

β-Barrel Assembly Machinery (BAM) Complex as Novel Antibacterial Drug Target


Abstract
:1. Introduction
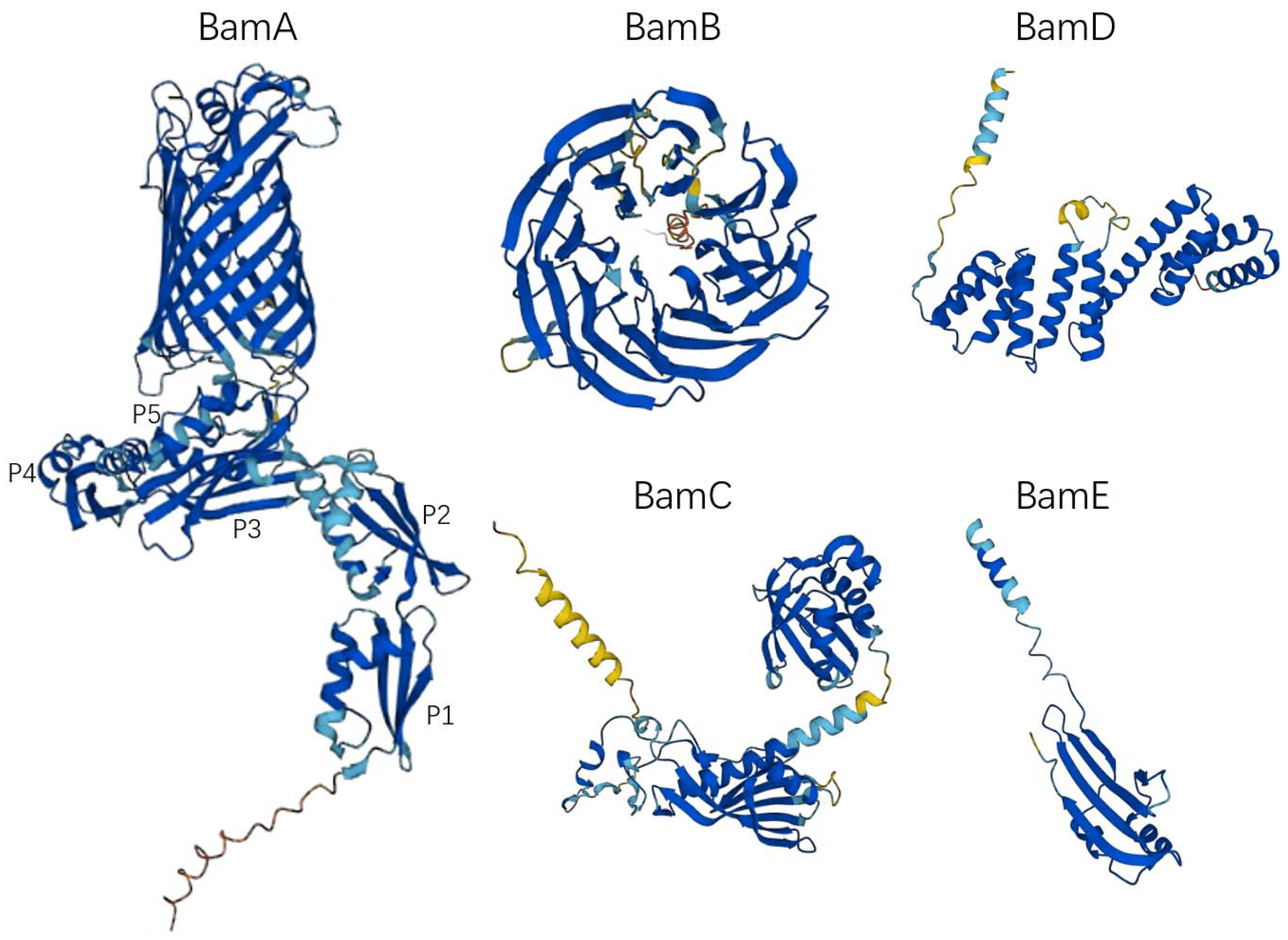
2. The Structure of the BAM Complex
2.1. BamA
2.2. BamB
2.3. BamC
2.4. BamD
2.5. BamE
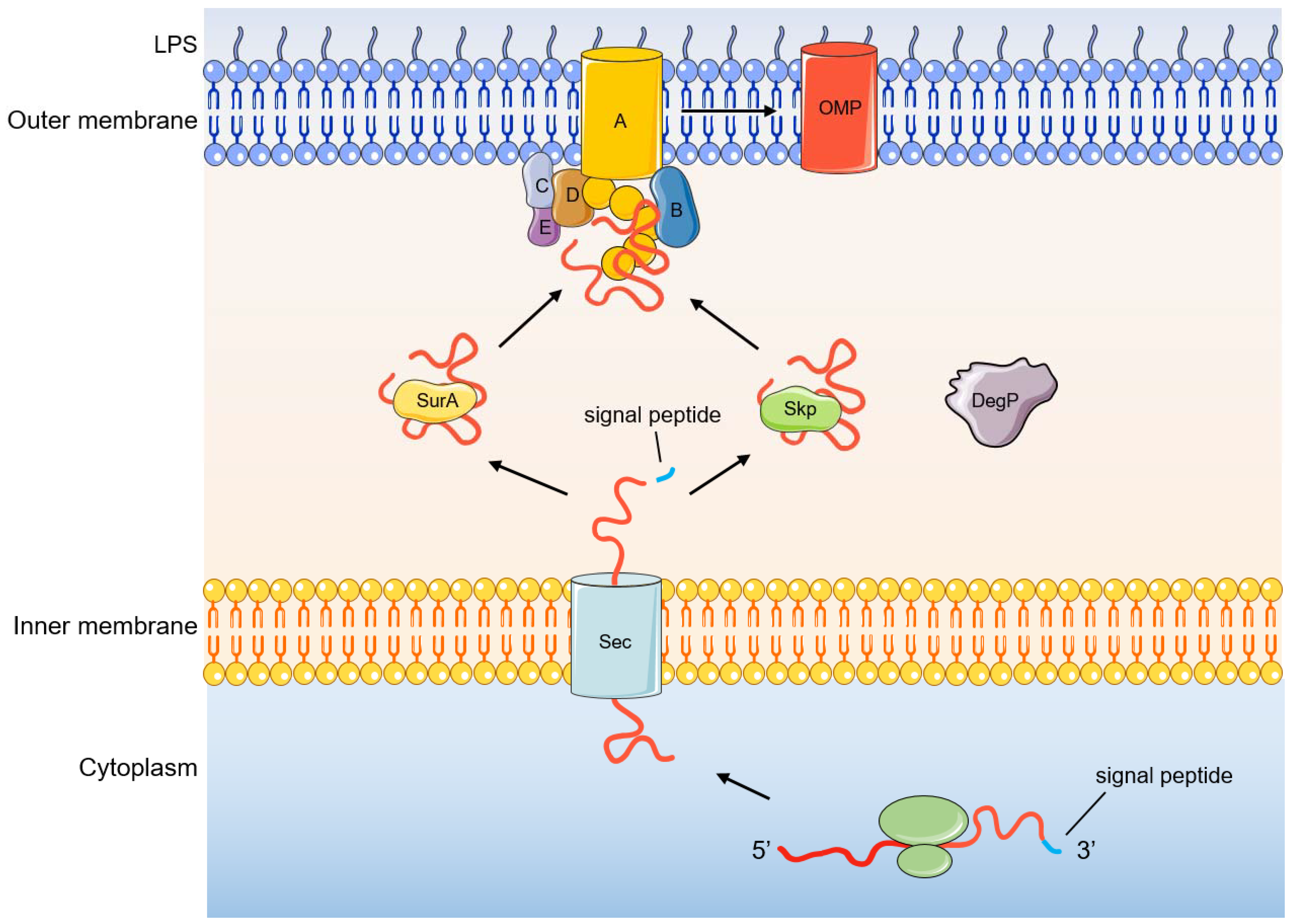
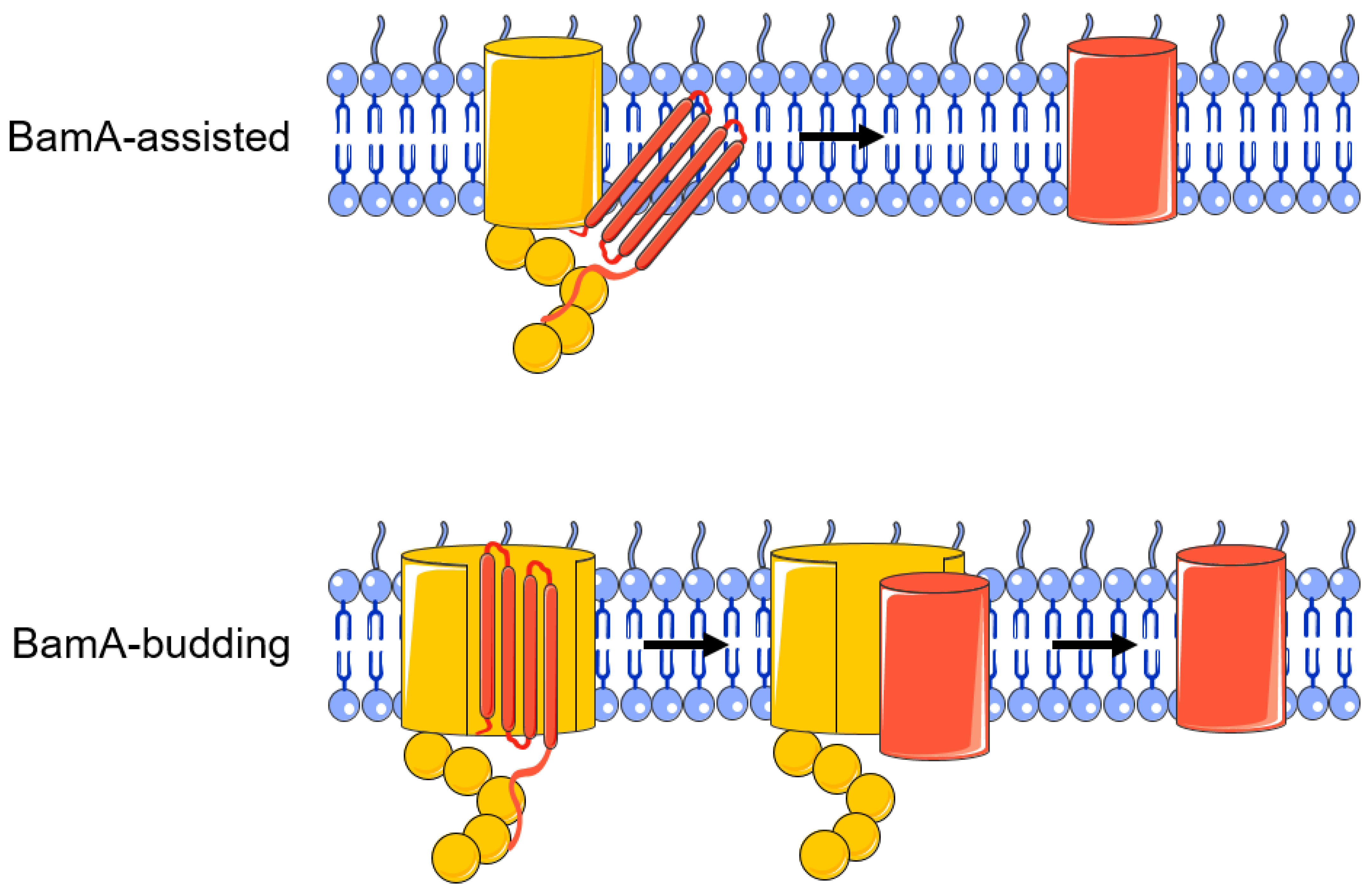
3. Folding and Insertion of OMPs by the BAM Complex
4. Research Progress of Drugs Targeting the BAM Complex
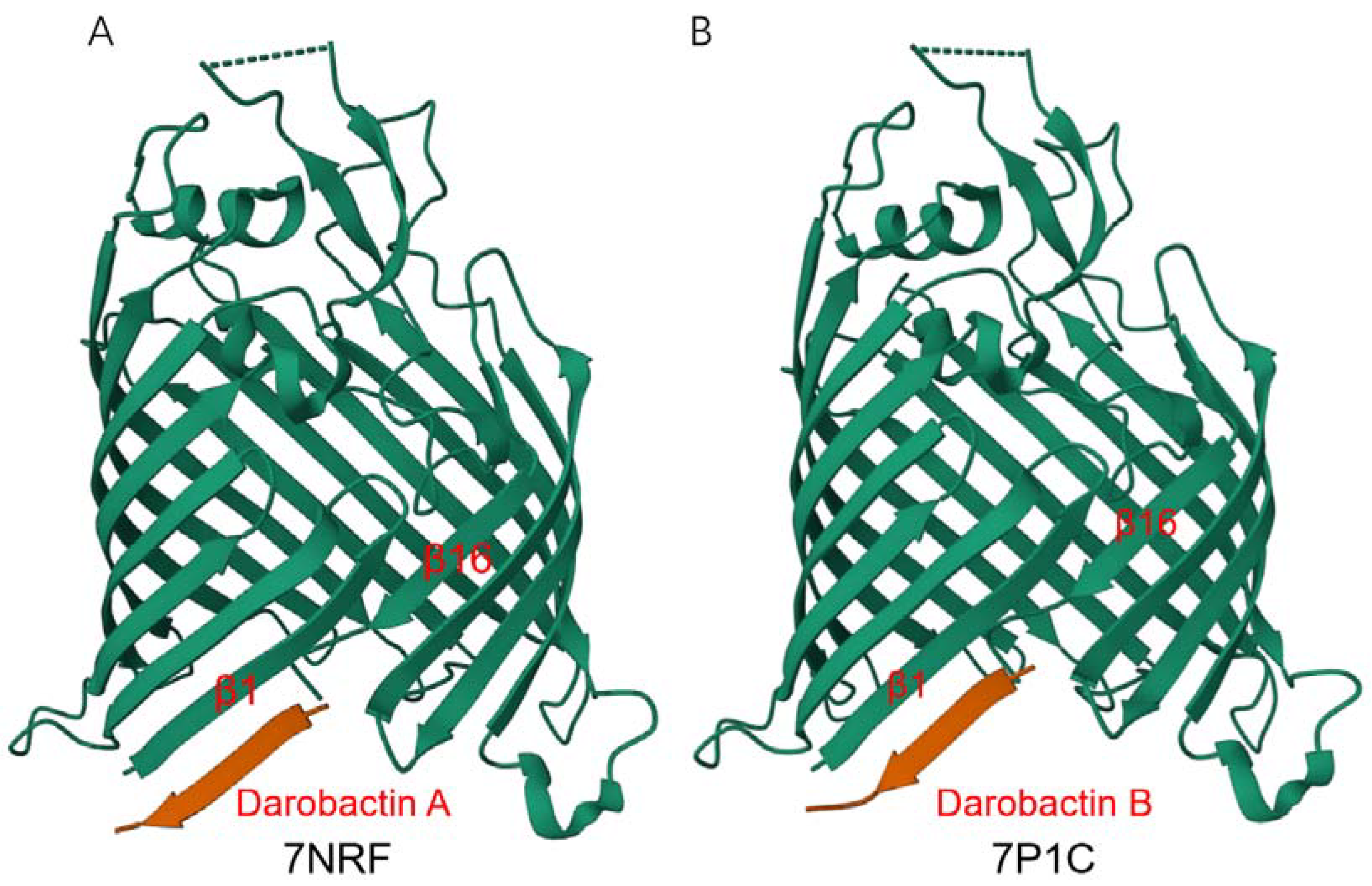
4.1. Darobactin
4.2. JB-95
4.3. LlpAs
4.4. MRL-494
4.5. MAB1
4.6. IMB-H4
4.7. FIRL
4.8. BamD Inhibitory Peptide
4.9. NTZ
4.10. VUF15259
5. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cavallo, G. The role of antibiotics in the evolution of microorganisms. G Di Batteriol. Virol. Ed. Immunol. 1992, 85, 3–11. [Google Scholar]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic Resistance and the MRSA Problem. Microbiol. Spectr. 2019, 7, GPP3-0057-2018. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Humphreys, G.; Fleck, F. United Nations meeting on antimicrobial resistance. Bull. World Health Organ. 2016, 94, 638–639. [Google Scholar] [CrossRef]
- Chahine, E.B.; Dougherty, J.A.; Thornby, K.-A.; Guirguis, E.H. Antibiotic Approvals in the Last Decade: Are We Keeping Up with Resistance? Ann. Pharmacother. 2022, 56, 441–462. [Google Scholar] [CrossRef]
- Rajagopal, M.; Walker, S. Envelope Structures of Gram-Positive Bacteria. In Protein and Sugar Export and Assembly in Gram-positive Bacteria; Bagnoli, F., Rappuoli, R., Eds.; Springer: Cham, Switzerland, 2017; Volume 404, pp. 1–44. [Google Scholar] [CrossRef]
- Bos, M.P.; Robert, V.; Tommassen, J. Biogenesis of the Gram-Negative Bacterial Outer Membrane. Annu. Rev. Microbiol. 2007, 61, 191–214. [Google Scholar] [CrossRef]
- Wu, T.; Malinverni, J.; Ruiz, N.; Kim, S.; Silhavy, T.J.; Kahne, D. Identification of a Multicomponent Complex Required for Outer Membrane Biogenesis in Escherichia coli. Cell 2005, 121, 235–245. [Google Scholar] [CrossRef]
- Konovalova, A.; Kahne, D.E.; Silhavy, T.J. Outer Membrane Biogenesis. Annu. Rev. Microbiol. 2017, 71, 539–556. [Google Scholar] [CrossRef]
- Cox, G.; Wright, G.D. Intrinsic antibiotic resistance: Mechanisms, origins, challenges and solutions. Int. J. Med. Microbiol. 2013, 303, 287–292. [Google Scholar] [CrossRef]
- Masi, M.; Winterhalter, M.; Pagès, J.-M. Outer Membrane Porins. Bact. Cell Walls Membr. 2019, 92, 79–123. [Google Scholar] [CrossRef]
- Fischer, N.; Raunest, M.; Schmidt, T.H.; Koch, D.C.; Kandt, C. Efflux pump-mediated antibiotics resistance: Insights from computational structural biology. Interdiscip. Sci. Comput. Life Sci. 2014, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Paulsson, M.; Kragh, K.N.; Su, Y.-C.; Sandblad, L.; Singh, B.; Bjarnsholt, T.; Riesbeck, K. Peptidoglycan-Binding Anchor Is a Pseudomonas aeruginosa OmpA Family Lipoprotein with Importance for Outer Membrane Vesicles, Biofilms, and the Periplasmic Shape. Front. Microbiol. 2021, 12, 639582. [Google Scholar] [CrossRef] [PubMed]
- Nie, D.; Hu, Y.; Chen, Z.; Li, M.; Hou, Z.; Luo, X.; Mao, X.; Xue, X. Outer membrane protein A (OmpA) as a potential therapeutic target for Acinetobacter baumannii infection. J. Biomed. Sci. 2020, 27, 26. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Bhowmick, B.; Upadhyay, A.; Han, Q. Structure and Functions of Bacterial Outer Membrane Protein A, A Potential Therapeutic Target for Bacterial Infection. Curr. Top. Med. Chem. 2021, 21, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.E.; Sinsinbar, G.; Gudlur, S.; Nallani, M.; Huang, C.-F.; Liedberg, B.; Mrksich, M. A Bottom-Up Proteomic Approach to Identify Substrate Specificity of Outer-Membrane Protease OmpT. Angew. Chem. Int. Ed. 2017, 56, 16531–16535. [Google Scholar] [CrossRef] [PubMed]
- Fairman, J.W.; Noinaj, N.; Buchanan, S.K. The structural biology of β-barrel membrane proteins: A summary of recent reports. Curr. Opin. Struct. Biol. 2011, 21, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Gatsos, X.; Perry, A.J.; Anwari, K.; Dolezal, P.; Wolynec, P.P.; Likić, V.A.; Purcell, A.W.; Buchanan, S.K.; Lithgow, T. Protein secretion and outer membrane assembly in Alphaproteobacteria. FEMS Microbiol. Rev. 2008, 32, 995–1009. [Google Scholar] [CrossRef] [PubMed]
- Schiffrin, B.; Brockwell, D.J.; Radford, S.E. Outer membrane protein folding from an energy landscape perspective. BMC Biol. 2017, 15, 153. [Google Scholar] [CrossRef] [PubMed]
- Ricci, D.P.; Silhavy, T.J. Outer Membrane Protein Insertion by the β-barrel Assembly Machine. EcoSal Plus 2019, 8, ESP-0035-2018. [Google Scholar] [CrossRef] [PubMed]
- Hagan, C.L.; Silhavy, T.J.; Kahne, D. β-Barrel Membrane Protein Assembly by the Bam Complex. Annu. Rev. Biochem. 2011, 80, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Malinverni, J.C.; Werner, J.; Kim, S.; Sklar, J.G.; Kahne, D.; Misra, R.; Silhavy, T.J. YfiO stabilizes the YaeT complex and is essential for outer membrane protein assembly in Escherichia coli. Mol. Microbiol. 2006, 61, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Voulhoux, R.; Bos, M.P.; Geurtsen, J.; Mols, M.; Tommassen, J. Role of a Highly Conserved Bacterial Protein in Outer Membrane Protein Assembly. Science 2003, 299, 262–265. [Google Scholar] [CrossRef]
- Doerrler, W.T.; Raetz, C.R. Loss of Outer Membrane Proteins without Inhibition of Lipid Export in an Escherichia coli YaeT Mutant. J. Biol. Chem. 2005, 280, 27679–27687. [Google Scholar] [CrossRef] [PubMed]
- Voulhoux, R.; Tommassen, J. Omp85, an evolutionarily conserved bacterial protein involved in outer-membrane-protein assembly. Res. Microbiol. 2004, 155, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Gentle, I.E.; Burri, L.; Lithgow, T. Molecular architecture and function of the Omp85 family of proteins. Mol. Microbiol. 2005, 58, 1216–1225. [Google Scholar] [CrossRef]
- Ni, D.; Wang, Y.; Yang, X.; Zhou, H.; Hou, X.; Cao, B.; Lu, Z.; Zhao, X.; Yang, K.; Huang, Y. Structural and functional analysis of the β-barrel domain of BamA from Escherichia coli. FASEB J. 2014, 28, 2677–2685. [Google Scholar] [CrossRef]
- Noinaj, N.; Kuszak, A.J.; Gumbart, J.C.; Lukacik, P.; Chang, H.; Easley, N.C.; Lithgow, T.; Buchanan, S.K. Structural insight into the biogenesis of β-barrel membrane proteins. Nature 2013, 501, 385–390. [Google Scholar] [CrossRef]
- Gatzeva-Topalova, P.Z.; Warner, L.R.; Pardi, A.; Sousa, M.C. Structure and Flexibility of the Complete Periplasmic Domain of BamA: The Protein Insertion Machine of the Outer Membrane. Structure 2010, 18, 1492–1501. [Google Scholar] [CrossRef]
- Gatzeva-Topalova, P.Z.; Walton, T.A.; Sousa, M.C. Crystal Structure of YaeT: Conformational Flexibility and Substrate Recognition. Structure 2008, 16, 1873–1881. [Google Scholar] [CrossRef]
- Sánchez-Pulido, L.; Devos, D.; Genevrois, S.; Vicente, M.; Valencia, A. POTRA: A conserved domain in the FtsQ family and a class of β-barrel outer membrane proteins. Trends Biochem. Sci. 2003, 28, 523–526. [Google Scholar] [CrossRef]
- Kim, S.; Malinverni, J.C.; Sliz, P.; Silhavy, T.J.; Harrison, S.C.; Kahne, D. Structure and Function of an Essential Component of the Outer Membrane Protein Assembly Machine. Science 2007, 317, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Bos, M.P.; Robert, V.; Tommassen, J. Functioning of outer membrane protein assembly factor Omp85 requires a single POTRA domain. EMBO Rep. 2007, 8, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Paetzel, M. Crystal Structure of Escherichia coli BamB, a Lipoprotein Component of the β-Barrel Assembly Machinery Complex. J. Mol. Biol. 2011, 406, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Jansen, K.B.; Baker, S.L.; Sousa, M.C. Crystal Structure of BamB Bound to a Periplasmic Domain Fragment of BamA, the Central Component of the β-Barrel Assembly Machine. J. Biol. Chem. 2015, 290, 2126–2136. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhan, L.-H.; Hou, H.-F.; Gao, Z.-Q.; Xu, J.-H.; Dong, C.; Dong, Y.-H. Structural basis for the interaction of BamB with the POTRA3–4 domains of BamA. Acta Crystallogr. Sect. D Struct. Biol. 2016, 72, 236–244. [Google Scholar] [CrossRef]
- Noinaj, N.; Fairman, J.W.; Buchanan, S.K. The Crystal Structure of BamB Suggests Interactions with BamA and Its Role within the BAM Complex. J. Mol. Biol. 2011, 407, 248–260. [Google Scholar] [CrossRef]
- Ruiz, N.; Falcone, B.; Kahne, D.; Silhavy, T.J. Chemical Conditionality: A GeneticStrategy to Probe Organelle Assembly. Cell 2005, 121, 307–317. [Google Scholar] [CrossRef]
- Hsieh, P.-F.; Hsu, C.-R.; Chen, C.-T.; Lin, T.-L.; Wang, J.-T. TheKlebsiella pneumoniaeYfgL (BamB) lipoprotein contributes to outer membrane protein biogenesis, type-1 fimbriae expression, anti-phagocytosis, andin vivovirulence. Virulence 2016, 7, 587–601. [Google Scholar] [CrossRef]
- Rolhion, N.; Barnich, N.; Claret, L.; Darfeuille-Michaud, A. Strong Decrease in Invasive Ability and Outer Membrane Vesicle Release in Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Strain LF82 with the yfgL Gene Deleted. J. Bacteriol. 2005, 187, 2286–2296. [Google Scholar] [CrossRef]
- Fardini, Y.; Chettab, K.; Grépinet, O.; Rochereau, S.; Trotereau, J.; Harvey, P.; Amy, M.; Bottreau, E.; Bumstead, N.; Barrow, P.A.; et al. The YfgL Lipoprotein Is Essential for Type III Secretion System Expression and Virulence of Salmonella enterica Serovar Enteritidis. Infect. Immun. 2007, 75, 358–370. [Google Scholar] [CrossRef]
- Ureta, A.R.; Endres, R.G.; Wingreen, N.S.; Silhavy, T.J. Kinetic Analysis of the Assembly of the Outer Membrane Protein LamB in Escherichia coli Mutants Each Lacking a Secretion or Targeting Factor in a Different Cellular Compartment. J. Bacteriol. 2007, 189, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Gunasinghe, S.D.; Shiota, T.; Stubenrauch, C.J.; Schulze, K.E.; Webb, C.T.; Fulcher, A.J.; Dunstan, R.A.; Hay, I.D.; Naderer, T.; Whelan, D.R.; et al. The WD40 Protein BamB Mediates Coupling of BAM Complexes into Assembly Precincts in the Bacterial Outer Membrane. Cell Rep. 2018, 23, 2782–2794. [Google Scholar] [CrossRef] [PubMed]
- Heuck, A.; Schleiffer, A.; Clausen, T. Augmenting β-Augmentation: Structural Basis of How BamB Binds BamA and May Support Folding of Outer Membrane Proteins. J. Mol. Biol. 2011, 406, 659–666. [Google Scholar] [CrossRef]
- Warner, L.R.; Varga, K.; Lange, O.F.; Baker, S.L.; Baker, D.; Sousa, M.C.; Pardi, A. Structure of the BamC Two-Domain Protein Obtained by Rosetta with a Limited NMR Data Set. J. Mol. Biol. 2011, 411, 83–95. [Google Scholar] [CrossRef]
- Kim, K.H.; Aulakh, S.; Paetzel, M. Crystal Structure of β-Barrel Assembly Machinery BamCD Protein Complex. J. Biol. Chem. 2011, 286, 39116–39121. [Google Scholar] [CrossRef]
- Sklar, J.G.; Wu, T.; Gronenberg, L.S.; Malinverni, J.C.; Kahne, D.; Silhavy, T.J. Lipoprotein SmpA is a component of the YaeT complex that assembles outer membrane proteins in Escherichia coli. Proc. Natl. Acad. Sci. USA 2007, 104, 6400–6405. [Google Scholar] [CrossRef]
- Onufryk, C.; Crouch, M.-L.; Fang, F.C.; Gross, C.A. Characterization of Six Lipoproteins in the σ E Regulon. J. Bacteriol. 2005, 187, 4552–4561. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.T.; Selkrig, J.; Perry, A.J.; Noinaj, N.; Buchanan, S.K.; Lithgow, T. Dynamic Association of BAM Complex Modules Includes Surface Exposure of the Lipoprotein BamC. J. Mol. Biol. 2012, 422, 545–555. [Google Scholar] [CrossRef]
- Xiang, S.; Pinto, C.; Baldus, M. Divide and Conquer: A Tailored Solid-state NMR Approach to Study Large Membrane Protein Complexes. Angew. Chem. Int. Ed. 2022, 61, e202203319. [Google Scholar] [CrossRef]
- Sandoval, C.M.; Baker, S.L.; Jansen, K.; Metzner, S.I.; Sousa, M.C. Crystal Structure of BamD: An Essential Component of the β-Barrel Assembly Machinery of Gram-Negative Bacteria. J. Mol. Biol. 2011, 409, 348–357. [Google Scholar] [CrossRef]
- Hagan, C.L.; Westwood, D.B.; Kahne, D. Bam Lipoproteins Assemble BamA in Vitro. Biochemistry 2013, 52, 6108–6113. [Google Scholar] [CrossRef] [PubMed]
- Hagan, C.L.; Wzorek, J.S.; Kahne, D. Inhibition of the β-barrel assembly machine by a peptide that binds BamD. Proc. Natl. Acad. Sci. USA 2015, 112, 2011–2016. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, R.; Zeth, K. Structural Basis of Outer Membrane Protein Biogenesis in Bacteria. J. Biol. Chem. 2011, 286, 27792–27803. [Google Scholar] [CrossRef]
- Lecker, S.H.; Driessen, A.J.; Wickner, W. ProOmpA contains secondary and tertiary structure prior to translocation and is shielded from aggregation by association with SecB protein. EMBO J. 1990, 9, 2309–2314. [Google Scholar] [CrossRef] [PubMed]
- Ernst, F.; Hoffschulte, H.; Thome-Kromer, B.; Swidersky, U.; Werner, P.; Müller, M. Precursor-specific requirements for SecA, SecB, and delta muH+ during protein export of Escherichia coli. J. Biol. Chem. 1994, 269, 12840–12845. [Google Scholar] [CrossRef]
- Gelis, I.; Bonvin, A.M.; Keramisanou, D.; Koukaki, M.; Gouridis, G.; Karamanou, S.; Economou, A.; Kalodimos, C.G. Structural Basis for Signal-Sequence Recognition by the Translocase Motor SecA as Determined by NMR. Cell 2007, 131, 756–769. [Google Scholar] [CrossRef]
- Chen, Y.; Tai, P.C.; Sui, S.-F. The active ring-like structure of SecA revealed by electron crystallography: Conformational change upon interaction with SecB. J. Struct. Biol. 2007, 159, 149–153. [Google Scholar] [CrossRef]
- Berg, B.V.D.; Clemons, W.M., Jr.; Collinson, I.; Modis, Y.; Hartmann, E.; Harrison, S.C.; Rapoport, T.A. X-ray structure of a protein-conducting channel. Nature 2004, 427, 36–44. [Google Scholar] [CrossRef]
- Behrens, S.; Maier, R.; de Cock, H.; Schmid, F.X.; Gross, C.A. The SurA periplasmic PPIase lacking its parvulin domains functions in vivo and has chaperone activity. EMBO J. 2001, 20, 285–294. [Google Scholar] [CrossRef]
- Hennecke, G.; Nolte, J.; Volkmer-Engert, R.; Schneider-Mergener, J.; Behrens, S. The Periplasmic Chaperone SurA Exploits Two Features Characteristic of Integral Outer Membrane Proteins for Selective Substrate Recognition. J. Biol. Chem. 2005, 280, 23540–23548. [Google Scholar] [CrossRef]
- Sklar, J.G.; Wu, T.; Kahne, D.; Silhavy, T.J. Defining the roles of the periplasmic chaperones SurA, Skp, and DegP in Escherichia coli. Genes Dev. 2007, 21, 2473–2484. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Henning, U. Aperiplasmic protein (Skp) of Escherichia coli selectively binds a class of outer membrane proteins. Mol. Microbiol. 1996, 19, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Lipinska, B.; Zylicz, M.; Georgopoulos, C. The HtrA (DegP) protein, essential for Escherichia coli survival at high temperatures, is an endopeptidase. J. Bacteriol. 1990, 172, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Spiess, C.; Beil, A.; Ehrmann, M. A Temperature-Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein. Cell 1999, 97, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Wang, R.; Ma, J.; Liu, Y.; Ezemaduka, A.N.; Chen, P.R.; Fu, X.; Chang, Z. DegP primarily functions as a protease for the biogenesis of β-barrel outer membrane proteins in the Gram-negative bacterium Escherichia coli. FEBS J. 2014, 281, 1226–1240. [Google Scholar] [CrossRef]
- Vertommen, D.; Ruiz, N.; Leverrier, P.; Silhavy, T.J.; Collet, J.-F. Characterization of the role of the Escherichia coli periplasmic chaperone SurA using differential proteomics. Proteomics 2009, 9, 2432–2443. [Google Scholar] [CrossRef] [PubMed]
- Denoncin, K.; Schwalm, J.; Vertommen, D.; Silhavy, T.J.; Collet, J.-F. Dissecting the Escherichia coli periplasmic chaperone network using differential proteomics. Proteomics 2012, 12, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Rollauer, S.E.; Sooreshjani, M.A.; Noinaj, N.; Buchanan, S.K. Outer membrane protein biogenesis in Gram-negative bacteria. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20150023. [Google Scholar] [CrossRef]
- Rizzitello, A.E.; Harper, J.R.; Silhavy, T.J. Genetic Evidence for Parallel Pathways of Chaperone Activity in the Periplasm of Escherichia coli. J. Bacteriol. 2001, 183, 6794–6800. [Google Scholar] [CrossRef]
- Noinaj, N.; Rollauer, S.E.; Buchanan, S.K. The β-barrel membrane protein insertase machinery from Gram-negative bacteria. Curr. Opin. Struct. Biol. 2015, 31, 35–42. [Google Scholar] [CrossRef]
- Fleming, K.G. A combined kinetic push and thermodynamic pull as driving forces for outer membrane protein sorting and folding in bacteria. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20150026. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H. Folding of β-barrel membrane proteins in lipid bilayers—Unassisted and assisted folding and insertion. Biochim. Et Biophys. Acta BBA Biomembr. 2015, 1848, 1927–1943. [Google Scholar] [CrossRef]
- Burgess, N.K.; Dao, T.P.; Stanley, A.M.; Fleming, K.G. β-Barrel Proteins That Reside in the Escherichia coli Outer Membrane in Vivo Demonstrate Varied Folding Behavior in Vitro. J. Biol. Chem. 2008, 283, 26748–26758. [Google Scholar] [CrossRef]
- Gessmann, D.; Chung, Y.H.; Danoff, E.J.; Plummer, A.M.; Sandlin, C.W.; Zaccai, N.R.; Fleming, K.G. Outer membrane β-barrel protein folding is physically controlled by periplasmic lipid head groups and BamA. Proc. Natl. Acad. Sci. USA 2014, 111, 5878–5883. [Google Scholar] [CrossRef] [PubMed]
- Noinaj, N.; Kuszak, A.; Balusek, C.; Gumbart, J.C.; Buchanan, S.K. Lateral Opening and Exit Pore Formation Are Required for BamA Function. Structure 2014, 22, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.D.; Iinishi, A.; Modaresi, S.M.; Yoo, B.-K.; Curtis, T.D.; Lariviere, P.J.; Liang, L.; Son, S.; Nicolau, S.; Bargabos, R.; et al. Computational identification of a systemic antibiotic for Gram-negative bacteria. Nat. Microbiol. 2022, 7, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Meyer, K.J.; Iinishi, A.; Favre-Godal, Q.; Green, R.; Manuse, S.; Caboni, M.; Mori, M.; Niles, S.; Ghiglieri, M.; et al. A new antibiotic selectively kills Gram-negative pathogens. Nature 2019, 576, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Jakob, R.P.; Marzinek, J.K.; Green, R.; Imai, Y.; Bolla, J.R.; Agustoni, E.; Robinson, C.V.; Bond, P.J.; Lewis, K.; et al. The antibiotic darobactin mimics a β-strand to inhibit outer membrane insertase. Nature 2021, 593, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.H.; Doyle, M.T.; Bernstein, H.D. Small Molecule Antibiotics Inhibit Distinct Stages of Bacterial Outer Membrane Protein Assembly. Mbio 2022, 13, e0228622. [Google Scholar] [CrossRef] [PubMed]
- Böhringer, N.; Green, R.; Liu, Y.; Mettal, U.; Marner, M.; Modaresi, S.M.; Jakob, R.P.; Wuisan, Z.G.; Maier, T.; Iinishi, A.; et al. Mutasynthetic Production and Antimicrobial Characterization of Darobactin Analogs. Microbiol. Spectr. 2021, 9, e0153521. [Google Scholar] [CrossRef]
- Groß, S.; Panter, F.; Pogorevc, D.; Seyfert, C.E.; Deckarm, S.; Bader, C.D.; Herrmann, J.; Müller, R. Improved broad-spectrum antibiotics against Gram-negative pathogens via darobactin biosynthetic pathway engineering. Chem. Sci. 2021, 12, 11882–11893. [Google Scholar] [CrossRef] [PubMed]
- Wuisan, Z.G.; Kresna, I.D.M.; Böhringer, N.; Lewis, K.; Schäberle, T.F. Optimization of heterologous Darobactin A expression and identification of the minimal biosynthetic gene cluster. Metab. Eng. 2021, 66, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Seyfert, C.E.; Porten, C.; Yuan, B.; Deckarm, S.; Panter, F.; Bader, C.D.; Coetzee, J.; Deschner, F.; Tehrani, K.H.; Higgins, P.G.; et al. Darobactins Exhibiting Superior Antibiotic Activity by Cryo-EM Structure Guided Biosynthetic Engineering. Angew. Chem. Int. Ed. 2022, 62, e202214094. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Schneider, F.; Eberle, K.J.; Chiodi, D.; Nakamura, H.; Reisberg, S.H.; Chen, J.; Saito, M.; Baran, P.S. Atroposelective Total Synthesis of Darobactin A. J. Am. Chem. Soc. 2022, 144, 14458–14462. [Google Scholar] [CrossRef]
- Srinivas, N.; Jetter, P.; Ueberbacher, B.J.; Werneburg, M.; Zerbe, K.; Steinmann, J.; Van der Meijden, B.; Bernardini, F.; Lederer, A.; Dias, R.L.A.; et al. Peptidomimetic Antibiotics Target Outer-Membrane Biogenesis in Pseudomonas aeruginosa. Science 2010, 327, 1010–1013. [Google Scholar] [CrossRef]
- Martin-Loeches, I.; Dale, G.E.; Torres, A. Murepavadin: A new antibiotic class in the pipeline. Expert Rev. Anti-infective Ther. 2018, 16, 259–268. [Google Scholar] [CrossRef]
- Díez-Aguilar, M.; Hernández-García, M.; Morosini, M.-I.; Fluit, A.; Tunney, M.M.; Huertas, N.; del Campo, R.; Obrecht, D.; Bernardini, F.; Ekkelenkamp, M.; et al. Murepavadin antimicrobial activity against and resistance development in cystic fibrosis Pseudomonas aeruginosa isolates. J. Antimicrob. Chemother. 2021, 76, 984–992. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, e00031-19. [Google Scholar] [CrossRef]
- Urfer, M.; Bogdanovic, J.; Monte, F.L.; Moehle, K.; Zerbe, K.; Omasits, U.; Ahrens, C.H.; Pessi, G.; Eberl, L.; Robinson, J.A. A Peptidomimetic Antibiotic Targets Outer Membrane Proteins and Disrupts Selectively the Outer Membrane in Escherichia coli. J. Biol. Chem. 2016, 291, 1921–1932. [Google Scholar] [CrossRef]
- Luther, A.; Urfer, M.; Zahn, M.; Müller, M.; Wang, S.-Y.; Mondal, M.; Vitale, A.; Hartmann, J.-B.; Sharpe, T.; Monte, F.L.; et al. Chimeric peptidomimetic antibiotics against Gram-negative bacteria. Nature 2019, 576, 452–458. [Google Scholar] [CrossRef]
- Ghequire, M.G.K.; Öztürk, B.; De Mot, R. Lectin-Like Bacteriocins. Front. Microbiol. 2018, 9, 2706. [Google Scholar] [CrossRef] [PubMed]
- Ghequire, M.G.K.; De Mot, R. LlpB represents a second subclass of lectin-like bacteriocins. Microb. Biotechnol. 2019, 12, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Hart, E.M.; Mitchell, A.M.; Konovalova, A.; Grabowicz, M.; Sheng, J.; Han, X.; Rodriguez-Rivera, F.P.; Schwaid, A.G.; Malinverni, J.C.; Balibar, C.J.; et al. A small-molecule inhibitor of BamA impervious to efflux and the outer membrane permeability barrier. Proc. Natl. Acad. Sci. USA 2019, 116, 21748–21757. [Google Scholar] [CrossRef] [PubMed]
- Wade, N.; Wesseling, C.M.J.; Innocenti, P.; Slingerland, C.J.; Koningstein, G.M.; Luirink, J.; Martin, N.I. Synthesis and Structure–Activity Studies of β-Barrel Assembly Machine Complex Inhibitor MRL-494. ACS Infect. Dis. 2022, 8, 2242–2252. [Google Scholar] [CrossRef] [PubMed]
- Storek, K.M.; Auerbach, M.R.; Shi, H.; Garcia, N.K.; Sun, D.; Nickerson, N.N.; Vij, R.; Lin, Z.; Chiang, N.; Schneider, K.; et al. Monoclonal antibody targeting the β-barrel assembly machine of Escherichia coli is bactericidal. Proc. Natl. Acad. Sci. USA 2018, 115, 3692–3697. [Google Scholar] [CrossRef]
- Storek, K.M.; Vij, R.; Sun, D.; Smith, P.A.; Koerber, J.T.; Rutherford, S.T. The Escherichia coli β-Barrel Assembly Machinery Is Sensitized to Perturbations under High Membrane Fluidity. J. Bacteriol. 2019, 201, e00517-18. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, X.; Zhang, J.; Lin, Y.; You, X.; Chen, M.; Wang, Y.; Zhu, N.; Si, S. Identification of a Compound That Inhibits the Growth of Gram-Negative Bacteria by Blocking BamA–BamD Interaction. Front. Microbiol. 2020, 11, 1252. [Google Scholar] [CrossRef]
- Mori, N.; Ishii, Y.; Tateda, K.; Kimura, S.; Kouyama, Y.; Inoko, H.; Mitsunaga, S.; Yamaguchi, K.; Yoshihara, E. A peptide based on homologous sequences of the β-barrel assembly machinery component BamD potentiates antibiotic susceptibility of Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2012, 67, 2173–2181. [Google Scholar] [CrossRef]
- Shamir, E.R.; Warthan, M.; Brown, S.P.; Nataro, J.P.; Guerrant, R.L.; Hoffman, P.S. Nitazoxanide Inhibits Biofilm Production and Hemagglutination by Enteroaggregative Escherichia coli Strains by Blocking Assembly of AafA Fimbriae. Antimicrob. Agents Chemother. 2010, 54, 1526–1533. [Google Scholar] [CrossRef]
- Chahales, P.; Hoffman, P.S.; Thanassi, D.G. Nitazoxanide Inhibits Pilus Biogenesis by Interfering with Folding of the Usher Protein in the Outer Membrane. Antimicrob. Agents Chemother. 2016, 60, 2028–2038. [Google Scholar] [CrossRef]
- Psonis, J.J.; Chahales, P.; Henderson, N.S.; Rigel, N.W.; Hoffman, P.S.; Thanassi, D.G. The small molecule nitazoxanide selectively disrupts BAM-mediated folding of the outer membrane usher protein. J. Biol. Chem. 2019, 294, 14357–14369. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.C.; Popat, R.; Diggle, S.P.; Brown, S.P. Targeting virulence: Can we make evolution-proof drugs? Nat. Rev. Microbiol. 2014, 12, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Steenhuis, M.; Abdallah, A.M.; De Munnik, S.M.; Kuhne, S.; Sterk, G.; van den Berg van Saparoea, B.; Westerhausen, S.; Wagner, S.; van der Wel, N.; Wijtmans, M.; et al. Inhibition of autotransporter biogenesis by small molecules. Mol. Microbiol. 2019, 112, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Ghequire, M.G.K.; Swings, T.; Michiels, J.; Buchanan, S.K.; De Mot, R. Hitting with a BAM: Selective Killing by Lectin-Like Bacteriocins. Mbio 2018, 9, e02138-17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Also Known As | Length (Amino Acids) | Mass (Da) | Identifier (Uniprot) |
|---|---|---|---|---|
| BamA | ecfK; ECK0176; yaeT; yzzN; yzzY | 810 | 90,553 | P0A940 |
| BamB | ECK2508; yfgL | 392 | 41,887 | P77774 |
| BamC | dapX; ECK2473; nlpB | 344 | 36,842 | P0A903 |
| BamD | ecfD; ECK2593; yfiO | 245 | 27,829 | P0AC02 |
| BamE | b2617, ECK2613, smpA, smqA | 113 | 12,302 | P0A937 |
| Inhibitors | Published Year | Molecular Target | Type | Reference |
|---|---|---|---|---|
| Darobactin A | 2019 | BamA (lateral gate) | peptide | [78] |
| Darobactin B | 2021 | BamA (lateral gate) | peptide | [81] |
| Darobactin 9 | 2021 | BamA (lateral gate) | peptide | [82] |
| Darobactin 22 | 2022 | BamA (lateral gate) | peptide | [84] |
| Dynobactin A | 2022 | BamA (lateral gate) | peptide | [77] |
| JB-95 | 2015 | BamA, LptD | peptide | [90] |
| BamA(extracellular loops 4, 6 and 7 of BamA) | ||||
| LlpAs | 2018 | BamA (extracellular loop 6) | peptide | [105] |
| MRL-494 | 2019 | BamA | small molecule | [94] |
| MAB1 | 2018 | BamA (extracellular loop 4) | monoclonal antibody | [96] |
| IMB-H4 | 2020 | BamA | small molecule | [98] |
| FIRL | 2012 | BamA | peptide | [99] |
| BamD Inhibitory Peptide | 2015 | BamD | peptide | [53] |
| NTZ | 2019 | BAM complex | small molecule | [102] |
| VUF15259 | 2019 | BAM complex | small molecule | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Guo, M.; Yu, F. β-Barrel Assembly Machinery (BAM) Complex as Novel Antibacterial Drug Target. Molecules 2023, 28, 3758. https://doi.org/10.3390/molecules28093758
Xu Q, Guo M, Yu F. β-Barrel Assembly Machinery (BAM) Complex as Novel Antibacterial Drug Target. Molecules. 2023; 28(9):3758. https://doi.org/10.3390/molecules28093758
Chicago/Turabian StyleXu, Qian, Min Guo, and Feiyuan Yu. 2023. "β-Barrel Assembly Machinery (BAM) Complex as Novel Antibacterial Drug Target" Molecules 28, no. 9: 3758. https://doi.org/10.3390/molecules28093758