Wang, M.; Ding, F.; Deng, S.; Guo, X.; Wang, W.; Iliff, J.J.; Nedergaard, M. Focal solute trapping and global glymphatic pathway impairment in a murine model of multiple microinfarcts. J. Neurosci. 2017, 37, 2870–2877. [Google Scholar] [CrossRef]
- Benveniste, H.; Liu, X.; Koundal, S.; Sanggaard, S.; Lee, H.; Wardlaw, J. The glymphatic system and waste clearance with brain aging: A review. Gerontology 2018, 11, 1–14. [Google Scholar] [CrossRef]
- Verheggen, I.C.M.; van Boxtel, M.P.J.; Verhey, F.R.J.; Jansen, J.F.A.; Backes, W.H. Interaction between blood–brain barrier and glymphatic system in solute clearance. Neurosci. Biobehav. Rev. 2018, 90, 26–33. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Nam, K.I.; Song, J. The glymphatic system in diabetes-induced dementia. Front. Neurol. 2018, 9, 867. [Google Scholar] [CrossRef]
- Abbott, N.J.; Pizzo, M.E.; Preston, J.E.; Janigro, D.; Thorne, R.G. The role of brain barriers in fluid movement in the CNS: is there a ‘glymphatic’ system? Acta Neuropathol. 2018, 135, 387–407. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zlokovic, B.V. A lymphatic waste-disposal system implicated in Alzheimer’s disease. Nature 2018, 560, 172–174. [Google Scholar] [CrossRef]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood–brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 2019, 25, 270–276. [Google Scholar] [CrossRef]
- Brown, R.; Benveniste, H.; Black, S.E.; Charpak, S.; Dichgans, M.; Joutel, A.; Nedergaard, M.; Smith, K.J.; Zlokovic, B.V.; Wardlaw, J.M. Understanding the role of the perivascular space in cerebral small vessel disease. Cardiovasc. Res. 2018, 114, 1462–1473. [Google Scholar] [CrossRef]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef]
- Mawuenyega, K.G.; Sigurdson, W.; Ovod, V.; Munsell, L.; Kasten, T.; Morris, J.C.; Yarasheski, K.E.; Bateman, R.J. Decreased clearance of CNS β-amyloid in Alzheimer’s disease. Science 2010, 330, 1774. [Google Scholar] [CrossRef]
- Chesser, A.S.; Pritchard, S.M.; Johnson, G.V.W. Tau clearance mechanisms and their possible role in the pathogenesis of Alzheimer disease. Front. Neurol. 2013, 4, 122. [Google Scholar] [CrossRef]
- Farrall, A.J.; Wardlaw, J.M. Blood–brain barrier: ageing and microvascular disease-systematic review and meta-analysis. Neurobiol. Aging 2009, 30, 337–352. [Google Scholar] [CrossRef]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood–brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef]
- Ueno, M.; Sakamoto, H.; Kanenishi, K.; Onodera, M.; Akiguchi, I.; Hosokawa, M. Ultrastructural and permeability features of microvessels in the hippocampus, cerebellum and pons of senescence-accelerated mice (SAM). Neurobiol. Aging 2001, 22, 469–478. [Google Scholar] [CrossRef]
- Verhaaren, B.F.J.; Vernooij, M.W.; de Boer, R.; Hofman, A.; Niessen, W.J.; van der Lugt, A.; Ikram, M.A. High blood pressure and cerebral white matter lesion progression in the general population. Hypertension 2013, 61, 1354–1359. [Google Scholar] [CrossRef]
- Vaslievko, V.; Passos, G.; Quiring, D.; Head, E.; Kim, R.C.; Fisher, M.; Cribbs, D.H. Aging and cerebrovascular dysfunction: contribution of hypertension, cerebral amyloid angiopathy, and immunotherapy. Ann. N.Y. Acad. Sci. 2010, 1207, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Ueno, M.; Sakamoto, H.; Tomimoto, H.; Akiguchi, I.; Onodera, M.; Huang, C.L.; Kanenishi, K. Blood–brain barrier is impaired in the hippocampus of young adult spontaneously hypertensive rats. Acta Neuropathol. 2004, 107, 532–538. [Google Scholar] [CrossRef]
- Sabbatini, M.; Catalani, A.; Consoli, C.; Marletta, N.; Tomassoni, D.; Avola, R. The hippocampus in spontaneously hypertensive rats: an animal model of vascular dementia. Mech. Ageing Dev. 2002, 123, 547–559. [Google Scholar] [CrossRef]
- Starr, J.M.; Wardlaw, J.; Ferguson, K.; MacLullich, A.; Deary, I.J.; Marshall, I. Increased blood–brain barrier permeability in type II diabetes demonstrated by gadolinium magnetic resonance imaging. J. Neurol. Neurosurg. Psychiatry 2003, 74, 70–76. [Google Scholar] [CrossRef]
- Mooradian, A.D.; Haas, M.J.; Batejko, O.; Hovsepyan, M.; Feman, S.S. Statins ameliorate endothelial barrier permeability changes in the cerebral tissue of streptozotocin-induced diabetic rats. Diabetes 2005, 54, 2977–2982. [Google Scholar] [CrossRef]
- Liao, Y.J.; Ueno, M.; Nakagawa, T.; Huang, C.; Kanenishi, K.; Onodera, M.; Sakamoto, H. Oxidative damage in cerebral vessels of diabetic db/db mice. Diabetes Metab. Res. Rev. 2005, 21, 554–559. [Google Scholar] [CrossRef]
- Fujihara, R.; Chiba, Y.; Nakagawa, T.; Nishi, N.; Murakami, R.; Matsumoto, K.; Kawauchi, M.; Yamamoto, T.; Ueno, M. Albumin microvascular leakage in brains with diabetes mellitus. Microsc. Res. Tech. 2016, 79, 833–837. [Google Scholar] [CrossRef]
- Simpkins, A.N.; Dias, C.; Leigh, R.; National Institutes of Health Natural History of Stroke Investigators. Identification of reversible disruption of the human blood–brain barrier following acute ischemia. Stroke 2016, 47, 2405–2408. [Google Scholar] [CrossRef]
- Abboud, H.; Labreuche, J.; Meseguer, E.; Lavallee, P.C.; Simon, O.; Olivot, J.M.; Mazighi, M.; Dehoux, M.; Benessiano, J.; Steg, P.G.; et al. Ischemia-modified albumin in acute stroke. Cerebrovasc. Dis. 2007, 23, 216–220. [Google Scholar] [CrossRef]
- Shinnou, M.; Ueno, M.; Sakamoto, H.; Ide, M. Blood–brain barrier damage in reperfusion following ischemia in the hippocampus of the Mongolian gerbil brain. Acta Neurol. Scand. 1998, 98, 406–411. [Google Scholar] [CrossRef]
- Tomimoto, H.; Akiguchi, I.; Suenaga, T.; Nishimura, M.; Wakita, H.; Nakamura, S.; Kimura, J. Alterations of the blood–brain barrier and glial cells in white-matter lesions in cerebrovascular and Alzheimer’s disease patients. Stroke 1996, 27, 2069–2074. [Google Scholar] [CrossRef]
- Ueno, M.; Tomimoto, H.; Akiguchi, I.; Wakita, H.; Sakamoto, H. Blood–brain barrier disruption in white matter of chronic cerebral hypoperfusion. J. Cereb. Blood Flow Metab. 2002, 22, 97–104. [Google Scholar] [CrossRef]
- Ueno, M.; Tomita, S.; Nakagawa, T.; Ueki, M.; Iwanaga, Y.; Ono, J.; Onodera, M.; Huang, C.L.; Kanenishi, K.; Shimada, A.; et al. Effects of aging and HIF-1α deficiency on permeability of hippocampal vessels. Microsc. Res. Tech. 2006, 69, 29–35. [Google Scholar] [CrossRef]
- Srinivasan, V.; Braidy, N.; Chan, E.K.; Xu, Y.H.; Chan, D.K.Y. Genetic and environmental factors in vascular dementia: an update of blood brain barrier dysfunction. Clin. Exp. Pharmacol. Physiol. 2016, 43, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Romanitan, M.O.; Pepescu, B.O.; Winblad, B.; Bajenaru, O.A.; Bogdanovic, N. Occludin is overexpressed in Alzheimer’s disease and vascular dementia. J. Cell. Mol. Med. 2007, 11, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Romanitan, M.O.; Popescu, B.O.; Spulber, S.; Băjenaru, O.; Popescu, L.M.; Winblad, B.; Bogdanovic, N. Altered expression of claudin family proteins in Alzheimer’s disease and vascular dementia brains. J. Cell. Mol. Med. 2010, 14, 1088–1100. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, Y.; Ueno, M.; Ueki, M.; Huang, C.L.; Tomita, S.; Okamoto, Y.; Ogawa, T.; Ueda, N.; Maekawa, N.; Sakamoto, H. The expression of osteopontin is increased in vessels with blood–brain barrier impairment. Neuropathol. Appl. Neurobiol. 2008, 34, 145–154. [Google Scholar] [CrossRef]
- Ueno, M.; Nakagawa, T.; Nagai, Y.; Nishi, N.; Kusaka, T.; Kanenishi, K.; Onodera, M.; Hosomi, N.; Huang, C.; Yokomise, H.; et al. The expression of CD36 in vessels with blood–brain barrier impairment in a stroke-prone hypertensive model. Neuropathol. Appl. Neurobiol. 2011, 37, 727–737. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueno, M.; Chiba, Y.; Murakami, R.; Matsumoto, K.; Fujihara, R.; Uemura, N.; Yanase, K.; Kamada, M. Disturbance of Intracerebral Fluid Clearance and Blood–Brain Barrier in Vascular Cognitive Impairment. Int. J. Mol. Sci. 2019, 20, 2600. https://doi.org/10.3390/ijms20102600
Ueno M, Chiba Y, Murakami R, Matsumoto K, Fujihara R, Uemura N, Yanase K, Kamada M. Disturbance of Intracerebral Fluid Clearance and Blood–Brain Barrier in Vascular Cognitive Impairment. International Journal of Molecular Sciences. 2019; 20(10):2600. https://doi.org/10.3390/ijms20102600
Chicago/Turabian StyleUeno, Masaki, Yoichi Chiba, Ryuta Murakami, Koichi Matsumoto, Ryuji Fujihara, Naoya Uemura, Ken Yanase, and Masaki Kamada. 2019. "Disturbance of Intracerebral Fluid Clearance and Blood–Brain Barrier in Vascular Cognitive Impairment" International Journal of Molecular Sciences 20, no. 10: 2600. https://doi.org/10.3390/ijms20102600