

Stilbenes, a Versatile Class of Natural Metabolites for Inflammation—An Overview

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Diversity of Stilbenes
3. Inflammatory Response
4. Mechanisms
4.1. Nuclear Factor Kappa-B (NF-κB) Pathway
4.2. MAPK Pathway
4.3. JAK-STAT Pathway
5. Effect of Stilbenes on Various Types of Inflammation
5.1. Inflammation in Macrophages
5.2. Inflammation in the Liver
5.3. Inflammation in the Cardiac Tissue
5.4. Inflammation in the Connective Tissue
5.5. Inflammation in the Nephrons
5.6. Inflammation in the Intestine
5.7. Inflammation in the Lungs
5.8. Inflammation in the Nervous Tissue
6. Effect of Stilbenes on AChE and BuChE
7. Bioavailability
8. Conclusions
9. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ansari, N.; Khodagholi, F. Natural Products as Promising Drug Candidates for the Treatment of Alzheimer’s Disease: Molecular Mechanism Aspect. Curr. Neuropharmacol. 2013, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, M.; Ninomiya, M.; Oshima, N.; Itoh, T.; Tanaka, K.; Nishina, A.; Koketsu, M. Comparative analysis of stilbene and benzofuran neolignan derivatives as acetylcholinesterase inhibitors with neuroprotective and anti-inflammatory activities. Bioorg. Med. Chem. Lett. 2019, 29, 17. [Google Scholar] [CrossRef] [PubMed]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N.K. Resveratrol: French Paradox Revisited. Front. Pharmacol. 2012, 3, 141. [Google Scholar] [CrossRef] [PubMed]
- Aja-Perez, I.; Krisa, S.; Hornedo-Ortega, R.; Begoña Ruiz-Larrea, M.; Ruiz-Sanz, J.I.; Richard, T.; Courtois, A. Stilbenes at Low Micromolar Concentrations Mitigate the NO, TNF-α, IL-1β and ROS Production in LPS-Stimulated Murine Macrophages. J. Biol. Act. Prod. Nat. 2021, 11, 3. [Google Scholar] [CrossRef]
- Lee, I.T.; Lin, H.-C.; Huang, T.-H.; Tseng, C.-N.; Cheng, H.-T.; Huang, W.-C.; Cheng, C.-Y. Anti-Inflammatory Effect of Resveratrol Derivatives via the Downregulation of Oxidative-Stress-Dependent and c-Src Transactivation EGFR Pathways on Rat Mesangial Cells. Antioxidants 2022, 11, 5. [Google Scholar] [CrossRef]
- Zhan, J.; Hu, T.; Shen, J.; Yang, G.; Ho, C.T.; Li, S. Pterostilbene is more efficacious than hydroxystilbenes in protecting liver fibrogenesis in a carbon tetracholride-induced rat model. J. Funct. Foods 2021, 84, 104604. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, Z.; Yao, C.; Bai, J.; Yang, H.; Ma, P.; Fan, Y.; Li, S.; Yuan, J.; Lin, M.; et al. Amurensin H, a derivative from resveratrol, ameliorates lipopolysaccharide/cigarette smoke-induced airway inflammation by blocking the Syk/NF-κB pathway. Front. Pharmacol. 2019, 10, 1157. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, M.; Jiang, L.; Xie, Q.; Yuan, H.; Yang, Y.; Zafar, S.; Liu, Y.; Jian, Y.; Li, B.; et al. The medicinal uses of the genus Bletilla in traditional Chinese medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2021, 280, 114263. [Google Scholar] [CrossRef]
- Nassra, M.; Krisa, S.; Papastamoulis, Y.; Deccaux, G.K.; Bisson, J.; André, C.; Konsman, J.; Schmitter, J.; Mérillon, J.; Waffo-Téguo, P. Inhibitory activity of plant stilbenoids against nitric oxide production by lipopolysaccharide-activated microglia. Planta Med. 2013, 79, 11. [Google Scholar] [CrossRef]
- Koronowski, K.B.; Dave, K.R.; Saul, I.; Camarena, V.; Thompson, J.W.; Neumann, J.T.; Young, J.I.; Perez-Pinzon, M.A. Resveratrol Preconditioning Induces a Novel Extended Window of Ischemic Tolerance in the Mouse Brain. Stroke 2015, 46, 2293–2298. [Google Scholar] [CrossRef]
- Colombo, F.; Di Lorenzo, C.; Regazzoni, L.; Fumagalli, M.; Sangiovanni, E.; de Sousa, L.P.; Bavaresco, L.; Tomasi, D.; Bosso, A.; Aldini, G.; et al. Phenolic profiles and anti-inflammatory activities of sixteen table grape (Vitis vinifera L.) varieties. Food Funct. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Baderschneider, B.; Winterhalter, P. Isolation and Characterization of Novel Stilbene Derivatives from Riesling Wine. J. Agric. Food Chem. 2000, 48, 2681–2686. [Google Scholar] [CrossRef] [PubMed]
- Hornedo-Ortega, R.; Jourdes, M.; Da Costa, G.; Courtois, A.; Gabaston, J.; Teissedre, P.-L.; Richard, T.; Krisa, S. Oxyresveratrol and Gnetol Glucuronide Metabolites: Chemical Production, Structural Identification, Metabolism by Human and Rat Liver Fractions, and in Vitro Anti-inflammatory Properties. J. Agric. Food Chem. 2022, 70, 41. [Google Scholar] [CrossRef]
- Du, Q.H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 11. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zeng, Z.; Li, T.; Xu, S.; Wang, X.; Chen, Z.; Lin, C. Polydatin inhibits mitochondrial dysfunction in the renal tubular epithelial cells of a rat model of sepsis-induced acute kidney injury. Anesth. Analg. 2015, 121, 5. [Google Scholar] [CrossRef]
- Romero-Pérez, A.I.; Lamuela-Raventós, R.M.; Andrés-Lacueva, C.; De La Carmen Torre-Boronat, M. Method for the quantitative extraction of resveratrol and piceid isomers in grape berry skins. Effect of powdery mildew on the stilbene content. J. Agric. Food Chem. 2001, 49, 1. [Google Scholar] [CrossRef]
- Waffo-Teguo, P.; Lee, D.; Cuendet, M.; Mërillon, J.M.; Pezzuto, J.M.; Kinghorn, A.D. Two new stilbene dimer glucosides from grape (Vitis vinifera) cell cultures. J. Nat. Prod. 2001, 64, 136–138. [Google Scholar] [CrossRef]
- Zghonda, N.; Yoshida, S.; Araki, M.; Kusunoki, M.; Mliki, A.; Ghorbel, A.; Miyazaki, H. Greater effectiveness of ε-viniferin in red wine than its monomer resveratrol for inhibiting vascular smooth muscle cell proliferation and migration. Biosci. Biotechnol. Biochem. 2011, 75, 7. [Google Scholar] [CrossRef]
- Fuloria, S.; Sekar, M.; Khattulanuar, F.S.; Gan, S.H.; Rani, N.N.I.M.; Ravi, S.; Subramaniyan, V.; Jeyabalan, S.; Begum, M.Y.; Chidambaram, K.; et al. Chemistry, Biosynthesis and Pharmacology of Viniferin: Potential Resveratrol-Derived Molecules for New Drug Discovery, Development and Therapy. Molecules 2022, 27, 5072. [Google Scholar] [CrossRef]
- Gabaston, J.; Fonayet, J.V.; Franc, C.; Waffo-Teguo, P.; de Revel, G.; Hilbert, G.; Gomès, E.; Richard, T.; Mérillon, J.-M. Characterization of Stilbene Composition in Grape Berries from Wild Vitis Species in Year-To-Year Harvest. J. Agric. Food Chem. 2020, 68, 47. [Google Scholar] [CrossRef]
- Privat, C.; Telo, J.P.; Bernardes-Genisson, V.; Vieira, A.; Souchard, J.P.; Nepveu, F. Antioxidant properties of trans-ε-Viniferin as compared to stilbene derivatives in aqueous and nonaqueous media. J. Agric. Food Chem. 2002, 50, 5. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.-R.; Yao, S.; Liu, Q.; Peng, Z.-L.; Deng, Q.-Q.; Liu, B.; Ma, Z.-H.; Wang, L.; Zhou, H.; Ye, Y.; et al. Dihydro-stilbene gigantol relieves CCl4-induced hepatic oxidative stress and inflammation in mice via inhibiting C5b-9 formation in the liver. Acta Pharmacol. Sin. 2020, 41, 11. [Google Scholar] [CrossRef] [PubMed]
- Nchiozem-Ngnitedem, V.A.; Omosa, L.K.; Bedane, K.G.; Derese, S.; Brieger, L.; Strohmann, C.; Spiteller, M. Anti-inflammatory steroidal sapogenins and a conjugated chalcone-stilbene from Dracaena usambarensis Engl. Fitoterapia 2020, 146, 104717. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk-Czepas, J.; Czepas, J. Rhaponticin as an anti-inflammatory component of rhubarb: A minireview of the current state of the art and prospects for future research. Phytochem. Rev. 2003, 18, 5. [Google Scholar] [CrossRef]
- Aaviksaar, A.; Haga, M.; Kuzina, K.; Püssa, T.; Raal, A.; Tsoupras, G. Hydroxystilbenes in the roots of Rheum rhaponticum. In Proceedings of the Estonian Academy of Sciences, Chemistry; Estonian Academy Publishers: Tallinn, Estonia, 2019; Volume 52. [Google Scholar] [CrossRef]
- Gatto, P.; Vrhovsek, U.; Muth, J.; Segala, C.; Romualdi, C.; Fontana, P.; Pruefer, D.; Stefanini, M.; Moser, C.; Mattivi, F.; et al. Ripening and genotype control stilbene accumulation in healthy grapes. J. Agric. Food Chem. 2008, 56, 24. [Google Scholar] [CrossRef]
- Dabauza, M.; Velasco, L.; Pazos-Navarro, M.; Pérez-Benito, E.; Hellín, P.; Flores, P.; Gómez-Garay, A.; Martínez, M.C.; Lacasa, A. Enhanced resistance to Botrytis cinerea in genetically-modified Vitis vinifera L. plants over-expressing the grapevine stilbene synthase gene. Plant. Cell Tissue Organ. Cult. 2015, 120, 1. [Google Scholar] [CrossRef]
- Limmongkon, A.; Nopprang, P.; Chaikeandee, P.; Somboon, T.; Wongshaya, P.; Pilaisangsuree, V. LC-MS/MS profiles and interrelationships between the anti-inflammatory activity, total phenolic content and antioxidant potential of Kalasin 2 cultivar peanut sprout crude extract. Food Chem. 2018, 239, 332–338. [Google Scholar] [CrossRef]
- Ma, P.; Yue, L.; Yang, H.; Fan, Y.; Bai, J.; Li, S.; Yuan, J.; Zhang, Z.; Yao, C.; Lin, M.; et al. Chondroprotective and anti-inflammatory effects of amurensin H by regulating TLR4/Syk/NF-κB signals. J. Cell Mol. Med. 2020, 24, 2. [Google Scholar] [CrossRef]
- Biais, B.; Krisa, S.; Cluzet, S.; Da Costa, G.; Waffo-Teguo, P.; Mérillon, J.; Richard, T. Antioxidant and Cytoprotective Activities of Grapevine Stilbenes. J. Agric. Food Chem. 2017, 65, 24. [Google Scholar] [CrossRef]
- Chiou, W.F.; Shen, C.C.; Chen, C.C.; Lin, C.H.; Huang, Y.L. Oligostilbenes from the roots of Vitis thunbergii. Planta Med. 2009, 75, 8. [Google Scholar] [CrossRef]
- Liu, S.; Li, Y.; Yi, F.; Liu, Q.; Chen, N.; He, X.; He, C.; Xiao, P. Resveratrol oligomers from Paeonia suffruticosa protect mice against cognitive dysfunction by regulating cholinergic, antioxidant and anti-inflammatory pathways. J. Ethnopharmacol. 2020, 260, 112983. [Google Scholar] [CrossRef]
- Hankittichai, P.; Lou, H.J.; Wikan, N.; Smith, D.R.; Potikanond, S.; Nimlamool, W. Oxyresveratrol inhibits IL-1β-induced inflammation via suppressing AKT and ERK1/2 activation in human microglia, HMC3. Int. J. Mol. Sci. 2020, 21, 6054. [Google Scholar] [CrossRef] [PubMed]
- Remsberg, C.M.; Martinez, S.E.; Akinwumi, B.C.; Anderson, H.D.; Takemoto, J.K.; Sayre, C.L.; Davies, N.M. Preclinical Pharmacokinetics and Pharmacodynamics and Content Analysis of Gnetol in Foodstuffs. Phytother. Res. 2015, 29, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Z.; Lee, S.O.; Jang, K.U.; Chung, S.H.; Park, S.H.; Kang, H.C.; Yang, E.Y.; Cho, H.J.; Lee, K.R. Antidiabetic stilbene and anthraquinone derivatives from Rheum. undulatum. Arch. Pharm. Res. 2005, 28, 9. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk-Czepas, J.; Liudvytska, O. Rheum rhaponticum and Rheum. rhabarbarum: A review of phytochemistry, biological activities and therapeutic potential. Phytochem. Rev. 2021, 20, 3. [Google Scholar] [CrossRef]
- Mittas, D.; Mawunu, M.; Magliocca, G.; Lautenschläger, T.; Schwaiger, S.; Stuppner, H.; Marzocco, S. Bioassay-Guided Isolation of Anti-Inflammatory Constituents of the Subaerial Parts of Cyperus articulatus (Cyperaceae). Molecules 2022, 27, 5937. [Google Scholar] [CrossRef]
- Limmongkon, A.; Pankam, J.; Somboon, T.; Wongshaya, P.; Nopprang, P. Evaluation of the DNA damage protective activity of the germinated peanut (Arachis hypogaea) in relation to antioxidant and anti-inflammatory activity. LWT 2019, 101, 259–268. [Google Scholar] [CrossRef]
- Wu, J.; Li, B.; Xiao, W.; Hu, J.; Xie, J.; Yuan, J.; Wang, L. Longistylin A, a natural stilbene isolated from the leaves of Cajanus cajan, exhibits significant anti-MRSA activity. Int. J. Antimicrob. Agents 2020, 55, 1. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.X.; Xia, T.Q.; He, Q.; Tang, W.; Huang, X.; Song, Q.; Li, Y.; Ye, W.; Wang, Y.; Wu, Z. Stilbenes from the leaves of Cajanus cajan and their in vitro anti-inflammatory activities. Fitoterapia 2022, 160, 105229. [Google Scholar] [CrossRef]
- Leláková, V.; Béraud-Dufour, S.; Hošek, J.; Šmejkal, K.; Prachyawarakorn, V.; Pailee, P.; Widmann, C.; Václavík, J.; Coppola, T.; Mazella, J.; et al. Therapeutic potential of prenylated stilbenoid macasiamenene F through its anti-inflammatory and cytoprotective effects on LPS-challenged monocytes and microglia. J. Ethnopharmacol. 2020, 263, 113147. [Google Scholar] [CrossRef]
- Bakrim, S.; Machate, H.; Benali, T.; Sahib, N.; Jaouadi, I.; Omari, N.E.; Aboulaghras, S.; Bangar, S.P.; Lorenzo, J.M.; Zengin, G.; et al. Natural Sources and Pharmacological Properties of Pinosylvin. Plants 2022, 11, 1541. [Google Scholar] [CrossRef]
- Liu, L.; Zou, M.; Zeng, K.; Ye, X.; Wang, R.; Wang, W.; Zhang, X. Chemical Constituents and their Antioxidant, Anti-Inflammatory and Anti-Acetylcholinesterase Activities from Pholidota cantonensis. Plant. Foods Human. Nutr. 2021, 76, 105–110. [Google Scholar] [CrossRef]
- Gomes, M.J.C.; Kolba, N.; Agarwal, N.; Kim, D.; Eshel, A.; Koren, O.; Tako, E. Modifications in the intestinal functionality, morphology and microbiome following intra-amniotic administration (Gallus gallus) of grape (Vitis vinifera) stilbenes (resveratrol and pterostilbene). Nutrients 2021, 13, 3247. [Google Scholar] [CrossRef] [PubMed]
- Gabaston, J.; Richard, T.; Biais, B.; Waffo-Teguo, P.; Pedrot, E.; Jourdes, M.; Corio-Costet, M.; Mérillon, J. Stilbenes from common spruce (Picea abies) bark as natural antifungal agent against downy mildew (Plasmopara viticola). Ind. Crops Prod. 2017, 103, 267–273. [Google Scholar] [CrossRef]
- Wang, S.Y.; Feng, R.; Bowman, L.; Lu, Y.; Ballington, J.R.; Ding, M. Antioxidant activity of Vaccinium stamineum: Exhibition of anticancer capability in human lung and leukemia cells. Planta Med. 2007, 73, 451–460. [Google Scholar] [CrossRef]
- Rimando, A.M.; Kalt, W.; Magee, J.B.; Dewey, J.; Ballington, J.R. Resveratrol, pterostilbene, and piceatannol in Vaccinium berries. J. Agric. Food Chem. 2004, 52, 15. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New Adventures of an Old Flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Moynagh, P.N. The NF-κB pathway. J. Cell Sci. 2005, 118, 4589–4592. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Natoli, G.; Ghosh, G. Transcriptional regulation via the NF-κB signaling module. Oncogene 2006, 25, 6706–6716. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF- B Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.-J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef]
- Boengler, K.; Hilfikerkleiner, D.; Drexler, H.; Heusch, G.; Schulz, R. The myocardial JAK/STAT pathway: From protection to failure. Pharmacol. Ther. 2008, 120, 172–185. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug. Target. Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Chung, K.-O.; Kim, B.; Lee, M.; Kim, Y.; Chung, H.; Park, J.; Moon, J. In-vitro and in-vivo anti-inflammatory effect of oxyresveratrol from Morus alba L. J. Pharm. Pharmacol. 2010, 55, 1695–1700. [Google Scholar] [CrossRef]
- Lee, H.; Kim, D.; Hong, J.; Lee, J.-Y.; Kim, E. Oxyresveratrol suppresses lipopolysaccharide-induced inflammatory responses in murine macrophages. Hum. Exp. Toxicol. 2015, 34, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.F.; McCall, C.E. Deacetylation by SIRT1 Reprograms Inflammation and Cancer. Genes. Cancer 2013, 4, 135–147. [Google Scholar] [CrossRef]
- Ali, M.; Gupta, M.; Wani, A.; Sharma, A.; Abdullaha, M.; Kour, D.; Choudhary, S.; Bharate, S.B.; Singh, G.; Kumar, A. IIIM-941, a Stilbene Derivative Inhibits NLRP3 Inflammasome Activation by Inducing Autophagy. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 7381. [Google Scholar] [CrossRef] [PubMed]
- Damsker, J.M.; Hansen, A.M.; Caspi, R.R. Th1 and Th17 cells: Adversaries and collaborators. Ann. N. Y. Acad. Sci. 2010, 1183, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.G.; Gray, A.I.; Anthony, N.G.; Mackay, S.P.; Pyne, S.; Pyne, N.J. Resveratrol and its oligomers: Modulation of sphingolipid metabolism and signaling in disease. Arch. Toxicol. 2014, 88, 12. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Wei, H.; Jia, T.; Tian, H. An improved highly sensitive method to determine low oxyresveratrol concentrations in rat plasma and its pharmacokinetic application. Biomed. Chromatogr. 2014, 28, 5. [Google Scholar] [CrossRef]
- Fonseca-Kelly, Z.; Nassrallah, M.; Uribe, J.; Khan, R.S.; Dine, K.; Dutt, M.; Shindler, K.S. Resveratrol Neuroprotection in a Chronic Mouse Model of Multiple Sclerosis. Front. Neurol. 2012, 3, 84. [Google Scholar] [CrossRef]
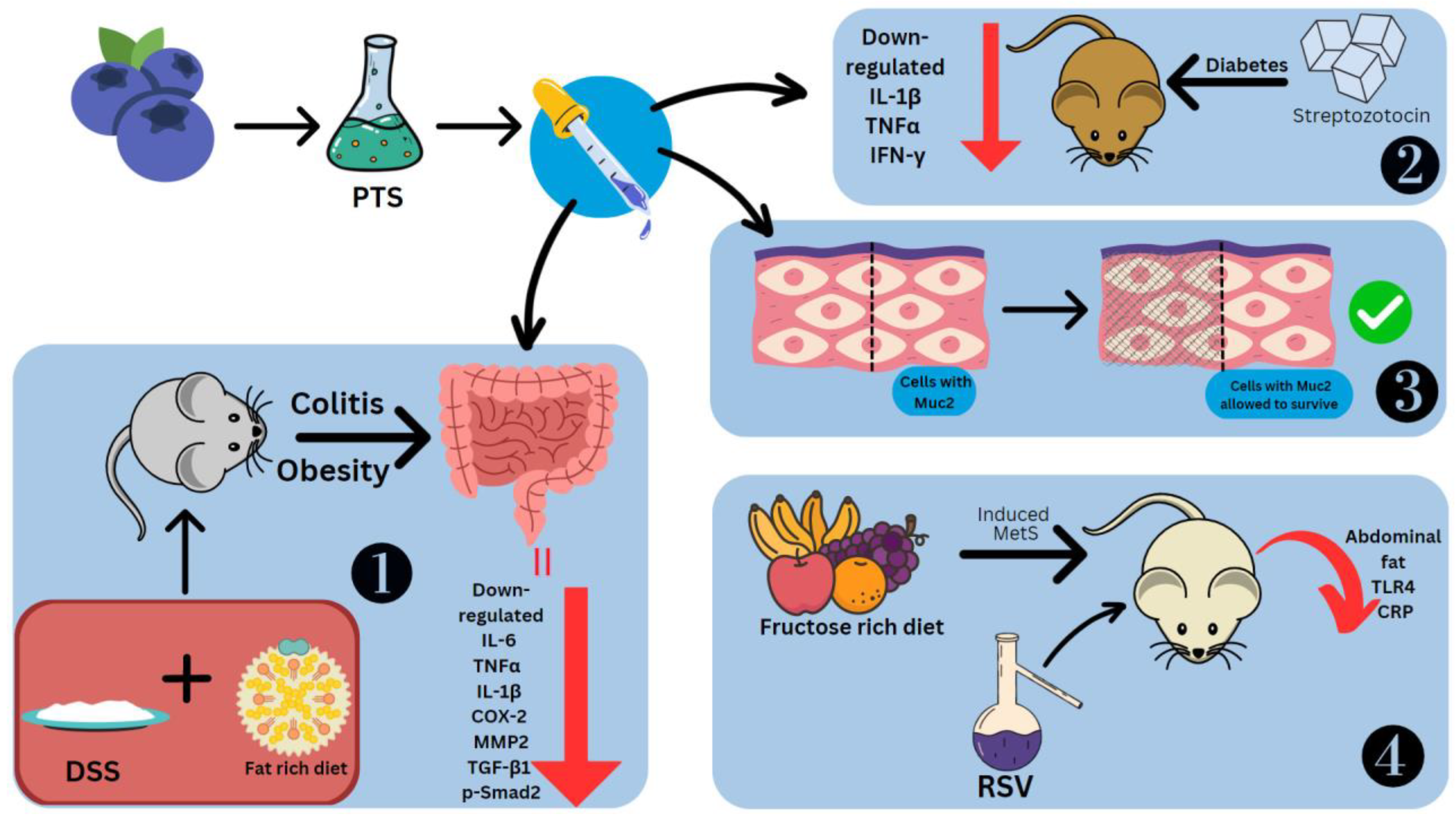
- Singh, U.P.; Singh, N.P.; Singh, B.; Hofseth, L.J.; Price, R.L.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol (trans-3,5,4′-trihydroxystilbene) induces silent mating type information regulation-1 and down-regulates nuclear transcription factor-κB activation to abrogate dextran sulfate sodium-induced colitis. J. Pharmacol. Exp. Ther. 2010, 332, 3. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Yang, Y.; Xia, F.; Huang, A.; Gao, X.; Fang, D. Resveratrol Inhibits CD4+ T Cell Activation by Enhancing the Expression and Activity of Sirt1. PLoS ONE 2013, 8, e75139. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.; Cohen, H. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Vaquero, A.; Scher, M.; Lee, D.; Erdjument-Bromage, H.; Tempst, P.; Reinberg, D. Human SirT1 Interacts with Histone H1 and Promotes Formation of Facultative Heterochromatin. Mol. Cell 2004, 16, 93–105. [Google Scholar] [CrossRef]
- Tsai, C.-F.; Wang, K.-T.; Chen, L.-G.; Lee, C.-J.; Tseng, S.-H.; Wang, C.-C. Anti-Inflammatory Effects of Vitis thunbergii var. taiwaniana on Knee Damage Associated with Arthritis. J. Med. Food 2014, 17, 479–486. [Google Scholar] [CrossRef]
- Song, J.; Jun, M.; Ahn, M.R.; Kim, O.Y. Involvement of miR-let7A in inflammatory response and cell survival/apoptosis regulated by resveratrol in THP-1 macrophage. Nutr. Res. Pract. 2016, 10, 377–384. [Google Scholar] [CrossRef]
- Park, H.S.; Vick, E.J.; Gao, Y.; He, C.; Almosnid, N.M.; Farone, M.; Farone, A.L. cis- and trans-gnetin H from Paeonia suffruticosa suppress inhibitor kappa B kinase phosphorylation in LPS-stimulated human THP-1 cells. J. Ethnopharmacol. 2016, 189, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Tasinov, O.; Dincheva, I.; Badjakov, I.; Kiselova-Kaneva, Y.; Galunska, B.; Nogueiras, R.; Ivanova, D. Phytochemical composition, anti-inflammatory and er stress-reducing potential of Sambucus ebulus L. Fruit extract. Plants 2021, 10, 2446. [Google Scholar] [CrossRef] [PubMed]
- Schueller, K.; Pignitter, M.; Somoza, V. Sulfated and Glucuronated trans-Resveratrol Metabolites Regulate Chemokines and Sirtuin-1 Expression in U-937 Macrophages. J. Agric. Food Chem. 2015, 63, 6535–6545. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, X.; Ruan, C.C.; Cheang, W.S. Two methoxy derivatives of resveratrol, 3,3′,4,5′-tetramethoxy-trans-stilbene and 3,4′,5-trimethoxy-trans-stilbene, suppress lipopolysaccharide-induced inflammation through inactivation of MAPK and NF-κB pathways in RAW 264.7 cells. Chin. Med. 2021, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kloypan, C.; Jeenapongsa, R.; Sri-in, P.; Chanta, S.; Dokpuang, D.; Tip-pyang, S.; Surapinit, N. Stilbenoids from Gnetum macrostachyum attenuate human platelet aggregation and adhesion. Phytother. Res. 2012, 26, 1564–1568. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Chung, H.T.; Pae, H.O. Differential effects of resveratrol and its natural analogs, piceatannol and 3,5,4′-trans-trimethoxystilbene, on anti-inflammatory heme oxigenase-1 expression in RAW264.7 macrophages. BioFactors 2014, 40, 138–145. [Google Scholar] [CrossRef]
- Kim, D.-H.; Lee, Y.-G.; Park, H.-J.; Lee, J.-A.; Kim, H.J.; Hwang, J.-K.; Choi, J.-M. Piceatannol inhibits effector T cell functions by suppressing TcR signaling. Int. Immunopharmacol. 2015, 25, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Khan, H.; Hoi, M.P.M.; Cheang, W.S. Piceatannol Protects Brain Endothelial Cell Line (bEnd.3) against Lipopolysaccharide-Induced Inflammation and Oxidative Stress. Molecules 2022, 27, 1206. [Google Scholar] [CrossRef]
- Hung, N.Q.; Anh, N.T.H.; Khang, N.S.; Huong, N.T.T.; Luyenb, N.T.; Hau, D.V.; Dat, N.T. Undescribed chalcone and stilbene constituents from Lysimachia baviensis and their anti-inflammatory effect. Nat. Prod. Res. 2021, 37, 1138–1145. [Google Scholar] [CrossRef]
- Hsu, C.L.; Lin, Y.J.; Ho, C.T.; Yen, G.C. The inhibitory effect of pterostilbene on inflammatory responses during the interaction of 3T3-L1 adipocytes and RAW 264.7 macrophages. J. Agric. Food Chem. 2013, 61, 602–610. [Google Scholar] [CrossRef]
- Qi, S.; Xin, Y.; Guo, Y.; Diao, Y.; Kou, X.; Luo, L.; Yin, Z. Ampelopsin reduces endotoxic inflammation via repressing ROS-mediated activation of PI3K/Akt/NF-κB signaling pathways. Int. Immunopharmacol. 2012, 12, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yuan, L.; Liu, J.; Muhammad, I.; Cao, C.; Shi, C.; Zhang, Y.; Li, R.; Li, C.; Liu, F. Dihydromyricetin attenuates Escherichia coli lipopolysaccharide-induced ileum injury in chickens by inhibiting NLRP3 inflammasome and TLR4/NF-κB signalling pathway. Vet. Res. 2020, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Jiang, F.; Huang, B.; Zheng, W.; Jiang, Y.; Cai, G.; Liu, D.; Hu, C.Y.; Wang, C. Dihydromyricetin resists inflammation-induced muscle atrophy via ryanodine receptor-CaMKK-AMPK signal pathway. J. Cell Mol. Med. 2021, 25, 21. [Google Scholar] [CrossRef]
- Ko, Y.-J.; Kim, H.; Kim, E.; Katakura, Y.; Lee, W.; Kim, G.; Ryu, C. Piceatannol inhibits mast cell-mediated allergic inflammation. Int. J. Mol. Med. 2013, 31, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H.; Park, E.-K.; Bae, E.-A.; Han, M.J. Metabolism of Rhaponticin and Chrysophanol 8-o-.BETA.-D-Glucopyranoside from the Rhizome of Rheum undulatum by Human Intestinal Bacteria and Their Anti-allergic Actions. Biol. Pharm. Bull. 2000, 23, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Labib, R.M.; Malak, L.G.; Youssef, F.S.; Ross, S.A. A new stilbene from Agonis flexuosa leaves and verification of its histamine release inhibitory activity using in silico and in vitro studies. S. Afr. J. Bot. 2020, 135, 384–390. [Google Scholar] [CrossRef]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef]
- Morgan, B.P. The membrane attack complex as an inflammatory trigger. Immunobiology 2016, 221, 6. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Schemitt, E.G.; Da Silva, J.; Marroni, N.P.; Lima, A.; Ferreira, R.B. Combination of trans-resveratrol and ε-viniferin induces a hepatoprotective effect in rats with severe acute liver failure via reduction of oxidative stress and MMP-9 expression. Nutrients 2021, 13, 3677. [Google Scholar] [CrossRef]
- Pham, C.G.; Bubici, C.; Zazzeroni, F.; Papa, S.; Jones, J.; Alvarez, K.; Jayawardena, S.; De Smaele, E.; Cong, R.; Beaumont, C.; et al. Ferritin heavy chain upregulation by NF-κB inhibits TNFα-induced apoptosis by suppressing reactive oxygen species. Cell 2004, 119, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Yu, L.-R.; Abdelmegeed, M.A.; Gao, Y.; Banerjee, A.; Song, B.-J. Critical role of c-jun N-terminal protein kinase in promoting mitochondrial dysfunction and acute liver injury. Redox Biol. 2015, 6, 552–564. [Google Scholar] [CrossRef] [PubMed]
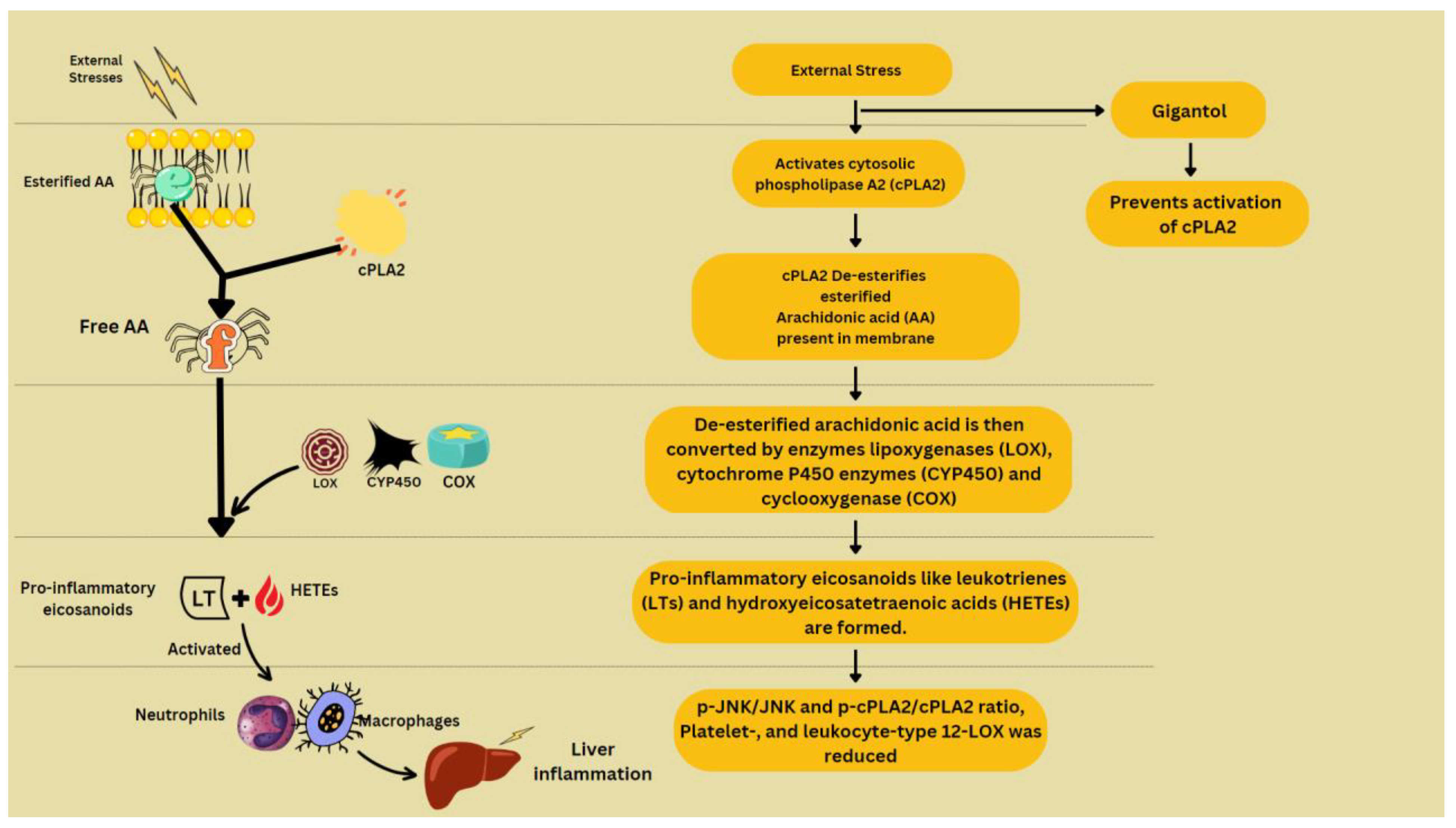
- Xue, Y.; Deng, Q.; Zhang, Q.; Ma, Z.; Chen, B.; Yu, X.; Peng, H.; Yao, S.; Liu, J.; Ye, Y.; et al. Gigantol ameliorates CCl4-induced liver injury via preventing activation of JNK/cPLA2/12-LOX inflammatory pathway. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Lin, D.; Sun, Z.; Jin, Z.; Lei, L.; Liu, Y.; Hu, B.; Wang, B.; Shen, Y.; Wang, Y. Matrix Remodeling Associated 7 Deficiency Alleviates Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Front. Immunol. 2018, 9, 773. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, P.; Courtois, A.; Atgié, C.; Richard, T.; Krisa, S. Correction to: In the shadow of resveratrol: Biological activities of epsilon-viniferin. J. Physiol. Biochem. 2022, 78, 465–484. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Zhang, T.; Gu, W.; Yang, X.; Zhao, R.; Yu, J. 2,3,5,4′-tetrahydroxy-stilbene-2-O-β-D-glucoside attenuates methionine and choline-deficient diet-induced non-alcoholic fatty liver disease. Exp. Ther. Med. 2018, 16, 2. [Google Scholar] [CrossRef]
- Feng, Y.; Cui, R.; Li, Z.; Zhang, X.; Jia, Y.; Zhang, X.; Shi, J.; Qu, K.; Liu, C.; Zhang, J. Methane Alleviates Acetaminophen-Induced Liver Injury by Inhibiting Inflammation, Oxidative Stress, Endoplasmic Reticulum Stress, and Apoptosis through the Nrf2/HO-1/NQO1 Signaling Pathway. Oxid. Med. Cell Longev. 2019, 2019, 7067619. [Google Scholar] [CrossRef]
- Jia, Y.N.; Peng, Y.L.; Zhao, Y.P.; Cheng, X.F.; Zhou, Y.; Chai, C.L.; Zeng, L.S.; Pan, M.H.; Xu, L. Comparison of the Hepatoprotective Effects of the Three Main Stilbenes from Mulberry Twigs. J. Agric. Food Chem. 2019, 67, 7245. [Google Scholar] [CrossRef]
- Niu, J.; Wang, K.; Kolattukudy, P.E. Cerium oxide nanoparticles inhibits oxidative stress and nuclear Factor-κB activation in H9c2 cardiomyocytes exposed to cigarette smoke extract. J. Pharmacol. Exp. Ther. 2011, 338, 53–61. [Google Scholar] [CrossRef]
- Zou, Y.; Kong, M. Tetrahydroxy stilbene glucoside alleviates palmitic acid-induced inflammation and apoptosis in cardiomyocytes by regulating miR-129-3p/Smad3 signaling. Cell Mol. Biol. Lett. 2019, 24, 5. [Google Scholar] [CrossRef]
- Boccellino, M.; Donniacuo, M.; Bruno, F.; Rinaldi, B.; Quagliuolo, L.; Ambruosi, M.; Pace, S.; De Rosa, M.; Olgaç, A.; Banoglu, E.; et al. Protective effect of piceatannol and bioactive stilbene derivatives against hypoxia-induced toxicity in H9c2 cardiomyocytes and structural elucidation as 5-LOX inhibitors. Eur. J. Med. Chem. 2019, 180, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H.; Kim, J.-E.; Song, N.R.; Son, J.E.; Hwang, M.K.; Byun, S.; Kim, J.H.; Lee, K.W.; Lee, H.J. Phosphoinositide 3-kinase is a novel target of piceatannol for inhibiting PDGF-BB-induced proliferation and migration in human aortic smooth muscle cells. Cardiovasc. Res. 2010, 85, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Seow, C.J.; Chue, S.C.; Wong, W.S.F. Piceatannol, a Syk-selective tyrosine kinase inhibitor, attenuated antigen challenge of guinea pig airways in vitro. Eur. J. Pharmacol. 2002, 443, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Liudvytska, O.; Ponczek, M.B.; Ciesielski, O.; Krzyżanowska-Kowalczyk, J.; Kowalczyk, M.; Balcerczyk, A.; Kolodziejczyk-Czepas, J. Rheum rhaponticum and Rheum rhabarbarum Extracts as Modulators of Endothelial Cell Inflammatory Response. Nutrients 2023, 15, 949. [Google Scholar] [CrossRef]
- Hu, X.B.; Kang, R.R.; Tang, T.T.; Li, Y.J.; Wu, J.Y.; Wang, J.M.; Liu, X.Y.; Xiang, D.X. Topical delivery of 3,5,4′-trimethoxy-trans-stilbene-loaded microemulsion-based hydrogel for the treatment of osteoarthritis in a rabbit model. Drug. Deliv. Transl. Res. 2019, 9, 357–365. [Google Scholar] [CrossRef]
- Wang, K.T.; Chen, L.G.; Tseng, S.H.; Huang, J.S.; Hsieh, M.S.; Wang, C.C. Anti-inflammatory effects of resveratrol and oligostilbenes from Vitis thunbergii var. taiwaniana against lipopolysaccharide-induced arthritis. J. Agric. Food Chem. 2011, 59, 8. [Google Scholar] [CrossRef]
- Tran, P.T.; Park, D.H.; Kim, O.; Kwon, S.H.; Min, B.S.; Lee, J.H. Desoxyrhapontigenin inhibits RANKL-induced osteoclast formation and prevents inflammation-mediated bone loss. Int. J. Mol. Med. 2018, 42, 569–578. [Google Scholar] [CrossRef]
- Susztak, K.; Raff, A.C.; Schiffer, M.; Böttinger, E.P. Glucose-induced reactive oxygen species cause apoptosis of podocytes and podocyte depletion at the onset of diabetic nephropathy. Diabetes 2006, 55, 894. [Google Scholar] [CrossRef]
- Navarro, J.; Morafernandez, C. The role of TNF-α in diabetic nephropathy: Pathogenic and therapeutic implications. Cytokine Growth Factor. Rev. 2006, 17, 441–450. [Google Scholar] [CrossRef]
- Vielhauer, V.; Mayadas, T.N. Functions of TNF and its Receptors in Renal Disease: Distinct Roles in Inflammatory Tissue Injury and Immune Regulation. Semin. Nephrol. 2007, 27, 286–308. [Google Scholar] [CrossRef]
- Lee, I.-T.; Yang, C.-M. Inflammatory Signalings Involved in Airway and Pulmonary Diseases. Mediat. Inflamm. 2013, 2013, 791231. [Google Scholar] [CrossRef]
- Lee, I.T.; Lin, C.F.; Huang, Y.L.; Chong, K.Y.; Hsieh, M.F.; Huang, T.H.; Cheng, C.Y. Protective mechanisms of resveratrol derivatives against TNF-α-induced inflammatory responses in rat mesangial cells. Cytokine 2019, 113, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Lan, Z.; Lin, Q.; Mi, X.; He, Y.; Wei, L.; Lin, Y.; Zhang, Y.; Deng, X. Polydatin ameliorates renal injury by attenuating oxidative stress-related inflammatory responses in fructose-induced urate nephropathic mice. Food Chem. Toxicol. 2013, 52, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Peng, J.; Huang, K.; Huang, J.; Shen, X.; Liu, P.; Huang, H. Polydatin ameliorates experimental diabetes-induced fibronectin through inhibiting the activation of NF-κB signaling pathway in rat glomerular mesangial cells. Mol. Cell Endocrinol. 2012, 362, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Zhou, R.; Tschopp, J. The NLRP3 inflammasome: A sensor for metabolic danger? Science 2010, 327, 5963. [Google Scholar] [CrossRef] [PubMed]
- Nunes, T.; De Souza, H.S. Inflammasome in intestinal inflammation and cancer. Mediat. Inflamm. 2013, 2013, 654963. [Google Scholar] [CrossRef]
- Li, J.; Wang, B.; Zhou, G.; Yan, X.; Zhang, Y. Tetrahydroxy Stilbene Glucoside Alleviates High Glucose-Induced MPC5 Podocytes Injury Through Suppression of NLRP3 Inflammasome. Am. J. Med. Sci. 2018, 355, 588–596. [Google Scholar] [CrossRef]
- Wang, P.-X.; Deng, X.-R.; Zhang, C.-H.; Yuan, H.-J. Gut microbiota and metabolic syndrome. Chin. Med. J. 2020, 133, 808–816. [Google Scholar] [CrossRef]
- Fan-Jiang, P.Y.; Lee, P.S.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Pterostilbene Attenuates High-Fat Diet and Dextran Sulfate Sodium-Induced Colitis via Suppressing Inflammation and Intestinal Fibrosis in Mice. J. Agric. Food Chem. 2021, 69, 25. [Google Scholar] [CrossRef]
- Shin, S.-H.; Song, J.-L.; Park, M.-G.; Park, M.-H.; Hwang, S.-J.; Park, K.-Y. Effects of natural raw meal (NRM) on high-fat diet and dextran sulfate sodium (DSS)-induced ulcerative colitis in C57BL/6J mice. Nutr. Res. Pract. 2015, 9, 619. [Google Scholar] [CrossRef]
- Sireesh, D.; Ganesh, M.-R.; Dhamodharan, U.; Sakthivadivel, M.; Sivasubramanian, S.; Gunasekaran, P.; Ramkumar, K.M. Role of pterostilbene in attenuating immune mediated devastation of pancreatic beta cells via Nrf2 signaling cascade. J. Nutr. Biochem. 2017, 44, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.K.; Kang, M.C.; Lee, S.; Darlami, O.; Shin, D.; Choi, I.; Kim, K.H.; Kim, S.Y. Generation of stilbene glycoside with promising cell rejuvenation activity through biotransformation by the entomopathogenic fungus Beauveria bassiana. Biomedicines 2021, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Heinzl, G.C.; Iametti, S.; Mattio, L.M.; Pinto, A.; Dallavalle, S.M.D.; Capraro, J.; Scarafoni, A. Polymeric Stilbene Derivatives in Winemaking Byproducts Affect NF-kB Mediated Inflammatory Response in Caco-2 Cells. SIB Congr. 2021. Available online: https://air.unimi.it/handle/2434/871256 (accessed on 15 April 2023).
- Kubyshkin, A.; Shevandova, A.; Petrenko, V.; Fomochkina, I.; Sorokina, L.; Kucherenko, A.; Gordienko, A.; Khimich, N.; Zyablitskaya, E.; Makalish, T.; et al. Anti-inflammatory and antidiabetic effects of grape-derived stilbene concentrate in the experimental metabolic syndrome. J. Diabetes Metab. Disord. 2020, 19, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, H.; Qin, L.; Zhang, Z.; Wang, Q.; Zhang, Q.; Lu, Z.; Wei, S.; Gao, X.; Tu, P. Global chemical profiling based quality evaluation approach of rhubarb using ultra performance liquid chromatography with tandem quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2015, 38, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kotlyarov, S.; Kotlyarova, A. Molecular Mechanisms of Lipid Metabolism Disorders in Infectious Exacerbations of Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2021, 22, 7634. [Google Scholar] [CrossRef]
- Shi, J.; Yin, N.; Xuan, L.L.; Yao, C.S.; Meng, A.M.; Hou, Q. Vam3, a derivative of resveratrol, attenuates cigarette smoke-induced autophagy. Acta Pharmacol. Sin. 2012, 33, 888–896. [Google Scholar] [CrossRef]
- de Oliveira, M.T.P.; Coutinho, D.d.S.; Guterres, S.S.; Pohlmann, A.R.; e Silva, P.M.R.; Martins, M.A.; Bernardi, A. Resveratrol-loaded lipid-core nanocapsules modulate acute lung inflammation and oxidative imbalance induced by lps in mice. Pharmaceutics 2021, 13, 683. [Google Scholar] [CrossRef]
- Da Silva, T.F.; Da Silva, T.F.; Teles, S.S.D.S.; De Assis, T.S.; Junior, M.S.O. Approaches from Resveratrol Activities on Central Nervous System Inflammation. Biotechnol. Res. 2019, 5, 28–34. Available online: http://br.biomedpress.org/index.php/br/article/view/783 (accessed on 15 April 2023).
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF Transcription Factors, Inflammation, and Immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef]
- Sharma, S.; Chopra, K.; Kulkarni, S.K.; Agrewala, J.N. Resveratrol and curcumin suppress immune response through CD28/CTLA-4 and CD80 co-stimulatory pathway. Clin. Exp. Immunol. 2007, 147, 155–163. [Google Scholar] [CrossRef]
- Singh, N.P.; Hegde, V.L.; Hofseth, L.J.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol (trans-3,5,4′-trihydroxystilbene) ameliorates experimental allergic encephalomyelitis, primarily via induction of apoptosis in T cells involving activation of aryl hydrocarbon receptor and estrogen receptor. Mol. Pharmacol. 2007, 72, 1508–1521. [Google Scholar] [CrossRef] [PubMed]
- Imler, T.J.; Petro, T.M. Decreased severity of experimental autoimmune encephalomyelitis during resveratrol administration is associated with increased IL-17+IL-10+ T cells, CD4- IFN-γ+ cells, and decreased macrophage IL-6 expression. Int. Immunopharmacol. 2009, 9, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Ates, O.; Cayli, S.; Altinoz, E.; Gurses, I.; Yucel, N.; Kocak, A.; Yologlu, S.; Turkoz, Y. Effects of resveratrol and methylprednisolone on biochemical, neurobehavioral and histopathological recovery after experimental spinal cord injury. Acta Pharmacol. Sin. 2006, 27, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xu, S.; Qian, Y.; Xiao, Q. Resveratrol regulates microglia M1/M2 polarization via PGC-1α in conditions of neuroinflammatory injury. Brain Behav. Immun. 2017, 64, 162–172. [Google Scholar] [CrossRef]
- Švajger, U.; Obermajer, N.; Jeras, M. Dendritic cells treated with resveratrol during differentiation from monocytes gain substantial tolerogenic properties upon activation. Immunology 2010, 129, 525–535. [Google Scholar] [CrossRef]
- Wight, R.D.; Tull, C.A.; Deel, M.W.; Stroope, B.L.; Eubanks, A.G.; Chavis, J.A.; Drew, P.D.; Hensley, L.L. Resveratrol effects on astrocyte function: Relevance to neurodegenerative diseases. Biochem. Biophys. Res. Commun. 2012, 426, 112–115. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, H.; Wang, L.; Tao, Y.; Du, G.; Guan, W.; Liu, J.; Brennan, C.; Ho, C.-T.; Li, S. Effects of Selected Resveratrol Analogues on Activation and Polarization of Lipopolysaccharide-Stimulated BV-2 Microglial Cells. J. Agric. Food Chem. 2020, 68, 3750–3757. [Google Scholar] [CrossRef]
- Ban, J.Y.; Jeon, S.-Y.; Nguyen, T.T.H.; Bae, K.; Song, K.-S.; Seonga, Y.H. Neuroprotective Effect of Oxyresveratrol from Smilacis Chinae Rhizome on Amyloid β Protein (25-35)-Induced Neurotoxicity in Cultured Rat Cortical Neurons. Biol. Pharm. Bull. 2006, 29, 2419–2424. [Google Scholar] [CrossRef]
- Weng, L.; Zhang, H.; Li, X.; Zhan, H.; Chen, F.; Han, L.; Xu, Y.; Cao, X. Ampelopsin attenuates lipopolysaccharide-induced inflammatory response through the inhibition of the NF-κB and JAK2/STAT3 signaling pathways in microglia. Int. Immunopharmacol. 2017, 44, 1–8. [Google Scholar] [CrossRef]
- Caillaud, M.; Guillard, J.; Richard, D.; Milin, S.; Chassaing, D.; Paccalin, M.; Page, G.; Bilan, A.R. Trans ε viniferin decreases amyloid deposits and inflammation in a mouse transgenic Alzheimer model. PLoS ONE 2019, 14, e0212663. [Google Scholar] [CrossRef]
- Ha, D.T.; Long, P.T.; Hien, T.T.; Tuan, D.T.; An, N.T.T.; Khoi, N.M.; Oanh, H.V.; Hung, T.M. Anti-inflammatory effect of oligostilbenoids from Vitis heyneana in LPS-stimulated RAW 264.7 macrophages via suppressing the NF-ΚB activation. Chem. Cent. J. 2018, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Chang, W.; Liu, B.; Chen, G.; Yang, Y.; Hao, Y.; Hou, Y.; Li, N. Stilbenes from the tubers of Bletilla striata with potential anti-neuroinflammatory activity. Bioorg. Chem. 2020, 97, 103715. [Google Scholar] [CrossRef] [PubMed]
- Petrovic-Djergovic, D.; Goonewardena, S.N.; Pinsky, D.J. Inflammatory disequilibrium in stroke. Circ. Res. 2016, 119, 142–158. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, L.; Yue, L.; Wang, B.; Li, X.; Guo, H.; Ma, Y.; Yao, C.; Gao, L.; Deng, J.; et al. Pterostilbene Attenuates Early Brain Injury Following Subarachnoid Hemorrhage via Inhibition of the NLRP3 Inflammasome and Nox2-Related Oxidative Stress. Mol. Neurobiol. 2017, 54, 5928–5940. [Google Scholar] [CrossRef]
- Liu, H.; Wu, X.; Luo, J.; Wang, X.; Guo, H.; Feng, D.; Zhao, L.; Bai, H.; Song, M.; Liu, X.; et al. Pterostilbene attenuates astrocytic inflammation and neuronal oxidative injury after ischemia-reperfusion by inhibiting nf-κb phosphorylation. Front. Immunol. 2019, 10, 2408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Deng, S.-M.; Chen, C.; He, Q.-H.; Peng, X.-W.; Liang, Q.-F.; Zhuang, G.-D.; Wang, S.-M.; Tang, D. Pterostilbene could alleviate diabetic cognitive impairment by suppressing TLR4/NF-κB pathway through microbiota-gut-brain axis. Phytother. Res. 2023, in press. [Google Scholar] [CrossRef]
- Akar, A.; Öztopuz, R.Ö.; Büyük, B.; Ovali, M.A.; Aykora, D.; Malçok, Ü.A. Neuroprotective Effects of Piceatannol on Olfactory Bulb Injury after Subarachnoid Hemorrhage. Mol. Neurobiol. 2023. [Google Scholar] [CrossRef]
- Mamada, N.; Tanokashira, D.; Ishii, K.; Tamaoka, A.; Araki, W. Mitochondria are devoid of amyloid β-protein (Aβ)-producing secretases: Evidence for unlikely occurrence within mitochondria of Aβ generation from amyloid precursor protein. Biochem. Biophys. Res. Commun. 2017, 486, 321–328. [Google Scholar] [CrossRef]
- Choi, C.W.; Choi, Y.H.; Cha, M.-R.; Kim, Y.S.; Yon, G.H.; Hong, K.S.; Park, W.-K.; Kim, Y.H.; Ryu, S.Y. In vitro BACE-1 inhibitory activity of resveratrol oligomers from the seed extract of Paeonia lactiflora. Planta Med. 2011, 77, 374–376. [Google Scholar] [CrossRef]
- Han, Y.S.; Zheng, W.H.; Bastianetto, S.; Chabot, J.G.; Quirion, R. Neuroprotective effects of resveratrol against β-amyloid-induced neurotoxicity in rat hippocampal neurons: Involvement of protein kinase C. Br. J. Pharmacol. 2004, 141, 997. [Google Scholar] [CrossRef]
- Chang, J.; Rimando, A.; Pallas, M.; Camins, A.; Porquet, D.; Reeves, J.; Shukitt-Hale, B.; Smith, M.A.; Joseph, J.A.; Casadesus, G. Low-dose pterostilbene, but not resveratrol, is a potent neuromodulator in aging and Alzheimer’s disease. Neurobiol. Aging 2012, 33, 2062–2071. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Yun, L.; Jiankui, G.; Zunjing, L.; Zhiqiang, Z.; Yong, W.; Zhao, L.; Xiangqun, S. Tetrahydroxy stilbene glucoside improved the behavioral disorders of APP695V717I transgenic mice by inhibiting the expression of Beclin-1 and LC3-II. J. Tradit. Chin. Med. 2015, 35, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, J.; Li, J.; Hu, C.; Zhang, L.; Yan, J.; Li, L.; Zhang, L. Tetrahydroxy stilbene glycoside alleviated inflammatory damage by mitophagy via AMPK related PINK1/Parkin signaling pathway. Biochem. Pharmacol. 2020, 177, 113997. [Google Scholar] [CrossRef] [PubMed]
- Gandy, K.A.O.; Zhang, J.; Nagarkatti, P.; Nagarkatti, M. Resveratrol (3, 5, 4′-Trihydroxy-trans-Stilbene) Attenuates a Mouse Model of Multiple Sclerosis by Altering the miR-124/Sphingosine Kinase 1 Axis in Encephalitogenic T Cells in the Brain. J. Neuroimmune Pharmacol. 2019, 14, 477. [Google Scholar] [CrossRef]
- Vion, E.; Page, G.; Bourdeaud, E.; Paccalin, M.; Guillard, J.; Bilan, A.R. Trans ε-viniferin is an amyloid-β disaggregating and anti-inflammatory drug in a mouse primary cellular model of Alzheimer’s disease. Mol. Cell Neurosci. 2018, 88, 1–6. [Google Scholar] [CrossRef]
- Trang, A.; Khandhar, P.B. Physiology, Acetylcholinesterase. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK539735/ (accessed on 16 April 2023).
- Cox, M.A.; Bassi, C.; Saunders, M.E.; Nechanitzky, R.; Morgado-Palacin, I.; Zheng, C.; Mak, T.W. Beyond neurotransmission: Acetylcholine in immunity and inflammation. J. Intern. Med. 2020, 287, 120–133. [Google Scholar] [CrossRef]
- McHardy, S.F.; Wang, H.Y.L.; McCowen, S.V.; Valdez, M.C. Recent advances in acetylcholinesterase Inhibitors and Reactivators: An update on the patent literature (2012–2015). Expert Opin. Ther. Pat. 2017, 27, 455–476. [Google Scholar] [CrossRef]
- Mlakić, M.; Odak, I.; Faraho, I.; Talić, S.; Bosnar, M.; Lasić, K.; Barić, D.; Škorić, I. New naphtho/thienobenzo-triazoles with interconnected anti-inflammatory and cholinesterase inhibitory activity. Eur. J. Med. Chem. 2022, 241, 114616. [Google Scholar] [CrossRef]
- Ślusarczyk, S.; Deniz, F.S.S.; Woźniak, D.; Pecio, L.; Pérez-Sánchez, H.; Cerón-Carrasco, J.P.; Stochmal, A.; den-Haan Alonso, H.; Matkowski, A.; Orhan, I.E. Selective in vitro and in silico cholinesterase inhibitory activity of isoflavones and stilbenes from Belamcandae chinensis rhizoma. Phytochem. Lett. 2019, 30, 261–272. [Google Scholar] [CrossRef]
- Agbo, E.N.; Gildenhuys, S.; Choong, Y.S.; Mphahlele, M.J.; More, G.K. Synthesis of furocoumarin–stilbene hybrids as potential multifunctional drugs against multiple biochemical targets associated with Alzheimer’s disease. Bioorg. Chem. 2020, 101, 103997. [Google Scholar] [CrossRef]
- Ohguchi, K.; Tanaka, T.; Iliya, I.; Ito, T.; Iinuma, M.; Matsumoto, K.; Akao, Y.; Nozawa, Y. Gnetol as a potent tyrosinase inhibitor from genus gnetum. Biosci. Biotechnol. Biochem. 2003, 67, 663–665. [Google Scholar] [CrossRef] [PubMed]
- Sermboonpaisarn, T.; Sawasdee, P. Potent and selective butyrylcholinesterase inhibitors from Ficus foveolata. Fitoterapia 2012, 83, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Kurundkar, D.; Kurundkar, A.R.; Bone, N.B.; Becker, E.J., Jr.; Liu, W.; Chacko, B.; Darley-Usmar, V.; Zmijewski, J.W.; Thannickal, V.J. SIRT3 diminishes inflammation and mitigates endotoxin-induced acute lung injury. JCI Insight 2019, 4, e120722. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, M.; Parhizkary, F.; Sharifi, R.; Ghotaslou, A.; Barati, M. Effects of resveratrol on coagulative, fibrinolytic, and inflammatory marker expression and secretion by endothelial cells (human umbilical vein endothelial cells). Blood Coagul. Fibrinolysis 2020, 31, 207–212. [Google Scholar] [CrossRef]
- Massimini, M.; Vedove, E.D.; Bachetti, B.; Di Pierro, F.; Ribecco, C.; D’Addario, C.; Pucci, M. Polyphenols and Cannabidiol Modulate Transcriptional Regulation of Th1/Th2 Inflammatory Genes Related to Canine Atopic Dermatitis. Front. Vet. Sci. 2021, 8, 606197. [Google Scholar] [CrossRef]
- Adhikari, N.; Aryal, Y.P.; Jung, J.K.; Ha, J.H.; Choi, S.Y.; Kim, J.Y.; Lee, T.H.; Kim, S.H.; Yamamoto, H.; Suh, J.Y.; et al. Resveratrol enhances bone formation by modulating inflammation in the mouse periodontitis model. J. Periodontal Res. 2021, 56, 735–745. [Google Scholar] [CrossRef]
- Hossain, E.; Anand-Srivastava, M.B. Resveratrol prevents angiotensin II-induced hypertrophy of vascular smooth muscle cells through the transactivation of growth factor receptors. Can. J. Physiol. Pharmacol. 2017, 95, 945–953. [Google Scholar] [CrossRef]
- Ho, Y.; Wu, C.Y.; Chin, Y.T.; Li, Z.L.; Pan, Y.S.; Huang, T.Y.; Su, P.Y.; Lee, S.Y.; Crawford, D.R.; Su, K.W.; et al. NDAT suppresses pro-inflammatory gene expression to enhance resveratrol-induced anti-proliferation in oral cancer cells. Food Chem. Toxicol. 2020, 136, 111092. [Google Scholar] [CrossRef]
- Cottart, C.-H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.-L. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef]
- Chen, W.; Yeo, S.C.M.; Elhennawy, M.G.A.A.; Lin, H.S. Oxyresveratrol: A bioavailable dietary polyphenol. J. Funct. Foods 2016, 22, 122–131. [Google Scholar] [CrossRef]
- Meenambiga, S.S.; Rajagopal, K. In silico studies on plant derived components of Cissus quadrangularis against COX-2 enzyme. Int. J. Pharm. Pharm. Sci. 2014, 6, 483–487. [Google Scholar]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.A.P.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Hyun, C.-G. Inhibitory Effects of Pinostilbene on Adipogenesis in 3T3-L1 Adipocytes: A Study of Possible Mechanisms. Int. J. Mol. Sci. 2021, 22, 13446. [Google Scholar] [CrossRef]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z. TN hompson, and DL McCormick, Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother. Pharmacol. 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Wang, P.; Sang, S. Metabolism and pharmacokinetics of resveratrol and pterostilbene. BioFactors 2018, 44, 16–25. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, X.; Cai, X.; Song, M.; Zheng, J.; Pan, C.; Qiu, P.; Zhang, L.; Zhou, S.; Tang, Z.; et al. Identification of pinostilbene as a major colonic metabolite of pterostilbene and its inhibitory effects on colon cancer cells. Mol. Nutr. Food Res. 2016, 60, 1924–1932. [Google Scholar] [CrossRef]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Kim, J.; Min, J.S.; Kim, D.; Zheng, Y.F.; Mailar, K.; Choi, W.J.; Lee, C.; Bae, S.K. A simple and sensitive liquid chromatography–tandem mass spectrometry method for trans-ε-viniferin quantification in mouse plasma and its application to a pharmacokinetic study in mice. J. Pharm. Biomed. Anal. 2017, 134, 116–121. [Google Scholar] [CrossRef]
- Courtois, A.; Jourdes, M.; Dupin, A.; Lapèze, C.; Renouf, E.; Biais, B.; Teissedre, P.-L.; Mérillon, J.-M.; Richard, T.; Krisa, S. In Vitro Glucuronidation and Sulfation of ε-Viniferin, a Resveratrol Dimer, in Humans and Rats. Molecules 2017, 22, 733. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Han, X.; Yang, Y.; Fu, A.; Li, G. Design and synthesis of 2,6-dihalogenated stilbene derivatives as potential anti-inflammatory and antitumor agents. Fitoterapia 2023, 167, 105493. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, R.; Bassin, P.; Cook, M.; Rossiter, S.; Martin, A.; Hutter, V. Late Breaking Abstract—Anti-inflammatory activity of novel transtilbene sulfonamide analogues as potential novel therapeutic agents for inflammatory lung disease. Eur. Respir. J. 2019, 54, pa2471. [Google Scholar] [CrossRef]
- Hilt, S.; Liu, R.; Maezawa, I.; Rojalin, T.; Aung, H.H.; Budamagunta, M.; Slez, R.; Gong, Q.; Carney, R.P.; Voss, J.C. Novel Stilbene-Nitroxyl Hybrid Compounds Display Discrete Modulation of Amyloid Beta Toxicity and Structure. Front. Chem. 2022, 10, 896386. [Google Scholar] [CrossRef]
- Navarro-Orcajada, S.; Conesa, I.; Vidal-Sánchez, F.J.; Matencio, A.; Albaladejo-Maricó, L.; García-Carmona, F.; López-Nicolás, J.M. Stilbenes: Characterization, bioactivity, encapsulation and structural modifications. A review of their current limitations and promising approaches. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Plant | Source of Extract | Stilbenes Present | Effects | Ref. |
|---|---|---|---|---|---|
| Vitaceae | Vitis amurensis Rupr. (Amur grapes) | Leaves, stem and roots | Dimer: Amurensin H | Effective against asthma and chronic obstructive pulmonary disease | [29] |
| Vitaceae | Vitis vinifera L. (European wine grape) | Vine shoot | Monomers: trans-PICE Dimers: Ampelopsin A Pallidol trans-Scirpusin A Vitisinol C δ-, ω-, ε-Viniferin Trimer: trans-Miyabenol C Tetramers: Hopeaphenol Isohopeaphenol, Vitisin A, B | Antioxidant activity, cytoprotective effect against β-amyloid-induced toxicity | [30] |
| Vitaceae | Vitis thunbergii Sieb. & Zucc. (Lobular grape) | Root and stem | RSV, Dimers: Vitisinols A–D (+)-ε-Viniferin Viniferal Trimer: Ampelopsin C Tetramers: Miyabenol A (+)-Vitisin A, (+)-Vitisin C | Anti-platelet, anti-oxidative | [31] |
| Paeoniaceae | Paeonia suffruticosa Andr. (Tree peony) | Seed coat | RSV Dimers: trans- and cis-ε-Viniferin Trimer: Suffruticosol A, B, C, D trans- and cis-Gnetin H | Protects against osteoarthritis in chondrocytes | [32] |
| Moraceae | Morus alba L. (Mulberry) | Fruits | Monomers: ORV Mulberroside A (MulA) | Anti-inflammation through MAPK and PI3K/AKT/p70S6K signaling pathways | [33] |
| Gnetaceae | Gnetum gnemon L. (Melinjo) | Roots | Monomer: GN | Reduces cell viability in colorectal cancer cells and inhibits platelet aggregation | [34] |
| Polygonaceae | Rheum undulatum L. (Rhubarb) | Rhizomes | Monomer: Desoxyrhapontigenin | Attenuates the NF-κB and MAPK pathways | [35] |
| Polygonaceae | Rheum rhaponticum L. (Rhapontic rhubarb) | Roots | Monomer: Rhaponticin Desoxyrhaponticin Dimer: Rhapontigenin Desoxyrhapontigenin | Anti-inflammatory, antioxidant and anti-cancer | [36] |
| Cyperaceae | Cyperus articulatus L. (Jointed flatsedge) | Rhizomes and roots | Monomer: PICE, Dimer: trans-Scirpusin B Trimer: Cyperusphenol B | Inhibition of NO, iNOS and COX-2 | [37] |
| Fabaceae | Arachis hypogaea L. (Tainan 9 peanut cultivar) | First germinated sprout | trans-arachidin-1 and -3 IPP IPD Monomer: RSV Dimer: Arahypin-7 | Binds with cannabinoid receptors, anti-carcinogenic | [38] |
| Fabaceae | Cajanus cajan L. (Pigeon pea) | Leaves | Longistyline A Cajaninstilbene acid | Prevents MRSA infections | [39,40] |
| Euphorbiaceae | Macaranga siamensis S. J. Davies (Kanda) | Leaves and twigs | Macasiamenenes A, B, F, K, L, and P | Apoptotic effects on THP-1 leukemia cell line | [41] |
| Poaceae | Triticum turgidum ssp. Durum (Senatore Cappelli wheat) | Whole grains | Pinosylvins | Anticancer, anti-inflammatory, antioxidant, neuroprotective, anti-allergic | [42] |
| Orchidaceae | Pholidota cantonensis Rolfe (Rattlesnake orchids) | Whole grass | cis-3,3′-dihydroxy-5- methoxystilbene, Thunalbene, Batatasin III | AChE inhibitory, nitric oxide (NO) inhibition | [43] |
| Caesalpiniaceae | Chamaecrista pumila (Lam.) K. Larsen (Dwarf Cassia) | Aerial parts | Chamaecristanols A, B | Laxative, heal wounds, treat ulcers, snake and scorpion bites | [23,44] |
| Pinaceae | Picea abies L. (Norway spruce) | Bark | Astringin Isorhapontigenin Isorhapontin | Inhibits NO production without decreasing oxidative stress | [45] |
| Ericaceae | Vaccinium stamineum L. (Deerberry) | Berry | Pterostilbene Piceatannol | Inhibits activator protein-1 (AP-1) and NF- κB. Induced apoptosis of HL-60 cancer cells | [46,47] |
| Stilbenes | Concentration | Model System | Observation | Ref. |
|---|---|---|---|---|
| Amurensin H | 4 and 8 μmol/L | IL-1β-induced rat knee chondrocytes | Blocked elevation in IL-6, IL-17, TNFα, TLR4, TRAF6, Syk phosphorylation, NO levels and iNOS expression. Decreased PGE2 and COX-2 levels. Up-regulated COL2A1 and GAG, major components of ECM | [29] |
| 20 mg/kg | MIA-induced mice | Alleviated bone wear and cartilage loss from 60 ± 13.4% to 25.5 ± 11.4%. | ||
| 3,5,4′-Trimethoxy-trans-stilbene (BTM) | 1% and 0.5% | Rabbit knee joints | Topical administration reduced inflammatory cell infiltration, decreased bone destruction and roughness, and inhibited fibrous connective tissue proliferation. Microemulsion-based hydrogel method decreased IL-1β and TNFα. | [106] |
| Ampelopsin C | 25 μM | Human chondrocytes | Inhibits PGE2 with IC50 15.52 μM | [107] |
| Desoxyrhapontigenin | 50 mg/kg | LPS-stimulated mice and RANKL-induced osteoclastogenesis. | LPS induced trabecular separation, while bone surface and volume changes were attenuated. Anti-osteoporosis activity by inhibiting RANKL. Osteoclast formation is suppressed at an early stage by inhibiting the MAPK/AP-1 signaling pathway, ERK phosphorylation, and the expression of c-Fos and NFATc1. | [108] |
| Stilbenes | Concentration | Model System | Observation | Ref. |
|---|---|---|---|---|
| ε-viniferin | 120 mg/kg | Endotoxin-induced ALI in mice | During lung injury, they decrease inflammation through SIRT3 expression. | [166] |
| RSV | 100 mg/mL | Human umbilical vein endothelial cells (HUVECs) | Reduced amounts of factor VIII, t-PA-1, neutrophils recruiting factor von Willebrand factor (VWF) and IL-8. | [167] |
| Longistylin A | 10 µg | Mice model | Topical application on skin prevented MRSA infection in wounds, decreased IL-6 and TNFα. | [39] |
| Cannabidiol, luteolin and PICE | Mixture in ratio 10:25:25 µM | Canine epidermal keratinocytes (CPEK) | 8 h exposure increased cell viability, decreased DNA methylation in the promoter of ccl17, and increased it in tslp during inflammation. | [168] |
| 10:50:50 µM and 10:100:100 µM | Canine Macrophage DH82 Cell Line | Cell viability decreased, methylation increased in one il31ra and three ccl17 sites, and decreased in one ccl17 site. | ||
| RSV | 50 μM | Mice Periodontitis model | Decrease in IL-6 localization pattern, TNFα, MPO, Ki67-positive (tumor tendency), RUNX2-positive and osteoclastic cells. Increases bone tissue mass and healing, osteoblasts, collagen, CD31 cells. Macrophage marker F4/80. | [169] |
| RSV | 30 µM | Cells of vascular smooth muscle stimulated by angiotensin II | Resists hypertrophy via PDGFR, NADPH oxidase, EGFR and c-Src activation and it also links to the ERK1/2 and Akt signaling pathways. | [170] |
| RSV with NDAT | 10 μM of RSV and 10−7 M of NDAT | Mouth cancer SCC-25 and OEC-M1 cells | Reduction in IL-1β and TNFα, STAT3 phosphorylation and accumulation. Moderates integrin ligands, which control the immunomodulating PD-1/PD-L1 checkpoint. | [171] |
| Rhapontigenin | 1 µg/mL | HUVECs | Inhibition of 5-LOX | [105] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khayri, J.M.; Mascarenhas, R.; Harish, H.M.; Gowda, Y.; Lakshmaiah, V.V.; Nagella, P.; Al-Mssallem, M.Q.; Alessa, F.M.; Almaghasla, M.I.; Rezk, A.A.-S. Stilbenes, a Versatile Class of Natural Metabolites for Inflammation—An Overview. Molecules 2023, 28, 3786. https://doi.org/10.3390/molecules28093786
Al-Khayri JM, Mascarenhas R, Harish HM, Gowda Y, Lakshmaiah VV, Nagella P, Al-Mssallem MQ, Alessa FM, Almaghasla MI, Rezk AA-S. Stilbenes, a Versatile Class of Natural Metabolites for Inflammation—An Overview. Molecules. 2023; 28(9):3786. https://doi.org/10.3390/molecules28093786
Chicago/Turabian StyleAl-Khayri, Jameel M., Roseanne Mascarenhas, Himanshu Madapur Harish, Yashwanth Gowda, Vasantha Veerappa Lakshmaiah, Praveen Nagella, Muneera Qassim Al-Mssallem, Fatima Mohammed Alessa, Mustafa Ibrahim Almaghasla, and Adel Abdel-Sabour Rezk. 2023. "Stilbenes, a Versatile Class of Natural Metabolites for Inflammation—An Overview" Molecules 28, no. 9: 3786. https://doi.org/10.3390/molecules28093786