
Innovative Metrics for Reporting and Comparing the Glycan Structural Profile in Biotherapeutics

Abstract
:1. Introduction
2. Main N-Glycan-Related CQAs and Indices
2.1. Galactose-α1,3-Galactose (αGal)
2.2. N-Glycolylneuraminic Acids (Neu5Gc)
2.3. High Mannose (HM) Glycoforms
2.4. Hybrid Type
2.5. Antennarity
2.6. Sialylation
2.7. Sialic Acid O-Acetylation
2.8. Bisecting N-Acetylglucosamine (Bisecting GlcNac)
2.9. N-Acetyllactosamine (LacNAc)
2.10. Fucosylation
2.10.1. Core α1,6 Fucosylation
2.10.2. Antennae Fucosylation
2.11. Galactosylation
2.11.1. Non-Fc Glycans
2.11.2. Fc-Glycans
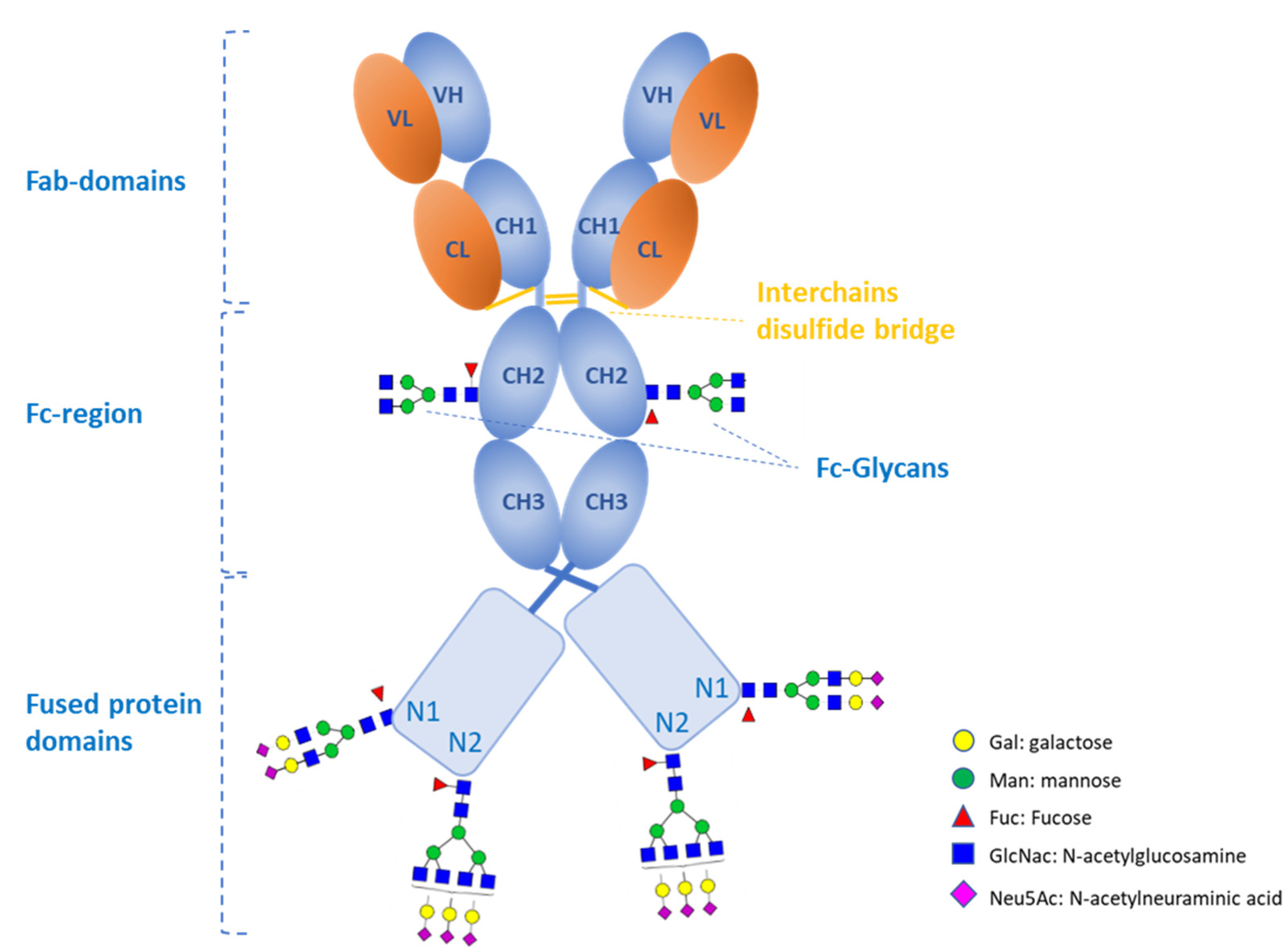
2.12. Site Occupancy
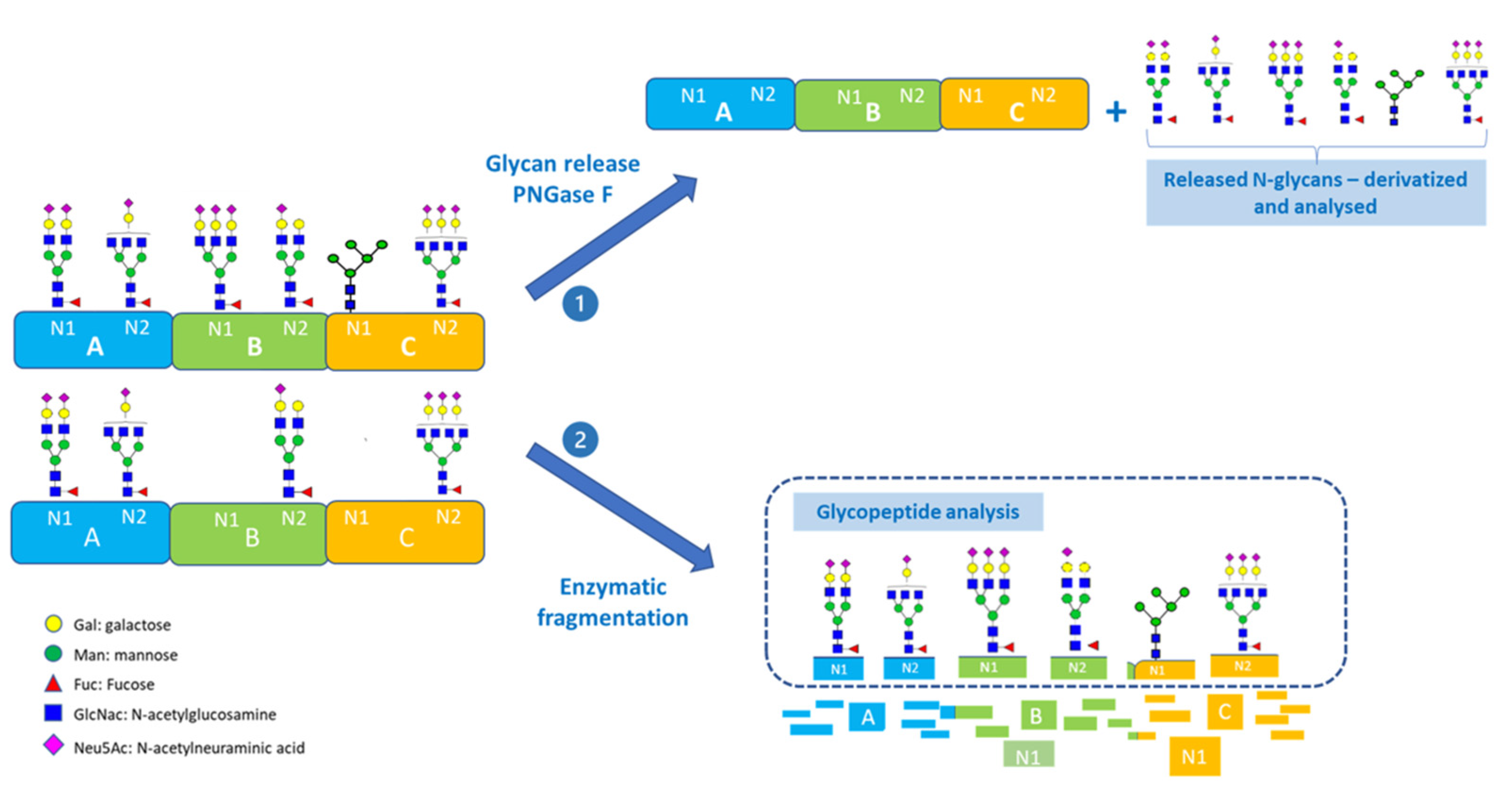
3. Analytical Workflow and Case Studies
3.1. Mass Spectrometry-Based Glycopeptide Analysis
3.2. Matrix of Glycan-Related CQAs
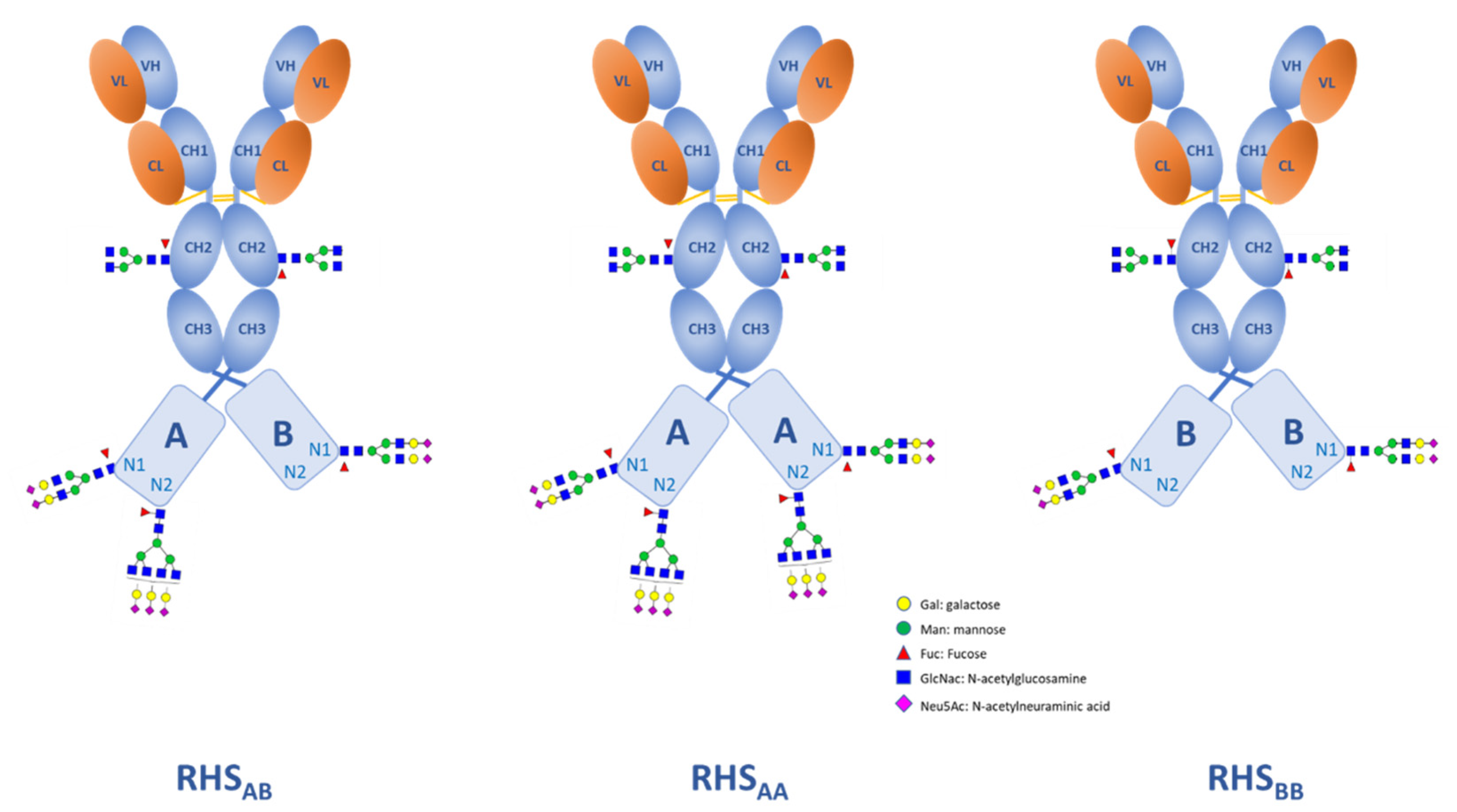
3.3. Case Study 1: Antibody Fusion Protein Expressed by CHO Cells
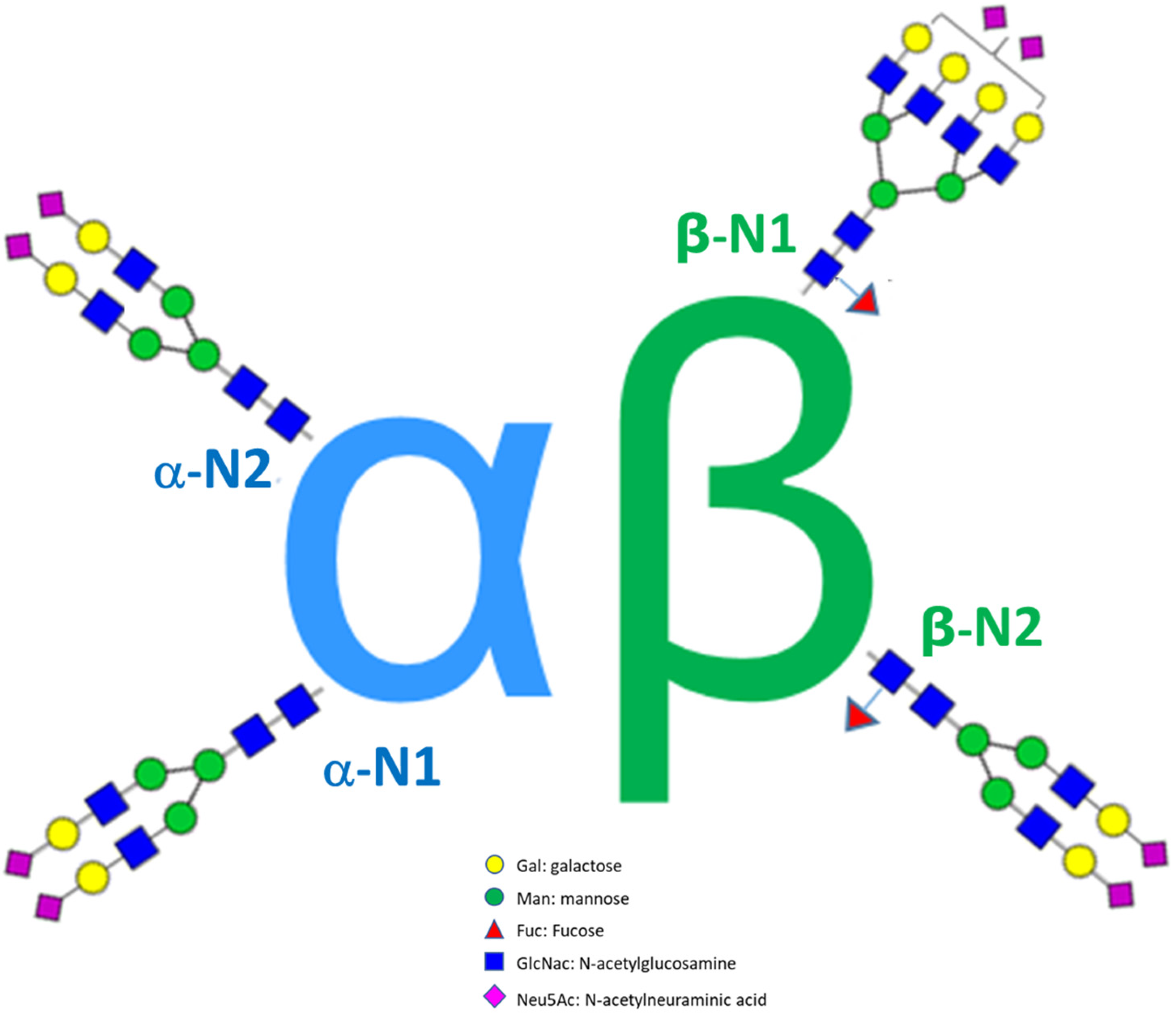
3.4. Case Study 2: Comparability Study of Three Recombinant FSH Products
4. Discussion
4.1. Need for Standardization
4.2. Risk Assessment
4.3. Adaptability and Versatility
5. Conclusions
6. Materials and Methods
6.1. Materials
6.2. Methods
6.2.1. Glycopeptide Mapping
6.2.2. Site Occupancy Analysis
6.2.3. Index Calculation
Site-Specific Indices
- Sialylation index (SI)
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- Neu5Gc Sialylation Index (SINeu5Gc)
- AnGmSy: non-fucosylated glycoforms with n antennae, m galactose residues, and y Neu5Gc residues present.
- FAnGmSy: fucosylated glycoforms with n antennae, m galactose residues, and y Neu5Gc residues present.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- y can range from 0 to m.
- Sialylation Extent (SE)
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- SIα2,6
- AnGmSy: non-fucosylated glycoforms with n antennae, m galactose residues, and y sialic acid residues in α2,6 linkage present.
- FAnGmSy: fucosylated glycoforms with n antennae, m galactose residues, and y sialic acid residues in α2,6 linkage present.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- y can range from 0 to m.
- Neu5Gc %
- AnGmSy: non-fucosylated glycoforms with n antennae, m galactose residues, and y Neu5Gc residues present.
- FAnGmSy: fucosylated glycoforms with n antennae, m galactose residues, and y Neu5Gc residues present.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- y can range from 0 to m.
- O-acetylated %
- AnGmSy: non-fucosylated glycoforms with n antennae, m galactose residues, and y O-acetylated sialic acid percent residues present.
- FAnGmSy: fucosylated glycoforms with n antennae, m galactose residues, and y O-acetylated sialic acid percent residues present.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- y can range from 0 to m.
- A-index (AI)
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- Site occupancy index (SOI)
- Core Fucosylation index (cFI)
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present;
- n can range from 1 to 5;
- m can range from 0 to n;
- x can range from 0 to m.
- Antennae fucose Index (aFI)
- yaF: number of fucose residues present in the antennae;
- aFAnGmSx: antennae fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present;
- n can range from 1 to 5;
- m can range from 0 to n;
- x can range from 0 to m.
- Galactosylation Index (GI)
- m: number of galactose residues present in the glycan.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present:
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- G0 (only for molecule containing the Fc domain)
- AnG0: non-fucosylated glycoforms with n antennae and 0 galactose residues;
- FAnG0: fucosylated glycoforms with n antennae and 0 galactose residues.
- G11,6 (only for molecules containing the Fc domain)
- AnG11,6: non-fucosylated glycoforms with n antennae and 1 galactose residue on the α1-6 arm:
- FAnG11,6: fucosylated glycoforms with n antennae and 1 galactose residue on the α1,6 arm;
- Note: the distinction between G11,6 and G11,3 is only possible with dedicated analytical techniques.
- G2 (only for molecules containing the Fc domain)
- AnG2: non-fucosylated glycoforms with n antennae and 2 galactose residues.
- FAnG2: fucosylated glycoforms with n antennae and 2 galactose residues.
- n can range from 0 to 2.
- α Gal Index (αGI)
- yαG: number of galactose residues in alpha-1-3 linkage.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- N-acetyllactosamine (LacNAc) Index (LI)
- yLacNAc: number of N-acetyllactosamine (LacNAc) units present in the glycan.
- AnGmSx: non-fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- FAnGmSx: fucosylated glycoforms with n antennae, m galactose residues, and x sialic acid residues (any type or linkage) present.
- n can range from 1 to 5.
- m can range from 0 to n.
- x can range from 0 to m.
- Mannose Index (MI)
- m: number of mannose residues present in the high mannose structures (M5–M9).
- m can range from 5 to 9;
- Note: the MI only takes into consideration the high mannose structures (M5–M9) in the calculation.
- Hybrid Index (HI)
- Bisecting Index (BI)
Calculation of the Indices for the Whole Molecule/Domains
7. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RHS Characterization Matrix | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | % Neu5Gc | % O-acetyl | GI | MI | cFI | |
| Fc | Mean | 1.0 | 1.74 | ND | ND | ND | ND | ND | 0.24 | 0.02 | 1.00 |
| CV% | - (1) | 0.36 | NA | NA | NA | NA | NA | 1.73 | 5.42 | 0.15 | |
| StDev | - (1) | 0.01 | NA | NA | NA | NA | NA | 0.00 | 0.00 | 0.00 | |
| N1 | Mean | 1.0 | 2.64 | 1.72 | 0.64 | 0.01 | 0.46 | 3.00 | 0.64 | 0.01 | 1.00 |
| CV% | - (1) | 0.93 | 1.20 | 0.66 | 9.16 | 9.37 | 7.16 | 2.33 | 10.69 | 0.21 | |
| StDev | - (1) | 0.02 | 0.02 | 0.00 | 0.00 | 0.04 | 0.21 | 0.01 | 0.00 | 0.00 | |
| N2 | Mean | 0.6 | 3.43 | 2.44 | 0.71 | 0.01 | 0.22 | 2.18 | 0.79 | ND | 1.00 |
| CV% | - (1) | 0.28 | 0.54 | 0.41 | 12.34 | 12.41 | 6.93 | 1.10 | NA | 0.12 | |
| StDev | - (1) | 0.01 | 0.01 | 0.00 | 0.00 | 0.03 | 0.15 | 0.01 | NA | 0.00 | |
| whole molecule/domains | 9.34 * | 6.33 * | 0.68 * | 0.03 * | 0.05 ** | ||||||
| Product A | Product B | Product C | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Index | Glycosite | Batch 1 | Batch 2 | Batch 3 | Batch 1 | Batch 2 | Batch 1 | Batch 2 | Batch 3 |
| AI | α-N1 | 2.12 | 2.11 | 2.12 | 2.25 | 2.29 | 2.08 | 2.11 | 2.11 |
| α-N2 | 2.08 | 2.07 | 2.10 | 2.23 | 2.25 | 2.07 | 2.08 | 2.08 | |
| β-N1 | 3.60 | 3.46 | 3.56 | 4.15 | 4.12 | 3.79 | 3.85 | 3.66 | |
| β-N2 | 2.15 | 2.18 | 2.16 | 2.24 | 2.25 | 2.20 | 2.18 | 2.27 | |
| Whole | 9.95 | 9.82 | 9.94 | 10.87 | 10.91 | 10.14 | 10.22 | 10.12 | |
| SI | α-N1 | 1.75 | 1.74 | 1.76 | 1.78 | 1.79 | 1.81 | 1.84 | 1.79 |
| α-N2 | 1.76 | 1.76 | 1.78 | 1.77 | 1.78 | 1.85 | 1.87 | 1.86 | |
| β-N1 | 2.11 | 2.35 | 2.29 | 2.65 | 2.74 | 2.17 | 2.18 | 2.22 | |
| β-N2 | 1.86 | 1.88 | 1.88 | 1.95 | 1.94 | 1.98 | 2.02 | 1.97 | |
| Whole | 7.48 | 7.73 | 7.71 | 8.15 | 8.25 | 7.81 | 7.91 | 7.84 | |
| SE | α-N1 | 0.83 | 0.82 | 0.82 | 0.80 | 0.79 | 0.87 | 0.87 | 0.85 |
| α-N2 | 0.85 | 0.85 | 0.86 | 0.81 | 0.81 | 0.90 | 0.91 | 0.90 | |
| β-N1 | 0.64 | 0.73 | 0.69 | 0.68 | 0.70 | 0.63 | 0.62 | 0.66 | |
| β-N2 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.92 | 0.94 | 0.90 | |
| Whole | 0.75 | 0.79 | 0.78 | 0.75 | 0.76 | 0.77 | 0.77 | 0.77 | |
| SINeu5Gc | α-N1 | 0.01 | 0.01 | 0.00 | 0.00 | 0.00 | 0.07 | 0.07 | 0.08 |
| α-N2 | 0.00 | 0.00 | 0.00 | 0.01 | 0.01 | 0.09 | 0.07 | 0.08 | |
| β-N1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| β-N2 | 0.00 | 0.01 | 0.00 | 0.01 | 0.01 | 0.07 | 0.08 | 0.07 | |
| Whole | 0.01 | 0.01 | 0.00 | 0.01 | 0.01 | 0.22 | 0.22 | 0.24 | |
| %Neu5Gc | α-N1 | 0.29 | 0.35 | 0.00 | 0.23 | 0.11 | 3.87 | 3.97 | 4.71 |
| α-N2 | 0.00 | 0.00 | 0.00 | 0.27 | 0.27 | 4.57 | 3.93 | 4.43 | |
| β-N1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| β-N2 | 0.00 | 0.43 | 0.11 | 0.26 | 0.25 | 3.48 | 3.69 | 3.62 | |
| %O-acetyl | α-N1 | 2.40 | 2.59 | 2.39 | 0.00 | 0.00 | 2.21 | 2.01 | 1.12 |
| α-N2 | 5.51 | 4.73 | 4.54 | 0.38 | 0.32 | 5.59 | 6.54 | 5.23 | |
| β-N1 | 2.51 | 2.12 | 1.40 | 0.34 | 0.18 | 1.66 | 3.03 | 1.39 | |
| β-N2 | 7.58 | 6.24 | 7.12 | 1.28 | 0.81 | 6.05 | 7.00 | 5.71 | |
| GI | α-N1 | 0.36 | 0.37 | 0.37 | 0.47 | 0.50 | 0.26 | 0.27 | 0.31 |
| α-N2 | 0.32 | 0.32 | 0.31 | 0.46 | 0.47 | 0.22 | 0.20 | 0.22 | |
| β-N1 | 1.68 | 1.20 | 1.41 | 1.27 | 1.16 | 1.79 | 1.80 | 1.61 | |
| β-N2 | 0.22 | 0.20 | 0.20 | 0.20 | 0.21 | 0.10 | 0.11 | 0.12 | |
| MI | α-N1 | ND | ND | ND | ND | ND | ND | ND | ND |
| α-N2 | ND | ND | ND | ND | ND | ND | ND | ND | |
| β-N1 | ND | ND | ND | ND | ND | ND | ND | ND | |
| β-N2 | ND | ND | ND | ND | ND | ND | ND | ND | |
| Whole | ND | ND | ND | ND | ND | ND | ND | ND | |
| cFI | α-N1 | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| α-N2 | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | |
| β-N1 | 0.51 | 0.63 | 0.56 | 0.53 | 0.56 | 0.48 | 0.48 | 0.51 | |
| β-N2 | 1.02 | 1.02 | 1.02 | 1.01 | 1.01 | 0.98 | 0.99 | 1.00 | |
| LI | α-N1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| α-N2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| β-N1 | 0.37 | 0.26 | 0.32 | 0.39 | 0.37 | 0.42 | 0.44 | 0.36 | |
| β-N2 | 0.00 | 0.00 | 0.00 | 0.01 | 0.01 | 0.00 | 0.00 | 0.00 | |
| Whole | 0.37 | 0.26 | 0.32 | 0.40 | 0.38 | 0.42 | 0.44 | 0.36 | |
References
- Bavbek, S.; Pagani, M.; Alvarez-Cuesta, E.; Castells, M.; Dursun, A.B.; Hamadi, S.; Madrigal-Burgaleta, R.; Sanchez-Sanchez, S.; Vultaggio, A. Hypersensitivity reactions to biologicals: An EAACI position paper. Allergy 2021, 77, 39–54. [Google Scholar] [CrossRef]
- Gulsen, A.; Wedi, B.; Jappe, U. Hypersensitivity reactions to biologics (part I): Allergy as an important differential diagnosis in complex immune-derived adverse events. Allergo J. Int. 2020, 29, 97–125. [Google Scholar] [CrossRef]
- Berg, E.A.; Platts-Mills, T.; Commins, S. Drug allergens and food--the cetuximab and galactose-alpha-1,3-galactose story. Ann. Allergy Asthma Immunol. 2014, 112, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Commins, S.P.; Kelly, L.A.; Rönmark, E.; James, H.R.; Pochan, S.L.; Peters, E.J.; Lundbäck, B.; Nganga, L.W.; Cooper, P.J.; Hoskins, J.M.; et al. Galactose-alpha-1,3-galactose-specific IgE is associated with anaphylaxis but not asthma. Am. J. Respir. Crit. Care Med. 2012, 185, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, C.H.; Mirakhur, B.; Chan, E.; Le, Q.-T.; Berlin, J.; Morse, M.; Murphy, B.A.; Satinover, S.M.; Hosen, J.; Mauro, D.; et al. Cetuximab-Induced Anaphylaxis and IgE Specific for Galactose-α-1,3-Galactose. N. Engl. J. Med. 2008, 358, 1109–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinke, J.W.; Platts-Mills, T.A.; Commins, S.P. The alpha-gal story: Lessons learned from connecting the dots. J. Allergy Clin. Immunol. 2015, 135, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Mastrangeli, R.; Audino, M.C.; Palinsky, W.; Broly, H.; Bierau, H. The Formidable Challenge of Controlling High Mannose-Type N-Glycans in Therapeutic mAbs. Trends Biotechnol. 2020, 38, 1154–1168. [Google Scholar] [CrossRef]
- Dong, X.; Storkus, W.; Salter, R.D. Binding and uptake of agalactosyl IgG by mannose receptor on macrophages and dendritic cells. J. Immunol. 1999, 163, 5427–5434. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.B.; Ng, S.K. Impact of host cell line choice on glycan profile. Crit. Rev. Biotechnol. 2017, 38, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Blondeel, E.J.; Aucoin, M.G. Supplementing glycosylation: A review of applying nucleotide-sugar precursors to growth medium to affect therapeutic recombinant protein glycoform distributions. Biotechnol. Adv. 2018, 36, 1505–1523. [Google Scholar] [CrossRef] [Green Version]
- Mastrangeli, R.; Palinsky, W.; Bierau, H. Glycoengineered antibodies: Towards the next-generation of immunotherapeutics. Glycobiology 2018, 29, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.; Grunert, I.; von Roman, M.F.; Walch, H.; Dams, T.; Thomann, M.; Falkenstein, R. Implementation of in vitro glycoengineering of monoclonal antibodies into downstream processing of industrial production. Glycobiology 2021, 32, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Reusch, D.; Tejada, M.L. Fc glycans of therapeutic antibodies as critical quality attributes. Glycobiology 2015, 25, 1325–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimura, Y.; Saldova, R.; Mimura-Kimura, Y.; Rudd, P.M.; Jefferis, R. Importance and Monitoring of Therapeutic Immunoglobulin G Glycosylation. In Antibody Glycosylation; Pezer, M., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 481–517. [Google Scholar]
- De Leoz, M.L.; Duewer, D.L.; Fung, A.; Liu, L.; Yau, H.K.; Potter, O.; Staples, G.O.; Furuki, K.; Frenkel, R.; Hu, Y.; et al. NIST Interlaboratory Study on Glycosylation Analysis of Monoclonal Antibodies: Comparison of Results from Diverse Analytical Methods. Mol. Cell Proteom. 2020, 19, 11–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everest-Dass, A.V.; Moh, E.S.X.; Ashwood, C.; Shathili, A.M.M.; Packer, N.H. Human disease glycomics: Technology advances enabling protein glycosylation analysis—Part 1. Expert Rev. Proteom. 2018, 15, 165–182. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Zauner, G.; Huhn, C.; Bruggink, C.; Deelder, A.M.; Wuhrer, M. Glycan labeling strategies and their use in identification and quantification. Anal. Bioanal. Chem. 2010, 397, 3457–3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faid, V.; Denguir, N.; Chapuis, V.; Bihoreau, N.; Chevreux, G. Site-specific N-glycosylation analysis of human factor XI: Identification of a noncanonical NXC glycosite. Proteomics 2014, 14, 2460–2470. [Google Scholar] [CrossRef] [PubMed]
- Lowenthal, M.S.; Davis, K.S.; Formolo, T.; Kilpatrick, L.E.; Phinney, K.W. Identification of Novel N-Glycosylation Sites at Noncanonical Protein Consensus Motifs. J. Proteome Res. 2016, 15, 2087–2101. [Google Scholar] [CrossRef] [Green Version]
- Zhen, Y.; Caprioli, R.M.; Staros, J.V. Characterization of Glycosylation Sites of the Epidermal Growth Factor Receptor. Biochemistry 2003, 42, 5478–5492. [Google Scholar] [CrossRef] [Green Version]
- Valliere-Douglass, J.F.; Kodama, P.; Mujacic, M.; Brady, L.J.; Wang, W.; Wallace, A.; Yan, B.; Reddy, P.; Treuheit, M.J.; Balland, A. Asparagine-linked Oligosaccharides Present on a Non-consensus Amino Acid Sequence in the CH1 Domain of Human Antibodies. J. Biol. Chem. 2009, 284, 32493–32506. [Google Scholar] [CrossRef] [Green Version]
- Valliere-Douglass, J.F.; Eakin, C.M.; Wallace, A.; Ketchem, R.R.; Wang, W.; Treuheit, M.J.; Balland, A. Glutamine-linked and Non-consensus Asparagine-linked Oligosaccharides Present in Human Recombinant Antibodies Define Novel Protein Glycosylation Motifs. J. Biol. Chem. 2010, 285, 16012–16022. [Google Scholar] [CrossRef] [Green Version]
- Mariño, K.; Bones, J.; Kattla, J.J.; Rudd, P.M. A systematic approach to protein glycosylation analysis: A path through the maze. Nat. Chem. Biol. 2010, 6, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Spearman, M.; Doering, J.; Lattová, E.; Perreault, H.; Butler, M. The availability of glucose to CHO cells affects the intracellular lipid-linked oligosaccharide distribution, site occupancy and the N-glycosylation profile of a monoclonal antibody. J. Biotechnol. 2013, 170, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermentin, P.; Witzel, R.; Kanzy, E.-J.; Diderrich, G.; Hoffmann, D.; Metzner, H.; Vorlop, J.; Haupt, H. The hypothetical N-glycan charge: A number that characterizes protein glycosylation. Glycobiology 1996, 6, 217–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervais, A.; Hammel, Y.-A.; Pelloux, S.; Lepage, P.; Baer, G.; Carte, N.; Sorokine, O.; Strub, J.-M.; Koerner, R.; Leize, E.; et al. Glycosylation of human recombinant gonadotrophins: Characterization and batch-to-batch consistency. Glycobiology 2002, 13, 179–189. [Google Scholar] [CrossRef]
- Park, E.I.; Mi, Y.; Unverzagt, C.; Gabius, H.-J.; Baenziger, J.U. The asialoglycoprotein receptor clears glycoconjugates terminating with sialic acidα2,6GalNAc. Proc. Natl. Acad. Sci. USA 2005, 102, 17125–17129. [Google Scholar] [CrossRef] [Green Version]
- Platts-Mills, T.A.; Commins, S.P.; Biedermann, T.; van Hage, M.; Levin, M.; Beck, L.A.; Diuk-Wasser, M.; Jappe, U.; Apostolovic, D.; Minnicozzi, M.; et al. On the cause and consequences of IgE to galactose-α-1,3-galactose: A report from the National Institute of Allergy and Infectious Diseases Workshop on Understanding IgE-Mediated Mammalian Meat Allergy. J. Allergy Clin. Immunol. 2020, 145, 1061–1071. [Google Scholar] [CrossRef]
- Dupont, B.; Mariotte, D.; Moldovan, C.; Grellard, J.-M.; Vergnaud, M.-C.; Laroche, D.; Gervais, R. Case Report about Fatal or Near-Fatal Hypersensitivity Reactions to Cetuximab: Anticetuximab IgE as a Valuable Screening Test. Clin. Med. Insights Oncol. 2014, 8, 91–94. [Google Scholar] [CrossRef]
- Commins, S.P.; James, H.R.; Kelly, L.A.; Pochan, S.L.; Workman, L.J.; Perzanowski, M.S.; Kocan, K.M.; Fahy, J.V.; Nganga, L.W.; Ronmark, E.; et al. The relevance of tick bites to the production of IgE antibodies to the mammalian oligosaccharide galactose-α-1,3-galactose. J. Allergy Clin. Immunol. 2011, 127, 1286–1293.e6. [Google Scholar] [CrossRef] [Green Version]
- Commins, S.P.; Satinover, S.M.; Hosen, J.; Mozena, J.; Borish, L.; Lewis, B.D.; Woodfolk, J.A.; Platts-Mills, T.A. Delayed anaphylaxis, angioedema, or urticaria after consumption of red meat in patients with IgE antibodies specific for galactose-alpha-1,3-galactose. J. Allergy Clin. Immunol. 2009, 123, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Koike, C.; Fung, J.J.; Geller, D.A.; Kannagi, R.; Libert, T.; Luppi, P.; Nakashima, I.; Profozich, J.; Rudert, W.; Sharma, S.B.; et al. Molecular basis of evolutionary loss of the alpha 1,3-galactosyltransferase gene in higher primates. J. Biol. Chem. 2002, 277, 10114–10120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tector, A.J.; Mosser, M.; Tector, M.; Bach, J.-M. The Possible Role of Anti-Neu5Gc as an Obstacle in Xenotransplantation. Front. Immunol. 2020, 11, 622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, Y.; Chen, L.; Wang, R.; Li, C.; Li, X.; Fang, B.; Ren, X.; Ruan, M.; Liu, J.; et al. Reducing immunoreactivity of porcine bioprosthetic heart valves by genetically-deleting three major glycan antigens, GGTA1/β4GalNT2/CMAH. Acta Biomater. 2018, 72, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Van Bueren, J.J.; Rispens, T.; Verploegen, S.; van der Palen-Merkus, T.; Stapel, S.; Workman, L.J.; James, H.; van Berkel, P.H.; van de Winkel, J.G.; Platts-Mills, T.A.; et al. Anti-galactose-α-1,3-galactose IgE from allergic patients does not bind α-galactosylated glycans on intact therapeutic antibody Fc domains. Nat. Biotechnol. 2011, 29, 574–576. [Google Scholar] [CrossRef]
- Larsen, R.D.; Rajan, V.P.; Ruff, M.M.; Kukowska-Latallo, J.; Cummings, R.D.; Lowe, J.B. Isolation of a cDNA encoding a murine UDPgalactose:beta-D-galactosyl- 1,4-N-acetyl-D-glucosaminide alpha-1,3-galactosyltransferase: Expression cloning by gene transfer. Proc. Natl. Acad. Sci. USA 1989, 86, 8227–8231. [Google Scholar] [CrossRef] [Green Version]
- Bosques, C.J.; Collins, B.E.; Meador, I.I.I.J.W.; Sarvaiya, H.; Murphy, J.L.; DelloRusso, G.; Bulik, D.A.; Hsu, I.H.; Washburn, N.; Sipsey, S.F.; et al. Chinese hamster ovary cells can produce galactose-α-1,3-galactose antigens on proteins. Nat. Biotechnol. 2010, 28, 1153–1156. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, N.; Parekh, R.B.; James, D.C. Getting the glycosylation right: Implications for the biotechnology industry. Nat. Biotechnol. 1996, 14, 975–981. [Google Scholar] [CrossRef]
- Taylor, R.E.; Gregg, C.J.; Padler-Karavani, V.; Ghaderi, D.; Yu, H.; Huang, S.; Sorensen, R.U.; Chen, X.; Inostroza, J.; Nizet, V.; et al. Novel mechanism for the generation of human xeno-autoantibodies against the nonhuman sialic acid N-glycolylneuraminic acid. J. Exp. Med. 2010, 207, 1637–1646. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi, D.; Taylor, R.E.; Padler-Karavani, V.; Diaz, S.; Varki, A. Implications of the presence of N-glycolylneuraminic acid in recombinant therapeutic glycoproteins. Nat. Biotechnol. 2010, 28, 863–867. [Google Scholar] [CrossRef] [Green Version]
- Yehuda, S.; Padler-Karavani, V. Glycosylated Biotherapeutics: Immunological Effects of N-Glycolylneuraminic Acid. Front. Immunol. 2020, 11, 21. [Google Scholar] [CrossRef]
- Breimer, M.E.; Holgersson, J. The Structural Complexity and Animal Tissue Distribution of N-Glycolylneuraminic Acid (Neu5Gc)-Terminated Glycans. Implications for Their Immunogenicity in Clinical Xenografting. Front. Mol. Biosci. 2019, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Mastrangeli, R.; Audino, M.C.; Palinsky, W.; Broly, H.; Bierau, H. Current views on N-glycolylneuraminic acid in therapeutic recombinant proteins. Trends Pharmacol. Sci. 2021, 42, 943–956. [Google Scholar] [CrossRef]
- Yu, C.; Gao, K.; Zhu, L.; Wang, W.; Wang, L.; Zhang, F.; Liu, C.; Li, M.; Wormald, M.R.; Rudd, P.M.; et al. Erratum: At least two Fc Neu5Gc residues of monoclonal antibodies are required for binding to anti-Neu5Gc antibody. Sci. Rep. 2017, 7, 46385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karst, D.J.; Scibona, E.; Serra, E.; Bielser, J.-M.; Souquet, J.; Stettler, M.; Broly, H.; Soos, M.; Morbidelli, M.; Villiger, T.K. Modulation and modeling of monoclonal antibody N-linked glycosylation in mammalian cell perfusion reactors. Biotechnol. Bioeng. 2017, 114, 1978–1990. [Google Scholar] [CrossRef]
- Sumit, M.; Dolatshahi, S.; Chu, A.H.; Cote, K.; Scarcelli, J.J.; Marshall, J.K.; Cornell, R.J.; Weiss, R.; Lauffenburger, D.A.; Mulukutla, B.C.; et al. Dissecting N-Glycosylation Dynamics in Chinese Hamster Ovary Cells Fed-batch Cultures using Time Course Omics Analyses. iScience 2019, 12, 102–120. [Google Scholar] [CrossRef] [Green Version]
- Sakae, Y.; Satoh, T.; Yagi, H.; Yanaka, S.; Yamaguchi, T.; Isoda, Y.; Iida, S.; Okamoto, Y.; Kato, K. Conformational effects of N-glycan core fucosylation of immunoglobulin G Fc region on its interaction with Fcgamma receptor IIIa. Sci. Rep. 2017, 7, 13780. [Google Scholar] [CrossRef] [Green Version]
- Shibata-Koyama, M.; Iida, S.; Misaka, H.; Mori, K.; Yano, K.; Shitara, K.; Satoh, M. Nonfucosylated rituximab potentiates human neutrophil phagocytosis through its high binding for FcgammaRIIIb and MHC class II expression on the phagocytotic neutrophils. Exp. Hematol. 2009, 37, 309–321. [Google Scholar] [CrossRef]
- Suzuki, E.; Niwa, R.; Saji, S.; Muta, M.; Hirose, M.; Iida, S.; Shiotsu, Y.; Satoh, M.; Shitara, K.; Kondo, M.; et al. A Nonfucosylated Anti-HER2 Antibody Augments Antibody-Dependent Cellular Cytotoxicity in Breast Cancer Patients. Clin. Cancer Res. 2007, 13, 1875–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raju, T.S. Terminal sugars of Fc glycans influence antibody effector functions of IgGs. Curr. Opin. Immunol. 2008, 20, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Shankara, S.; Roy, A.; Qiu, H.; Estes, S.; McVie-Wylie, A.; Culm-Merdek, K.; Park, A.; Pan, C.; Edmunds, T. Development of a simple and rapid method for producing non-fucosylated oligomannose containing antibodies with increased effector function. Biotechnol. Bioeng. 2007, 99, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Gorovits, B.; Krinos-Fiorotti, C. Proposed mechanism of off-target toxicity for antibody–drug conjugates driven by mannose receptor uptake. Cancer Immunol. Immunother. 2012, 62, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Mahalingaiah, P.K.; Ciurlionis, R.; Durbin, K.R.; Yeager, R.L.; Philip, B.K.; Bawa, B.; Mantena, S.R.; Enright, B.P.; Liguori, M.J.; Van Vleet, T.R. Potential mechanisms of target-independent uptake and toxicity of antibody-drug conjugates. Pharmacol. Ther. 2019, 200, 110–125. [Google Scholar] [CrossRef]
- Goetze, A.M.; Liu, Y.D.; Zhang, Z.; Shah, B.; Lee, E.; Bondarenko, P.V.; Flynn, G.C. High-mannose glycans on the Fc region of therapeutic IgG antibodies increase serum clearance in humans. Glycobiology 2011, 21, 949–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizuka, Y.; Taniguchi, N. Enzymes for N-Glycan Branching and Their Genetic and Nongenetic Regulation in Cancer. Biomolecules 2016, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrangeli, R.; Rossi, M.; Mascia, M.; Palinsky, W.; Datola, A.; Terlizzese, M.; Bierau, H. In vitro biological characterization of IFN-beta-1a major glycoforms. Glycobiology 2015, 25, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Cai, H.; Hu, Z.; Boswell, C.A.; Diao, J.; Li, C.; Zhang, L.; Shen, A.; Teske, C.A.; Zhang, B.; et al. Balancing the Affinity and Pharmacokinetics of Antibodies by Modulating the Size of Charge Patches on Complementarity-Determining Regions. J. Pharm. Sci. 2020, 109, 3690–3696. [Google Scholar] [CrossRef]
- D’Antonio, M.; Borrelli, F.; Datola, A.; Bucci, R.; Mascia, M.; Polletta, P.; Piscitelli, D.; Papoian, R. Biological characterization of recombinant human follicle stimulating hormone isoforms. Hum. Reprod. 1999, 14, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Egrie, J.C.; Dwyer, E.; Browne, J.K.; Hitz, A.; Lykos, M.A. Darbepoetin alfa has a longer circulating half-life and greater in vivo potency than recombinant human erythropoietin. Exp. Hematol. 2003, 31, 290–299. [Google Scholar] [CrossRef]
- Mulders, J.; Derksen, M.; Swolfs, A.; Maris, F. Prediction of the in vivo Biological Activity of Human Recombinant Follicle Stimulating Hormone Using Quantitative Isoelectric Focusing. Biologicals 1997, 25, 269–281. [Google Scholar] [CrossRef]
- Perlman, S.; Hazel, B.V.D.; Christiansen, J.; Gram-Nielsen, S.; Jeppesen, C.B.; Andersen, K.V.; Halkier, T.; Okkels, S.; Schambye, H.T. Glycosylation of an N-Terminal Extension Prolongs the Half-Life and Increases the in Vivo Activity of Follicle Stimulating Hormone. J. Clin. Endocrinol. Metab. 2003, 88, 3227–3235. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Li, H.; Zhang, Q.; Liu, X.; Zheng, Q.; Li, J. Characterization of O-acetylation in sialoglycans by MALDI-MS using a combination of methylamidation and permethylation. Sci. Rep. 2017, 7, 46206. [Google Scholar] [CrossRef] [Green Version]
- Schauer, R. Sialic acids: Fascinating sugars in higher animals and man. Zoology 2004, 107, 49–64. [Google Scholar] [CrossRef]
- Lewis, A.L.; Nizet, V.; Varki, A. Discovery and characterization of sialic acid O-acetylation in group B Streptococcus. Proc. Natl. Acad. Sci. USA 2004, 101, 11123–11128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.J.; Oh, H.; Choi, I.; Lee, K.; Shin, H.; Lee, Y.J.; Park, J.; Yang, Y.H.; Ha, G.-S.; Lee, D.-E.; et al. Differences in Proportion of N-acetyllactosamine and O-acetylated Sialic Acid Have No Significant Effect on the Pharmacokinetics and Biological Activity of Darbepoetin Alfa. Biotechnol. Bioprocess Eng. 2022, 27, 244–252. [Google Scholar] [CrossRef]
- Chen, Q.; Tan, Z.; Guan, F.; Ren, Y. The Essential Functions and Detection of Bisecting GlcNAc in Cell Biology. Front. Chem. 2020, 8, 511. [Google Scholar] [CrossRef] [PubMed]
- Cameron, F.; McCormack, P.L. Obinutuzumab: First Global Approval. Drugs 2013, 74, 147–154. [Google Scholar] [CrossRef]
- Nagae, M.; Yamanaka, K.; Hanashima, S.; Ikeda, A.; Morita-Matsumoto, K.; Satoh, T.; Matsumoto, N.; Yamamoto, K.; Yamaguchi, Y. Recognition of bisecting N-acetylglucosamine: Structural basis for asymmetric interaction with the mouse lectin dendritic cell inhibitory receptor 2. J. Biol. Chem. 2013, 288, 33598–33610. [Google Scholar] [CrossRef] [Green Version]
- Stanley, P.; Sundaram, S.; Tang, J.; Shi, S. Molecular analysis of three gain-of-function CHO mutants that add the bisecting GlcNAc to N-glycans. Glycobiology 2004, 15, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Raju, T.S.; Briggs, J.; Borge, S.M.; Jones, A.J.S. Species-specific variation in glycosylation of IgG: Evidence for the species-specific sialylation and branch-specific galactosylation and importance for engineering recombinant glycoprotein therapeutics. Glycobiology 2000, 10, 477–486. [Google Scholar] [CrossRef]
- Plaksin, D.; Jespersen, S.; Keinan, M.; Levin, I.; Cottingham, I. Glycosylation features of follitropin delta (rekovelle®) more resemble those of natural human fsh compared to follitropin alfa (Gonal f®). Fertil. Steril. 2021, 116, e417. [Google Scholar] [CrossRef]
- Karimi Alavijeh, M.; Meyer, A.S.; Gras, S.L.; Kentish, S.E. Synthesis of N-Acetyllactosamine and N-Acetyllactosamine-Based Bioactives. J. Agric. Food Chem. 2021, 69, 7501–7525. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D. Why are glycoproteins modified by poly-N-acetyllactosamine glycoconjugates? Curr. Protein Pept. Sci. 2003, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sytkowski, A.J.; Lunn, E.D.; Davis, K.L.; Feldman, L.; Siekman, S. Human erythropoietin dimers with markedly enhanced in vivo activity. Proc. Natl. Acad. Sci. USA 1998, 95, 1184–1188. [Google Scholar] [CrossRef] [Green Version]
- Nangia-Makker, P.; Hogan, V.; Raz, A. Galectin-3 and cancer stemness. Glycobiology 2018, 28, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Phang, R.; Lin, C.H. Synthesis of Type-I and Type-II LacNAc-Repeating Oligosaccharides as the Backbones of Tumor-Associated Lewis Antigens. Front. Immunol. 2022, 13, 858894. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; DiLillo, D.J.; Bournazos, S.; Giddens, J.P.; Ravetch, J.V.; Wang, L.-X. Modulating IgG effector function by Fc glycan engineering. Proc. Natl. Acad. Sci. USA 2017, 114, 3485–3490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalpathado, D.S.; Irungu, J.; Go, E.P.; Butnev, V.Y.; Norton, K.; Bousfield, G.R.; Desaire, H. Comparative Glycomics of the Glycoprotein Follicle Stimulating Hormone: Glycopeptide Analysis of Isolates from Two Mammalian Species. Biochemistry 2006, 45, 8665–8673. [Google Scholar] [CrossRef]
- Bishop, L.A.; Nguyen, T.V.; Schofield, P. Both of the beta-subunit carbohydrate residues of follicle-stimulating hormone determine the metabolic clearance rate and in vivo potency. Endocrinology 1995, 136, 2635–2640. [Google Scholar] [CrossRef]
- Dias, J.A.; Ulloa-Aguirre, A. New Human Follitropin Preparations: How Glycan Structural Differences May Affect Biochemical and Biological Function and Clinical Effect. Front. Endocrinol. 2021, 12, 636038. [Google Scholar] [CrossRef]
- North, S.J.; Huang, H.-H.; Sundaram, S.; Jang-Lee, J.; Etienne, A.T.; Trollope, A.; Chalabi, S.; Dell, A.; Stanley, P.; Haslam, S.M. Glycomics Profiling of Chinese Hamster Ovary Cell Glycosylation Mutants Reveals N-Glycans of a Novel Size and Complexity. J. Biol. Chem. 2010, 285, 5759–5775. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.A.H.; Bertozzi, C.R. The clinical impact of glycobiology: Targeting selectins, Siglecs and mammalian glycans. Nat. Rev. Drug Discov. 2021, 20, 217–243. [Google Scholar] [CrossRef]
- Jin, F.; Wang, F. The physiological and pathological roles and applications of sialyl Lewis x, a common carbohydrate ligand of the three selectins. Glycoconj. J. 2020, 37, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Uhler, R.; Popa-Wagner, R.; Kröning, M.; Brehm, A.; Rennert, P.; Seifried, A.; Peschke, M.; Krieger, M.; Kohla, G.; Kannicht, C.; et al. Glyco-engineered HEK 293-F cell lines for the production of therapeutic glycoproteins with human N-glycosylation and improved pharmacokinetics. Glycobiology 2021, 31, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.W.; Crispin, M.; Pritchard, L.; Robinson, H.; Gorny, M.; Yu, X.; Bailey-Kellogg, C.; Ackerman, M.E.; Scanlan, C.; Zolla-Pazner, S.; et al. Identification of antibody glycosylation structures that predict monoclonal antibody Fc-effector function. Aids 2014, 28, 2523–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houde, D.; Peng, Y.; Berkowitz, S.A.; Engen, J.R. Post-translational Modifications Differentially Affect IgG1 Conformation and Receptor Binding. Mol. Cell. Proteom. 2010, 9, 1716–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomann, M.; Schlothauer, T.; Dashivets, T.; Malik, S.; Avenal, C.; Bulau, P.; Rüger, P.; Reusch, D. In Vitro Glycoengineering of IgG1 and Its Effect on Fc Receptor Binding and ADCC Activity. PLoS ONE 2015, 10, e0134949. [Google Scholar] [CrossRef]
- Fokkink, W.J.; Selman, M.H.; Dortland, J.R.; Durmuş, B.; Kuitwaard, K.; Huizinga, R.; van Rijs, W.; Tio-Gillen, A.P.; van Doorn, P.A.; Deelder, A.M.; et al. IgG Fc N-glycosylation in Guillain-Barré syndrome treated with immunoglobulins. J. Proteome Res. 2014, 13, 1722–1730. [Google Scholar] [CrossRef] [PubMed]
- Karsten, C.M.; Pandey, M.K.; Figge, J.; Kilchenstein, R.; Taylor, P.R.; Rosas, M.; McDonald, J.U.; Orr, S.J.; Berger, M.; Petzold, D.; et al. Anti-inflammatory activity of IgG1 mediated by Fc galactosylation and association of FcgammaRIIB and dectin-1. Nat. Med. 2012, 18, 1401–1406. [Google Scholar] [CrossRef] [Green Version]
- Fortunato, M.E.; Colina, C.M. Effects of Galactosylation in Immunoglobulin G from All-Atom Molecular Dynamics Simulations. J. Phys. Chem. B 2014, 118, 9844–9851. [Google Scholar] [CrossRef]
- Krapp, S.; Mimura, Y.; Jefferis, R.; Huber, R.; Sondermann, P. Structural analysis of human IgG-Fc glycoforms reveals a correlation between glycosylation and structural integrity. J. Mol. Biol. 2003, 325, 979–989. [Google Scholar] [CrossRef]
- Aoyama, M.; Hashii, N.; Tsukimura, W.; Osumi, K.; Harazono, A.; Tada, M.; Kiyoshi, M.; Matsuda, A.; Ishii-Watabe, A. Effects of terminal galactose residues in mannose α1-6 arm of Fc-glycan on the effector functions of therapeutic monoclonal antibodies. MABS 2019, 11, 826–836. [Google Scholar] [CrossRef] [Green Version]
- Ju, M.-S.; Jung, S.T. Aglycosylated full-length IgG antibodies: Steps toward next-generation immunotherapeutics. Curr. Opin. Biotechnol. 2014, 30, 128–139. [Google Scholar] [CrossRef]
- Davis, J.S.; Kumar, T.R.; May, J.V.; Bousfield, G.R. Naturally Occurring Follicle-Stimulating Hormone Glycosylation Variants. J. Glycom. Lipidom. 2014, 4, e117. [Google Scholar]
- Carlsson, M.; Braddock, M.; Li, Y.; Wang, J.; Xu, W.; White, N.; Megally, A.; Hunter, G.; Colice, G. Evaluation of Antibody Properties and Clinically Relevant Immunogenicity, Anaphylaxis, and Hypersensitivity Reactions in Two Phase III Trials of Tralokinumab in Severe, Uncontrolled Asthma. Drug Saf. 2019, 42, 769–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, R.S.; Abernathy, M.; Richardson, D.D.; Rouse, J.C.; Sperry, J.B.; Swann, P.; Wypych, J.; Yu, C.; Zang, L.; Deshpande, R. A View on the Importance of “Multi-Attribute Method” for Measuring Purity of Biopharmaceuticals and Improving Overall Control Strategy. AAPS J. 2017, 20, 7. [Google Scholar] [CrossRef] [Green Version]
- Hennrich, M.L.; Gavin, A.-C. Quantitative mass spectrometry of posttranslational modifications: Keys to confidence. Sci. Signal. 2015, 8, re5. [Google Scholar] [CrossRef] [PubMed]
- Raymond, C.; Robotham, A.; Spearman, M.; Butler, M.; Kelly, J.; Durocher, Y. Production of IgGs with a human-like sialylation in CHO cells. BMC Proc. 2015, 9, O3. [Google Scholar] [CrossRef] [Green Version]
- Rathore, A.; Malani, H. Need for a risk-based control strategy for managing glycosylation profile for biosimilar products. Expert Opin. Biol. Ther. 2022, 22, 123–131. [Google Scholar] [CrossRef]
- Parenky, A.; Myler, H.; Amaravadi, L.; Bechtold-Peters, K.; Rosenberg, A.; Kirshner, S.; Quarmby, V. New FDA Draft Guidance on Immunogenicity. AAPS J. 2014, 16, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Brühlmann, D.; Vuillemin, T.; Satwekar, A.; Galano, E.; Palmese, A.; D’Angelo, A.; Manco, Z.; Souquet, J.; Broly, H.; Sauer, M.; et al. Generation of site-distinct N-glycan variants for in vitro bioactivity testing. Biotechnol. Bioeng. 2019, 116, 1017–1028. [Google Scholar] [CrossRef]
- Mastrangeli, R.; Satwekar, A.; Cutillo, F.; Ciampolillo, C.; Palinsky, W.; Longobardi, S. In-vivo biological activity and glycosylation analysis of a biosimilar recombinant human follicle-stimulating hormone product (Bemfola) compared with its reference medicinal product (GONAL-f). PLoS ONE 2017, 12, e0184139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Go, E.P.; Desaire, H. Absolute Quantitation of Glycosylation Site Occupancy Using Isotopically Labeled Standards and LC-MS. J. Am. Soc. Mass Spectrom. 2014, 25, 1012–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Li, W.; Lu, H.; Liu, Y. Quantification of N-glycosylation site occupancy status based on labeling/label-free strategies with LC-MS/MS. Talanta 2017, 170, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Delafield, D.G.; Li, L. Recent Advances in Analytical Approaches for Glycan and Glycopeptide Quantitation. Mol. Cell. Proteom. 2021, 20, 100054. [Google Scholar] [CrossRef]




| Cell Line | Product Class | Glycan Related Indices Per Site | Whole Molecule Glycosylation Indices | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOI | AI | SI | SIα2,6 | SE | SINeu5Gc | % Neu5Gc | % O-acetyl | αGI | GI | G2 | G11-6 | G1 | G0 | MI | HI | cFI | aFI | LI | BI | AI | SI | SE | SIα2,6 | SI-Neu5Gc | αGI | MI | ||
| Murine | mAbs (only Fc glycosylation) | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓Fc | ✓ Fc | ✓Fc | ✓Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||
| mAbs with Fc and Fab glycosylation | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |
| Fc and antibody fusion proteins | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |
| Any other class of protein | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||
| CHO | Mab (only Fc glycosylation) | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||
| mAbs with Fc and Fab glycosylation | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||
| Fc and antibody fusion proteins | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||
| Any other class of protein | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||
| Human | Mab (only Fc glycosylation) | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||||
| mAbs with Fc and Fab glycosylation | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| Fc and antibody fusion proteins | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ Fc | ✓ Fc | ✓ Fc | ✓ Fc | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| Any other class of protein | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| Cell Line | Specific PTM | Potential Impact |
|---|---|---|
| Murine (NS0 and Sp2/0) | αGal | Immunogenicity, clearance, and safety |
| Neu5Gc | Immunogenicity, clearance, and safety | |
| Sialic acid linkage α2,3 and α2,6 | Impact on clearance dependent on SE and SI. In rodents, glycoproteins carrying α2,6-linked sialic acid, but not those carrying α2,3-linked sialic acid, have been reported to interact with the asialoglycoprotein receptor resulting in a more rapid clearance [27]. A comparable effect in humans is still to be demonstrated. However, a differential impact on clearance cannot be ruled out. | |
| CHO | Neu5Gc (low level) | Immunogenicity, clearance, and safety |
| αGal (possible very low level) | Immunogenicity, clearance, and safety | |
| Sialic acid linkage: only α2,3 | Impact on clearance dependent on SE and SI. | |
| Human | Sialic acid linkage α2,3 and α2,6 | Impact on clearance dependent on SE and SI. In rodents, glycoproteins carrying α2,6-linked sialic acid, but not those carrying α2,3-linked sialic acid, have been reported to interact with the asialoglycoprotein receptor resulting in a more rapid clearance [27]. A comparable effect in humans is still to be demonstrated. However, a differential impact on clearance cannot be ruled out. |
| Bisecting GlcNAc | Impedes core fucosylation. Therefore, has an impact on mAb efficacy in Fc glycans. Potential interaction with C-type lectin receptors in exposed glycans. |
| Glycan Related CQA Indices | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glycosite | SOI | AI | SI | SIα2,6 | SE | SI Neu5Gc | % Neu5Gc | % O-acetyl | αGI | GI | G2 | G11-6 | G1 | G0 | MI | HI | cFI | aFI | LI | BI |
| Fc | ✓ | ✓ | ✓ | NA | ✓ | ✓ | ✓ | NA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | NA | NA | NA |
| N1 | ✓ | ✓ | ✓ | NA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | NA | NA | NA | NA | ✓ | ✓ | ✓ | ✓ | ✓ | NA |
| N2 | ✓ | ✓ | ✓ | NA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | NA | NA | NA | NA | ✓ | ✓ | ✓ | ✓ | ✓ | NA |
| Whole molecules/domains | NA | ✓ | ✓ | NA | ✓ * | ✓ | NA | NA | ✓ | NA | NA | NA | NA | NA | ✓ | NA | NA | ✓ | ✓ | NA |
| Glycosite | RHS Characterization Matrix | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI | ||
| Fc | Mean | 1.0 | 1.77 | ND | <0.01 | 0.00 | ND | ND | 0.20 | 0.02 | 0.98 |
| CV% | - (1) | 1.54 | NA | NA | NA | NA | NA | 10.69 | 23.99 | 0.52 | |
| StDev | - (1) | 0.03 | NA | NA | 0.00 | NA | NA | 0.02 | 0.00 | 0.01 | |
| N1 | Mean | 1.0 | 2.64 | 1.73 | 0.65 | 0.01 | 0.34 | 2.29 | 0.64 | 0.01 | 0.99 |
| CV% | - (1) | 2.50 | 3.39 | 2.85 | 30.68 | 29.50 | 56.98 | 7.20 | 15.74 | 0.32 | |
| StDev | - (1) | 0.07 | 0.06 | 0.02 | 0.00 | 0.10 | 1.30 | 0.05 | 0.00 | 0.00 | |
| N2 | Mean | 0.6 | 3.42 | 2.43 | 0.71 | 0.00 | 0.20 | 1.96 | 0.80 | ND | 1.00 |
| CV% | - (1) | 0.78 | 1.33 | 1.45 | 0.00 | 26.41 | 59.55 | 3.93 | NA | 0.46 | |
| StDev | - (1) | 0.03 | 0.03 | 0.01 | 25.64 | 0.05 | 1.17 | 0.03 | NA | 0.00 | |
| Overall whole molecule/domains | 9.33 | 6.34 | 0.68 | 0.02 | 0.06 | ||||||
| A | 60% Fused Protein Domain with N1 and N2 Glycosylated Sites | |||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI |
| Fc | 1.0 | 1.77 | ND | <0.01 | ND | ND | ND | 0.20 | 0.02 | 0.98 |
| N1 | 1.0 | 2.64 | 1.73 | 0.65 | 0.01 | 0.34 | 2.29 | 0.64 | 0.01 | 0.99 |
| N2 | 1.0 | 3.42 | 2.43 | 0.71 | 0.00 | 0.20 | 1.96 | 0.80 | 0.00 | 1.00 |
| Fused domain A | 6.03 | 4.14 | 0.69 | 0.01 | 0.03 | |||||
| B | 40% Fused Domain with N1 Glycosylated Sites Only | |||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI |
| Fc | 1.0 | 1.77 | ND | <0.01 | ND | ND | ND | 0.20 | 0.02 | 0.98 |
| N1 | 1.0 | 2.64 | 1.73 | 0.66 | 0.01 | 0.34 | 2.29 | 0.64 | 0.01 | 0.99 |
| N2 | 0.0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fused domain B | 2.61 | 1.71 | 0.66 | 0.01 | 0.03 | |||||
| N-Glycan Sites | RHS | P1 | P2 | P3 | P4 | P5 | P6 | |
|---|---|---|---|---|---|---|---|---|
| AI | Fc | 1.81 | 1.82 | 1.76 | 1.72 | 1.82 | 1.88 | 1.77 |
| N1 | 2.59 | 2.53 | 2.54 | 2.50 | 2.53 | 2.61 | 2.57 | |
| N2 | 3.46 | 3.41 | 3.58 | 3.50 | 3.39 | 3.27 | 3.54 | |
| Whole | 9.28 | 9.10 | 9.33 | 9.15 | 9.08 | 9.09 | 9.39 | |
| SI | Fc | ND | ND | ND | ND | ND | ND | ND |
| N1 | 1.68 | 1.43 | 2.08 | 2.04 | 1.45 | 2.14 | 1.47 | |
| N2 | 2.47 | 2.38 | 3.02 | 3.04 | 2.38 | 2.21 | 3.03 | |
| Whole | 6.29 | 5.69 | 7.74 | 7.69 | 5.73 | 6.89 | 6.58 | |
| SE | Fc | ND | ND | ND | ND | ND | ND | ND |
| N1 | 0.64 | 0.56 | 0.82 | 0.82 | 0.57 | 0.83 | 0.56 | |
| N2 | 0.71 | 0.68 | 0.85 | 0.88 | 0.68 | 0.66 | 0.86 | |
| Whole | 0.68 | 0.62 | 0.83 | 0.84 | 0.63 | 0.76 | 0.70 | |
| SINeu5Gc | Fc | ND | ND | ND | ND | ND | ND | ND |
| N1 | 0.01 | 0.01 | 0.03 | 0.03 | 0.01 | 0.02 | 0.02 | |
| N2 | 0.00 | 0.01 | 0.03 | 0.02 | 0.01 | 0.02 | 0.04 | |
| Whole | 0.02 | 0.04 | 0.10 | 0.10 | 0.04 | 0.07 | 0.08 | |
| %Neu5Gc | Fc | ND | ND | ND | ND | ND | ND | ND |
| N1 | 0.30 | 0.71 | 1.62 | 1.70 | 0.98 | 0.94 | 1.10 | |
| N2 | 0.20 | 0.55 | 0.99 | 0.79 | 0.59 | 1.13 | 1.19 | |
| %O-acetyl | Fc | ND | ND | ND | ND | ND | ND | ND |
| N1 | 1.67 | 0.21 | 0.49 | 0.65 | 0.21 | 0.33 | 0.21 | |
| N2 | 2.55 | 0.17 | 0.33 | 0.36 | 0.38 | 0.18 | 0.23 | |
| GI | Fc | 0.17 | 0.19 | 0.27 | 0.30 | 0.20 | 0.13 | 0.22 |
| N1 | 0.64 | 0.30 | 0.41 | 0.39 | 0.28 | 0.37 | 0.23 | |
| N2 | 0.82 | 0.35 | 0.51 | 0.43 | 0.32 | 0.26 | 0.47 | |
| MI | Fc | 0.01 | 0.01 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 |
| N1 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.00 | |
| N2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Whole * | 0.03 | 0.03 | 0.03 | 0.03 | 0.02 | 0.02 | 0.01 | |
| cFI | Fc | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 |
| N1 | 0.99 | 0.99 | 0.99 | 0.99 | 1.00 | 0.99 | 1.00 | |
| N2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Pre-Change Condition (Batches n = 4) | ||||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI |
| Fc | 1.0 (1) | 1.74 | ND | <0.01 | ND | ND | ND | 0.24 | 0.02 | 0.97 |
| (CV%) | −0.7 | NA | NA | NA | NA | NA | −5.9 | −14.1 | 0 | |
| N1 | 1.0 (1) | 2.55 | 1.85 | 0.72 | 0.01 | 0.53 | 4.09 | 0.48 | 0.01 | 0.99 |
| (CV%) | −1.1 | −1.8 | −1 | −12.9 | −11.6 | −16.8 | −8.2 | −22.3 | 0 | |
| N2 | 0.6 (1) | 3.42 | 2.66 | 0.77 | 0.01 | 0.22 | 3.97 | 0.57 | ND | 1 |
| (CV%) | −0.8 | −2.4 | −2.1 | −16.7 | −15.4 | −13.8 | −8.7 | NA | 0 | |
| 2 fused domains | 9.15 | 6.86 | 0.75 | 0.03 | 0.05 * | |||||
| Post-Change Condition (Batches n = 4) | ||||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI |
| Fc | 1.0 (1) | 1.73 | ND | <0.01 | ND | ND | ND | 0.24 | 0.02 | 0.97 |
| (CV%) | −0.6 | NA | 0 | NA | NA | NA | −2.5 | −2.5 | 0 | |
| N1 | 1.0 (1) | 2.56 | 1.91 | 0.75 | 0.01 | 0.46 | 4.67 | 0.43 | 0.01 | 1 |
| (CV%) | −1 | −1.8 | −3 | −10.9 | −12.2 | −9 | −9.9 | −22.3 | −0.5 | |
| N2 | 0.6 (1) | 3.44 | 2.66 | 0.77 | 0.01 | 0.22 | 3.35 | 0.58 | ND | 1 |
| (CV%) | −0.8 | −1.5 | −1.8 | −16.7 | −17.2 | −7.9 | −5.7 | NA | 0 | |
| 2 fused domains | 9.2 | 6.97 | 0.76 | 0.03 | 0.05 * | |||||
| Product A (Batches n = 3) | |||||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI | LI |
| α-N1 | 1 *1 | 2.12 | 1.75 | 0.82 | 0 | 0.21 | 2.46 | 0.37 | ND | 0.02 | 0 |
| α-N2 | 1 *1 | 2.08 | 1.77 | 0.85 | ND | ND | 4.93 | 0.32 | ND | 0.02 | 0 |
| β-N1 | 1 *1 | 3.54 | 2.25 | 0.69 | ND | ND | 2.01 | 1.43 | ND | 0.57 | 0.32 |
| β-N2 | 1 *1 | 2.16 | 1.87 | 0.88 | 0 | 0.18 | 6.98 | 0.21 | ND | 1.02 | 0 |
| Whole | 9.9 | 7.64 | 0.77 | 0.01 | 0.32 | ||||||
| Product B (Batches n = 2) | |||||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI | LI |
| α-N1 | 1 *1 | 2.27 | 1.79 | 0.79 | 0 | 0.17 | 0 | 0.49 | ND | 0.01 | 0 |
| α-N2 | 1 *1 | 2.24 | 1.78 | 0.81 | 0.01 | 0.27 | 0.35 | 0.47 | ND | 0.01 | 0 |
| β-N1 | 1 *1 | 4.14 | 2.7 | 0.69 | ND | ND | 0.26 | 1.22 | ND | 0.55 | 0.38 |
| β-N2 | 1 *1 | 2.25 | 1.95 | 0.88 | 0.01 | 0.26 | 1.05 | 0.21 | ND | 1.01 | 0.01 |
| Whole | 10.89 | 8.2 | 0.75 | 0.01 | 0.39 | ||||||
| Product C (Batches n = 3) | |||||||||||
| Glycosite | SOI | AI | SI | SE | SINeu5Gc | %Neu5Gc | %O-acetyl | GI | MI | cFI | LI |
| α-N1 | 1 *1 | 2.1 | 1.81 | 0.86 | 0.08 | 4.18 | 1.78 | 0.28 | ND | 0.01 | 0 |
| α-N2 | 1 *1 | 2.08 | 1.86 | 0.9 | 0.08 | 4.31 | 5.79 | 0.21 | ND | 0.01 | 0 |
| β-N1 | 1 *1 | 3.77 | 2.19 | 0.64 | ND | ND | 2.03 | 1.73 | ND | 0.49 | 0.41 |
| β-N2 | 1 *1 | 2.22 | 1.99 | 0.92 | 0.07 | 3.6 | 6.25 | 0.11 | ND | 0.99 | 0 |
| Whole | 10.16 | 7.85 | 0.77 | 0.23 | 0.41 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrangeli, R.; Satwekar, A.; Bierau, H. Innovative Metrics for Reporting and Comparing the Glycan Structural Profile in Biotherapeutics. Molecules 2023, 28, 3304. https://doi.org/10.3390/molecules28083304
Mastrangeli R, Satwekar A, Bierau H. Innovative Metrics for Reporting and Comparing the Glycan Structural Profile in Biotherapeutics. Molecules. 2023; 28(8):3304. https://doi.org/10.3390/molecules28083304
Chicago/Turabian StyleMastrangeli, Renato, Abhijeet Satwekar, and Horst Bierau. 2023. "Innovative Metrics for Reporting and Comparing the Glycan Structural Profile in Biotherapeutics" Molecules 28, no. 8: 3304. https://doi.org/10.3390/molecules28083304