Evaluation of Biosynthesis, Accumulation and Antioxidant Activityof Vitamin E in Sweet Corn (Zea mays L.) during Kernel Development

 , and
, and
Abstract
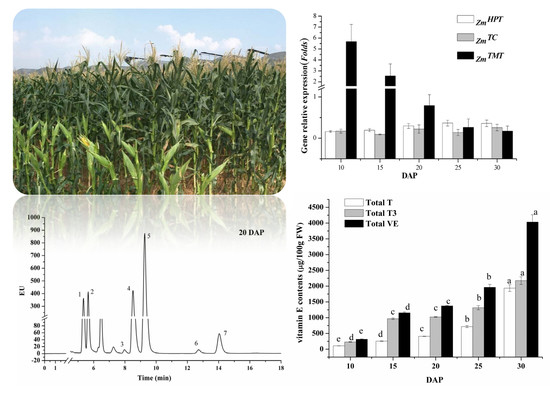
:
1. Introduction
2. Results and Discussion
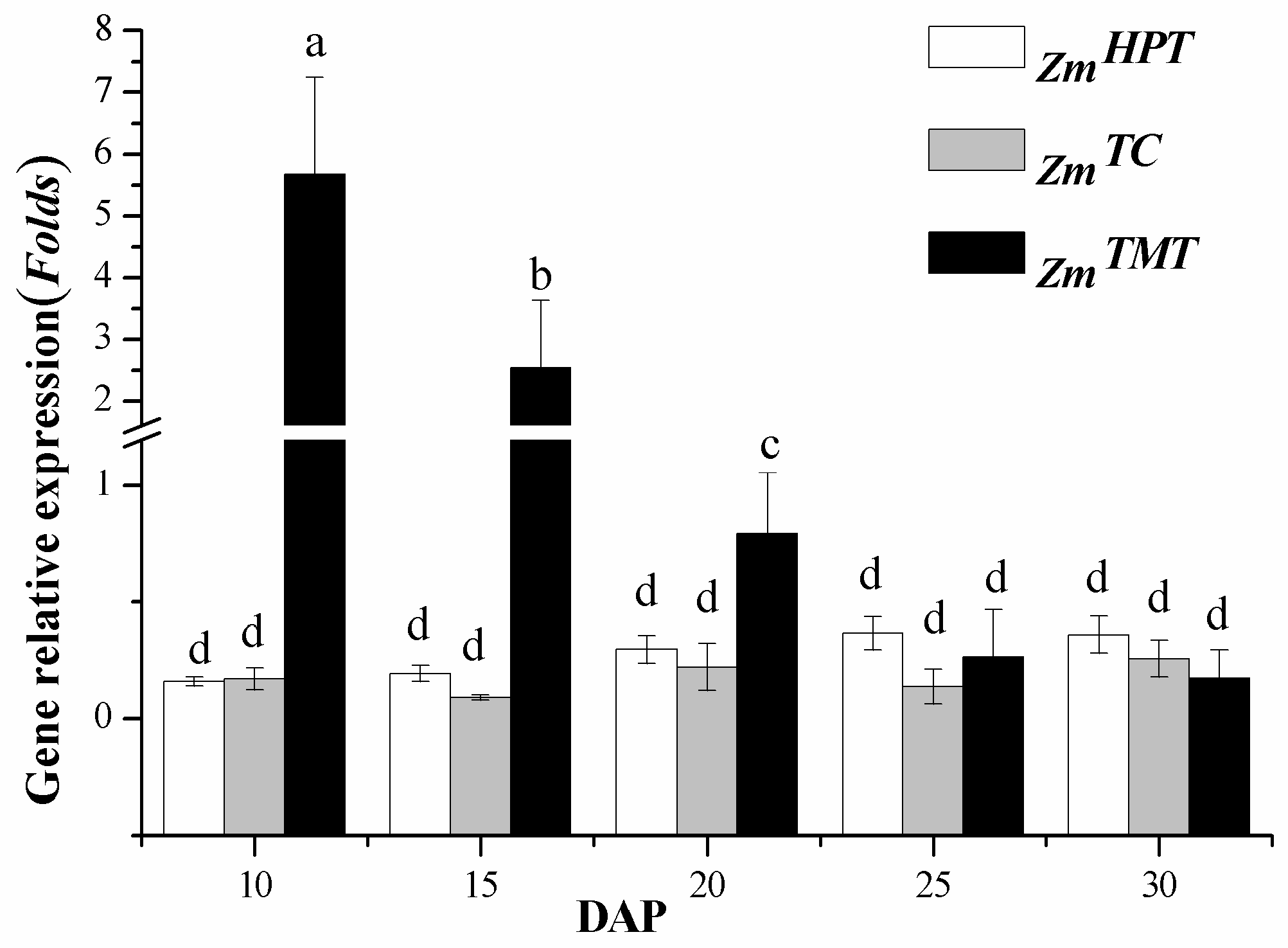
2.1. Expression Analysis of Key Synthetic Genes
2.2. NP-HPLC Chromatogram and Peak Identification of Vitamin E
2.3. Changes of Vitamin E Composition and Contents
2.4. Changes in the Lipophilic Antioxidant Activity of the Sweet Corn Extracts
3. Materials and Methods
3.1. Plant Material Preparation
3.2. RNA Extraction and Quantitative Gene Expression Analysis
3.3. Vitamin E Extraction in Sweet Corn
3.4. Quantification of the Vitamin E Isomers by NP-HPLC
3.5. Quantification of Lipophilic Antioxidant Activity
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ulatowski, L.M.; Manor, D. Vitamin E and neurodegeneration. Neurobiol. Dis. 2015, 84, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Zandi, P.P.; Anthony, J.C.; Khachaturian, A.S.; Stone, S.V.; Gustafson, D.; Tschanz, J.A.T.; Norton, M.C.; Welshbohmer, K.A.; Breitner, J.C.S. Reduced Risk of Alzheimer Disease in Users of Antioxidant Vitamin Supplements: The Cache County Study. Arch. Neurol. 2004, 61, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Falk, J.; Munne-Bosch, S. Tocochromanol functions in plants: Antioxidation and beyond. J. Exp. Bot. 2010, 61, 1549–1566. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C. Chemistry and biology of vitamin E. Mol. Nutr. Food Res. 2005, 49, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Saldeen, T.; Li, D.; Mehta, J.L. Differential effects of α- and γ-tocopherol on low-density lipoprotein oxidation, superoxide activity, platelet aggregation and arterial thrombogenesis. J. Am. Coll. Cardiol. 1999, 34, 1208–1215. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienols: Vitamin E beyond tocopherols. Life Sci. 2006, 78, 2088. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G.; Bucciarelli, T.; Mancini, B.; Ilio, C.D.; Capra, V.; D’Orazio, N. The role of the antioxidant vitamin supplementation in the prevention of cardiovascular diseases. Expert Opin. Investig. Drugs 2007, 16, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Shibata, A.; Sookwong, P.; Kawakami, Y.; Eitsuka, T.; Asai, A.; Oikawa, S.; Nakagawa, K. Antiangiogenic and anticancer potential of unsaturated vitamin E (tocotrienol). J. Nutr. Biochem. 2009, 20, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.-Y.; Ward, L.C.; Fong, C.W.; Yap, W.N.; Brown, L. Anti-inflammatory γ- and δ-tocotrienols improve cardiovascular, liver and metabolic function in diet-induced obese rats. Eur. J. Nutr. 2017, 56, 133–150. [Google Scholar] [CrossRef] [PubMed]
- DellaPenna, D.; Pogson, B.J. Vitamin synthesis in plants: Tocopherols and carotenoids. Ann. Rev. Plant Biol. 2006, 57, 711–738. [Google Scholar] [CrossRef]
- Yang, W.Y.; Cahoon, R.E.; Hunter, S.C.; Zhang, C.Y.; Han, J.X.; Borgschulte, T.; Cahoon, E.B. Vitamin E biosynthesis: Functional characterization of the monocot homogentisate geranylgeranyl transferase. Plant J. 2011, 65, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Mènesaffrané, L.; Dellapenna, D. Biosynthesis, regulation and functions of tocochromanols in plants. Plant Physiol. Biochem. 2010, 48, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Chander, S.; Guo, Y.Q.; Yang, X.H.; Yan, J.B.; Zhang, Y.R.; Song, T.M.; Li, J.S. Genetic dissection of tocopherol content and composition in maize grain using quantitative trait loci analysis and the candidate gene approach. Mol. Breed. 2008, 22, 353–365. [Google Scholar] [CrossRef]
- Panfili, G.; Fratianni, A.; Irano, M. Normal phase high-performance liquid chromatography method for the determination of tocopherols and tocotrienols in cereals. J. Agric. Food Chem. 2003, 51, 3940–3944. [Google Scholar] [CrossRef] [PubMed]
- 1Ibrahim, K.E.; Juvik, J.A. Feasibility for improving phytonutrient content in vegetable crops using conventional breeding strategies: Case study with carotenoids and tocopherols in sweet corn and broccoli. J. Agric. Food Chem. 2009, 57, 4636–4644. [Google Scholar] [CrossRef] [PubMed]
- 1Hunter, S.C.; Cahoon, E.B. Enhancing vitamin E in oilseeds: Unraveling tocopherol and tocotrienol biosynthesis. Lipids 2007, 42, 97–108. [Google Scholar] [CrossRef] [PubMed]
- De Vries, B.D.; Shuler, S.L.; Tracy, W.F. Endosperm Carbohydrates in Pseudostarchy and Extreme-sugary Maize Inbreds during Kernel Development. Crop Sci. 2016, 56, 2448–2456. [Google Scholar] [CrossRef]
- Xu, J.G.; Hu, Q.P.; Wang, X.D.; Luo, J.Y.; Liu, Y.; Tian, C.R. Changes in the main nutrients, phytochemicals, and antioxidant activity in yellow corn grain during maturation. J. Agric. Food Chem. 2010, 58, 5751–5756. [Google Scholar] [CrossRef] [PubMed]
- Sattler, S.E.; Gilliland, L.U.; Magallaneslundback, M.; Pollard, M.; Dellapenna, D. Vitamin E is essential for seed longevity and for preventing lipid peroxidation during germination. Plant Cell 2004, 16, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Wang, Q.; Zhang, J.; Yang, R.; Li, X. Assessment of carotenoid and tocopherol level in sweet corn inbred lines during kernel development stages. Indian J. Genet. Plant Breed. 2015, 75, 196. [Google Scholar] [CrossRef]
- Motto, M.; Balconi, C.; Hartings, H.; Rossi, V. Gene discovery for improvement of kernel quality-related traits in maize. Genetika 2010, 42, 23–56. [Google Scholar] [CrossRef] [Green Version]
- Lipka, A.E.; Gore, M.A.; Magallaneslundback, M.; Mesberg, A.; Lin, H.; Tiede, T.; Chen, C.; Buell, C.R.; Buckler, E.S.; Rocheford, T. Genome-Wide Association Study and Pathway-Level Analysis of Tocochromanol Levels in Maize Grain. G3 Genes Genomes Genet. 2013, 3, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Kurilich, A.C.; Juvik, J.A. Quantification of carotenoid and tocopherol antioxidants in Zea mays. J. Agric. Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Harish, M.C.; Dachinamoorthy, P.; Balamurugan, S.; Bala Murugan, S.; Sathishkumar, R. Overexpression of homogentisate phytyltransferase (HPT) and tocopherol cyclase (TC) enhances α-tocopherol content in transgenic tobacco. Biol. Plant. 2013, 57, 395–400. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, Y.; Zhu, Y.; Zhang, L.; Zhang, W.; Chen, R.; Xu, M.; Fan, Y.; Wang, L. GmTMT2a from soybean elevates the α-tocopherol content in corn and Arabidopsis. Transgenic Res. 2013, 22, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Horvath, G.; Wessjohann, L.; Bigirimana, J.; Monica, H.; Jansen, M.; Guisez, Y.; Caubergs, R.; Horemans, N. Accumulation oftocopherols andtocotrienols during seed development ofgrape (Vitisvinifera L. cv. Albert Lavallée). Plant Physiol. Biochem. 2006, 44, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Theile, K.; Böhm, V. In vitro antioxidant activity of tocopherols and tocotrienols and comparison of vitamin E concentration and lipophilic antioxidant capacity in human plasma. Mol. Nutr. Food Res. 2010, 54, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Adom, K.K.; Liu, R.H. Rapid peroxyl radical scavenging capacity (PSC) assay for assessing both hydrophilic and lipophilic antioxidants. J. Agric. Food Chem. 2005, 53, 6572–6580. [Google Scholar] [CrossRef] [PubMed]
- Galli, V.; Messias, R.d.S.; dos Anjos e Silva, S.D.; Rombaldi, C.V. Selection of reliable reference genes for quantitative real-time polymerase chain reaction studies in maize grains. Plant Cell Rep. 2013, 32, 1869–1877. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Nan, X.; Jian Guang, H.; Yan, S.; Xie, L.; Brennan, C.S.; Huang, W.; Guo, X. The manipulation of gene expression and the biosynthesis of Vitamin C, E and folate in light-and dark-germination of sweet corn seeds. Sci. Rep. 2017, 7, 7484. [Google Scholar] [CrossRef] [PubMed]
- Xiang, N.; Guo, X.; Liu, F.; Li, Q.; Hu, J.; Brennan, C.S. Effect of Light- and Dark-Germination on the Phenolic Biosynthesis, Phytochemical Profiles, and Antioxidant Activities in Sweet Corn (Zea mays L.) Sprouts. Int. J. Mol. Sci. 2017, 18, 1246. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Lai, Q.; Li, T.; Fu, X.; Guo, X.; Liu, R.H. The dynamic changes of ascorbic acid, tocopherols and antioxidant activity during germination of soya bean (Glycine max). Int. J. Food Sci. Technol. 2015, 50, 2367–2374. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 10 DAP | 15 DAP | 20 DAP | 25 DAP | 30 DAP | |
|---|---|---|---|---|---|
| α-T | 45.15 ± 1.46 e | 122.6 ± 2.9 d | 250.7 ± 2.4 c | 375.0 ± 18.9 b | 979.0 ± 55.6 a |
| β-T | 13.82 ± 0.04 e | 17.47 ± 0.12 d | 20.61 ± 0.05 c | 28.98 ± 0.81 b | 46.75 ± 2.08 a |
| γ-T | 29.29 ± 0.84 d | 91.79 ± 2.08 c | 115.6 ± 1.4 c | 272.3 ±12.4 b | 832.8 ±47.4 a |
| δ-T | 18.46 ± 0.09 d | 24.40 ± 0.32 c | 22.91 ± 0.10 c | 40.26 ± 1.10 b | 78.82 ± 3.62 a |
| α-T3 | 94.07 ± 3.22 e | 168.4 ± 4.0 d | 205.7 ± 1.3 c | 253.2 ± 12.1 b | 383.6 ± 21.8 a |
| γ-T3 | 115.2 ± 5.2 d | 728.7 ± 18.5 c | 758.8 ± 8.2 c | 992.1 ± 48.7 b | 1711 ± 100 a |
| δ-T3 | 21.14 ± 0.18 d | 70.74 ± 1.27 b | 58.80 ± 0.60 c | 69.17 ± 2.39 b | 81.17 ± 3.94 a |
| T/T3 | 0.46 | 0.26 | 0.40 | 0.55 | 0.89 |
| α/γ-T | 1.54 | 1.33 | 2.17 | 1.38 | 1.17 |
| α/γ-T3 | 0.81 | 0.23 | 0.27 | 0.26 | 0.22 |
| DAP | Concentration Range (mg/mL) | EC50 (mg/mL) | Dose Curve R2 | Lipo-PSC Value (μmol α-Tocopherol Equiv./100 g FW) |
|---|---|---|---|---|
| 10 | 80–400 | NC * | NC | NC |
| 15 | 50–400 | NC | NC | NC |
| 20 | 26.7–300 | 99.03 ± 20.48 | 0.964 | 39.08 ± 7.72 c |
| 25 | 20–200 | 63.08 ± 10.28 | 0.987 | 60.67 ± 9.10 b |
| 30 | 10–80 | 38.16 ± 8.01 | 0.981 | 101.8 ± 22.3 a |
| Gene Name | Gene ID | Prime Direction | Primer Sequence (5′-3′) |
|---|---|---|---|
| ZmHPT | 732789 | Forward | TCCATTGGCATCTGGGGAAT |
| Reward | TGCAGTCCCAAGAACAAAGC | ||
| ZmTC | 541877 | Forward | TGGGATGGAGAACGGTTTGA |
| Reward | CAGAAGCTCCTGGGAAGACA | ||
| ZmTMT | 732837 | Forward | CCATCATCACCTGTCGCAAG |
| Reward | AGATGAGTAGACGGCGATGG | ||
| ZmActin | 100282267 | Forward | TGTGGCTTTGGGATCGTAGTC |
| Reward | GAGCCACCGATCCAGACACT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Yu, Y.; Mao, J.; Liu, H.; Hu, J.G.; Li, T.; Guo, X.; Liu, R.H. Evaluation of Biosynthesis, Accumulation and Antioxidant Activityof Vitamin E in Sweet Corn (Zea mays L.) during Kernel Development. Int. J. Mol. Sci. 2017, 18, 2780. https://doi.org/10.3390/ijms18122780
Xie L, Yu Y, Mao J, Liu H, Hu JG, Li T, Guo X, Liu RH. Evaluation of Biosynthesis, Accumulation and Antioxidant Activityof Vitamin E in Sweet Corn (Zea mays L.) during Kernel Development. International Journal of Molecular Sciences. 2017; 18(12):2780. https://doi.org/10.3390/ijms18122780
Chicago/Turabian StyleXie, Lihua, Yongtao Yu, Jihua Mao, Haiying Liu, Jian Guang Hu, Tong Li, Xinbo Guo, and Rui Hai Liu. 2017. "Evaluation of Biosynthesis, Accumulation and Antioxidant Activityof Vitamin E in Sweet Corn (Zea mays L.) during Kernel Development" International Journal of Molecular Sciences 18, no. 12: 2780. https://doi.org/10.3390/ijms18122780