

Discovery, Antitumor Activity, and Fermentation Optimization of Roquefortines from Penicillium sp. OUCMDZ-1435
Abstract
:1. Introduction
2. Results
2.1. The Target Fungal Strain
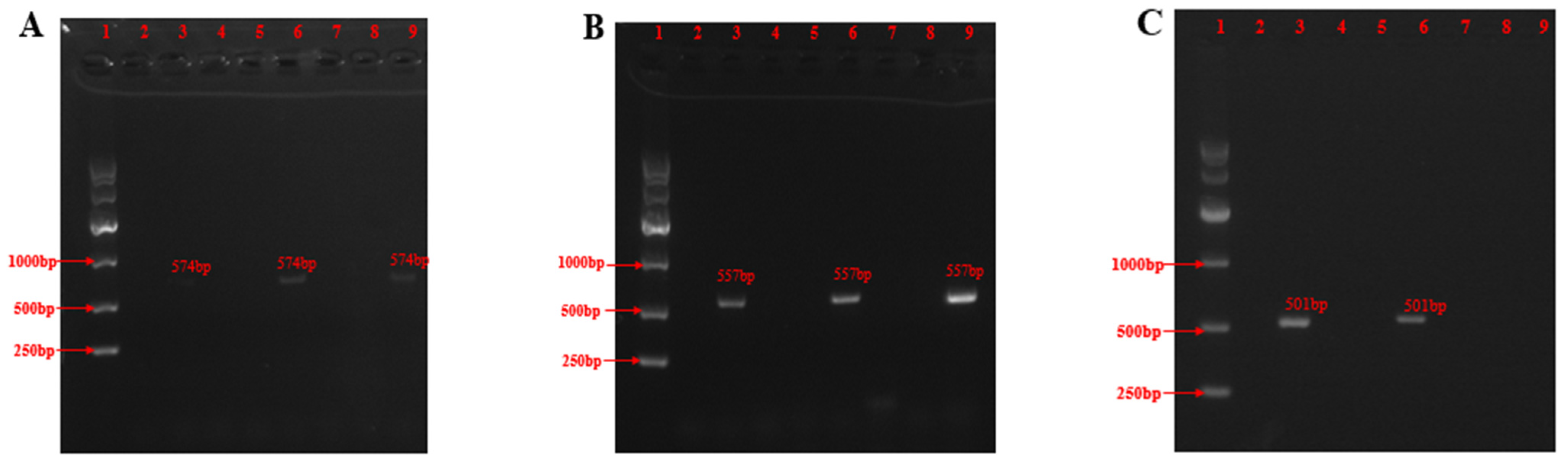
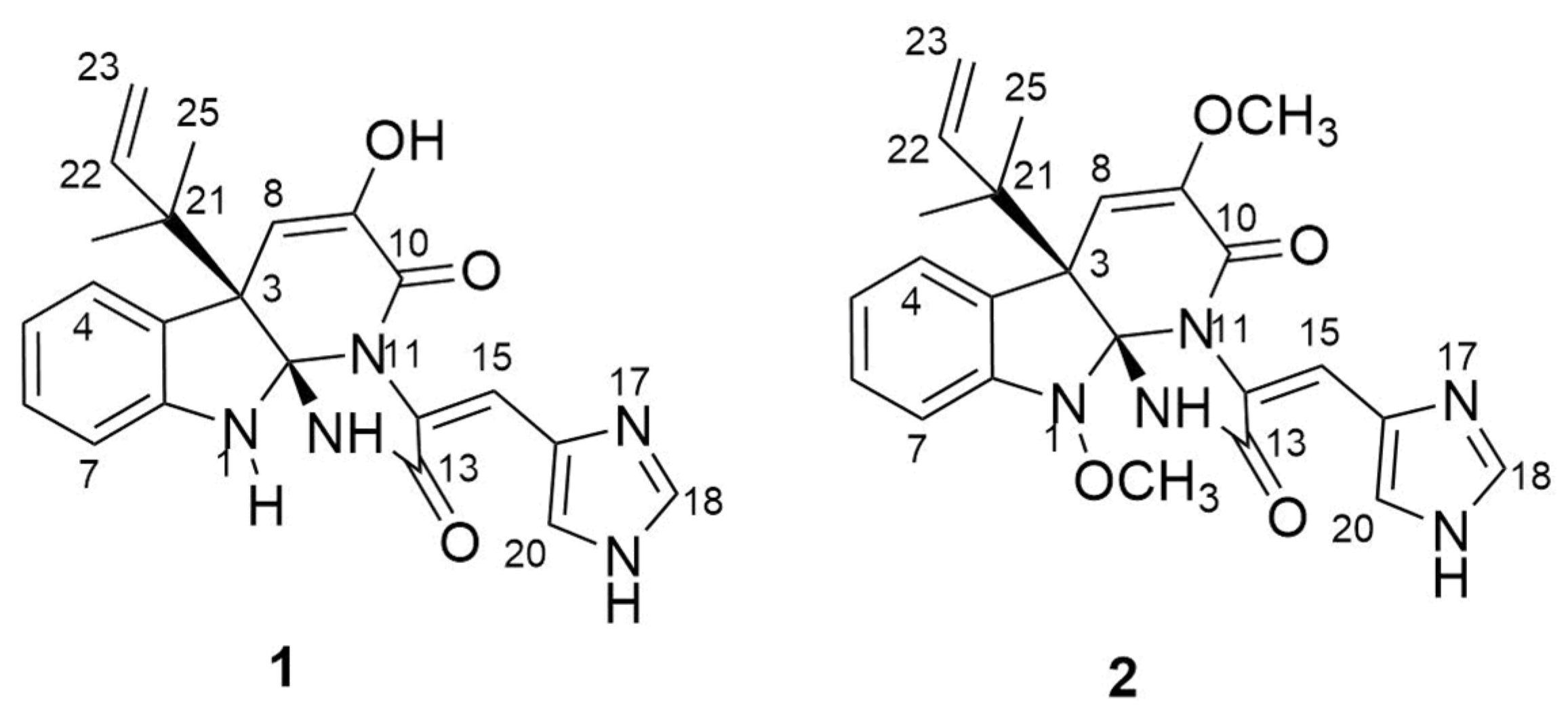
2.2. Isolation and Identification
2.3. Biological Activity
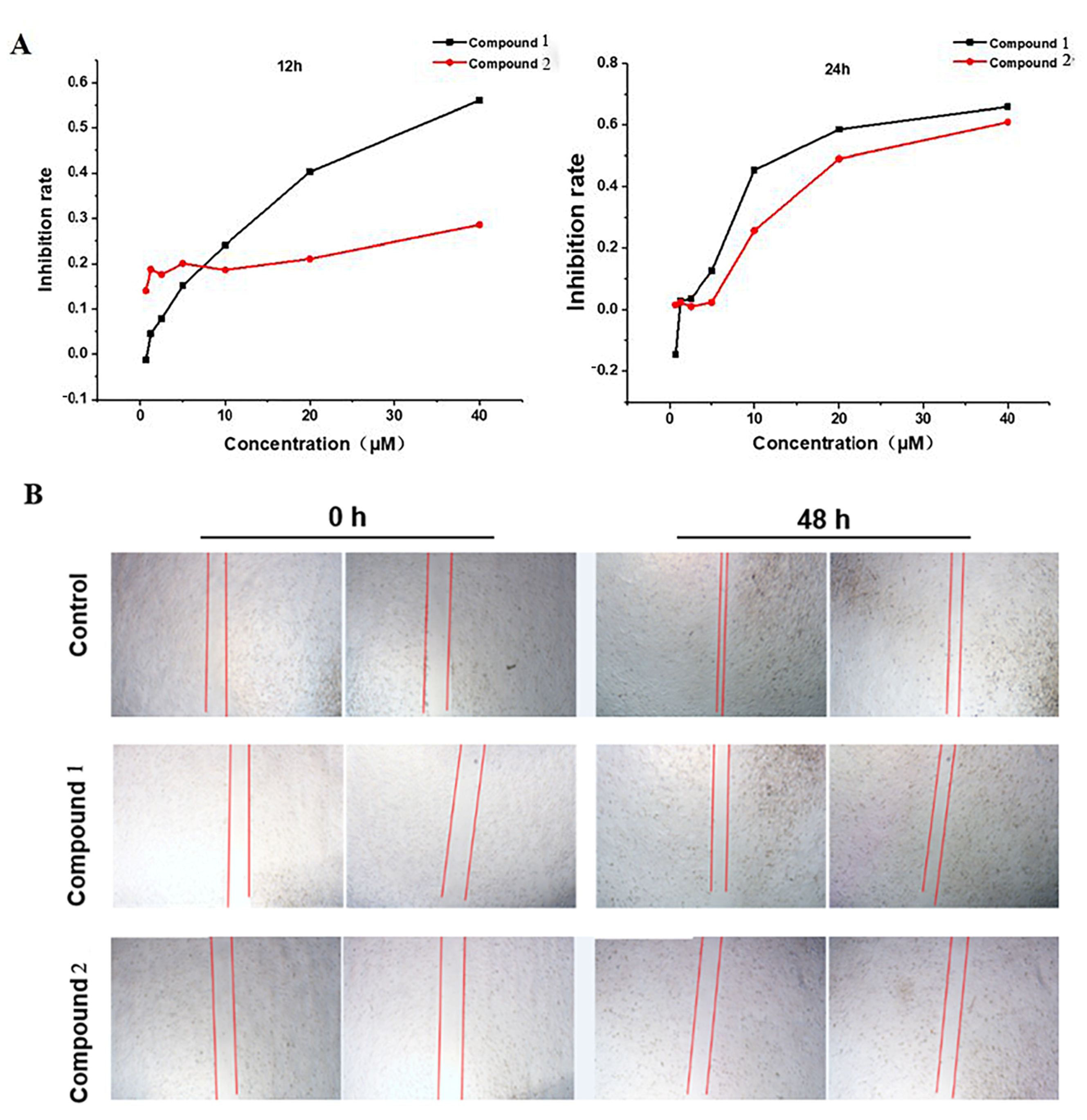
2.3.1. Cytotoxicity
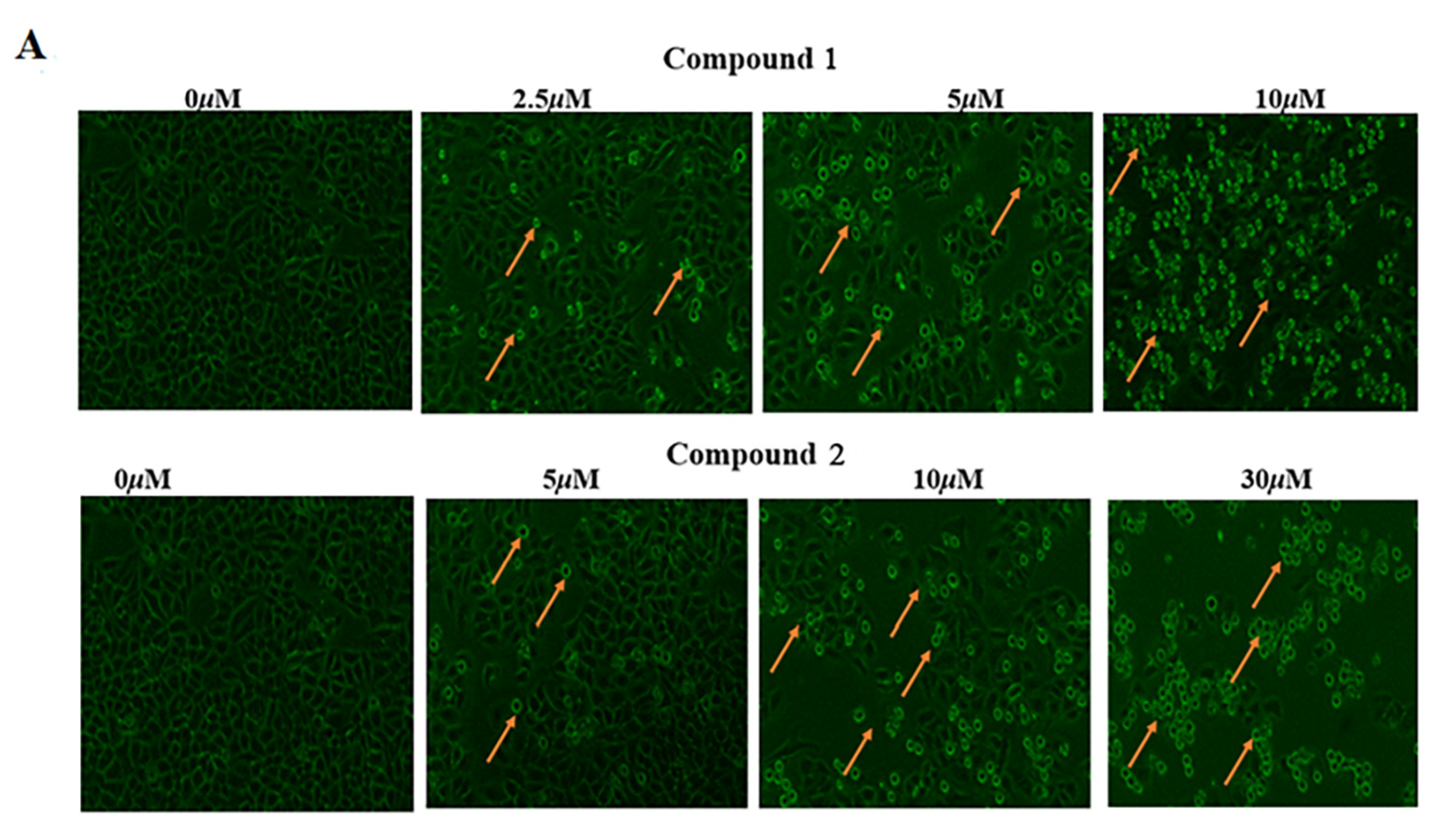
2.3.2. Cell Proliferation and Migration Assay
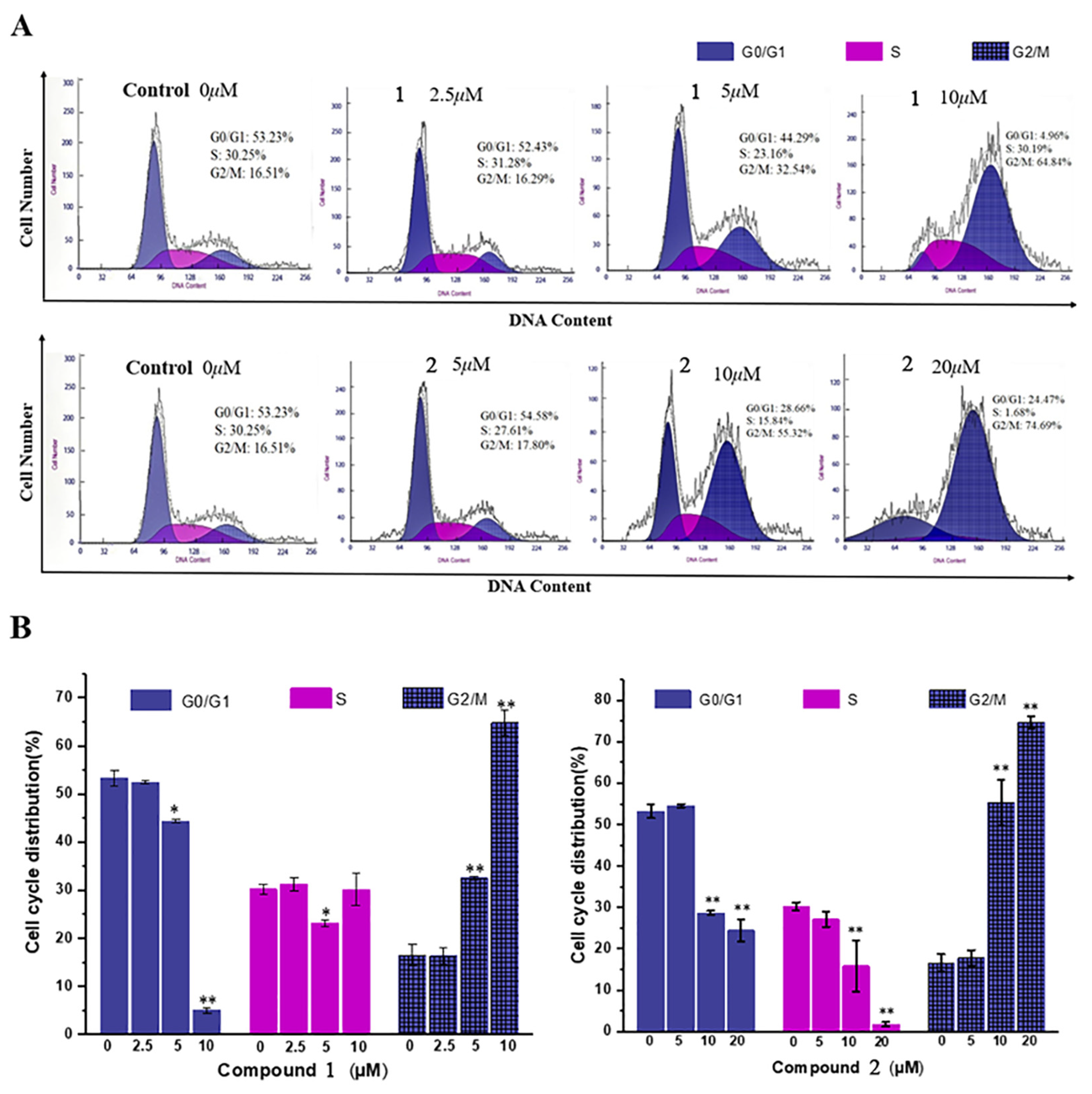
2.3.3. Cell Cycle Arrest Assay
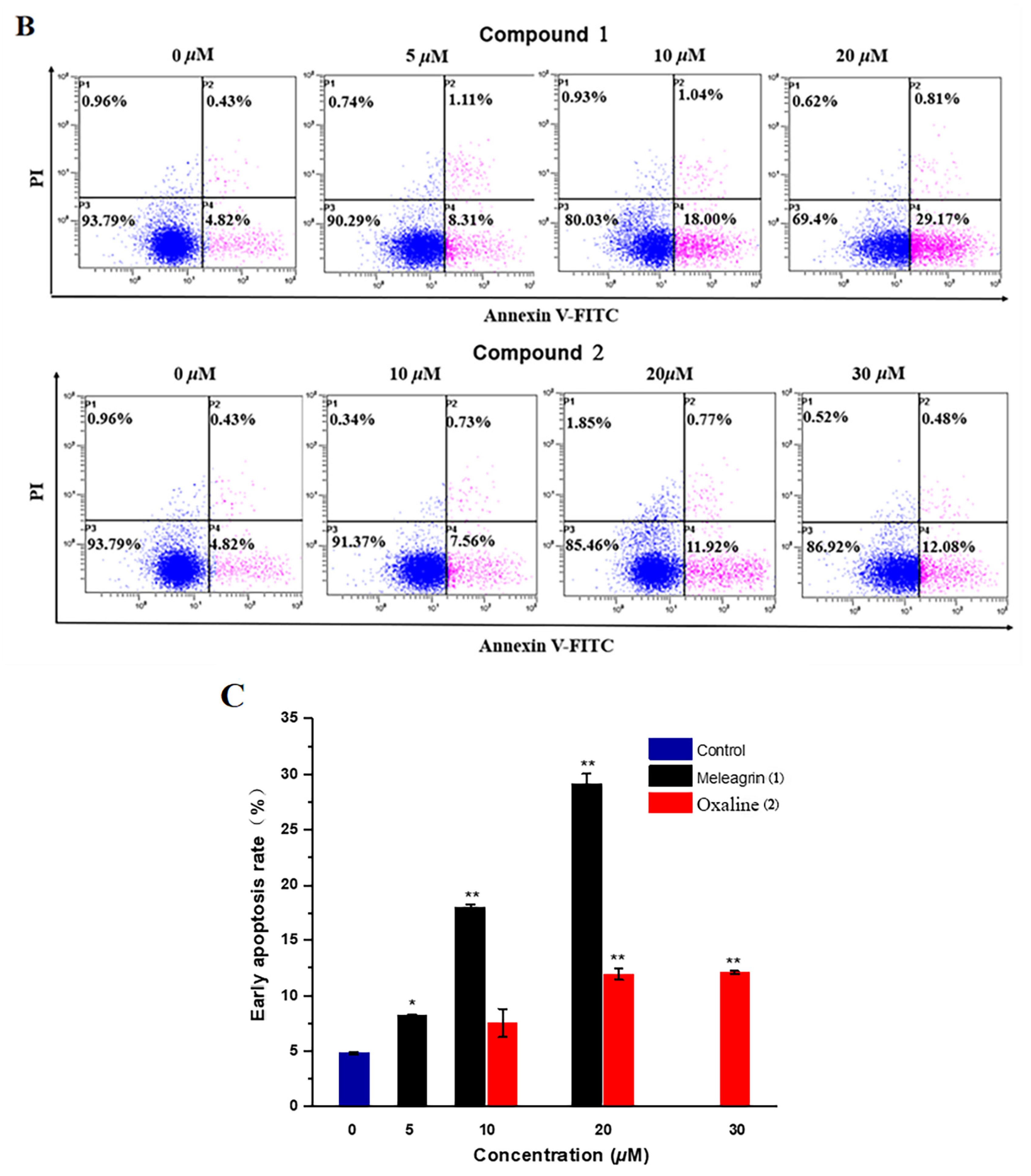
2.3.4. Cell Apoptosis Assay
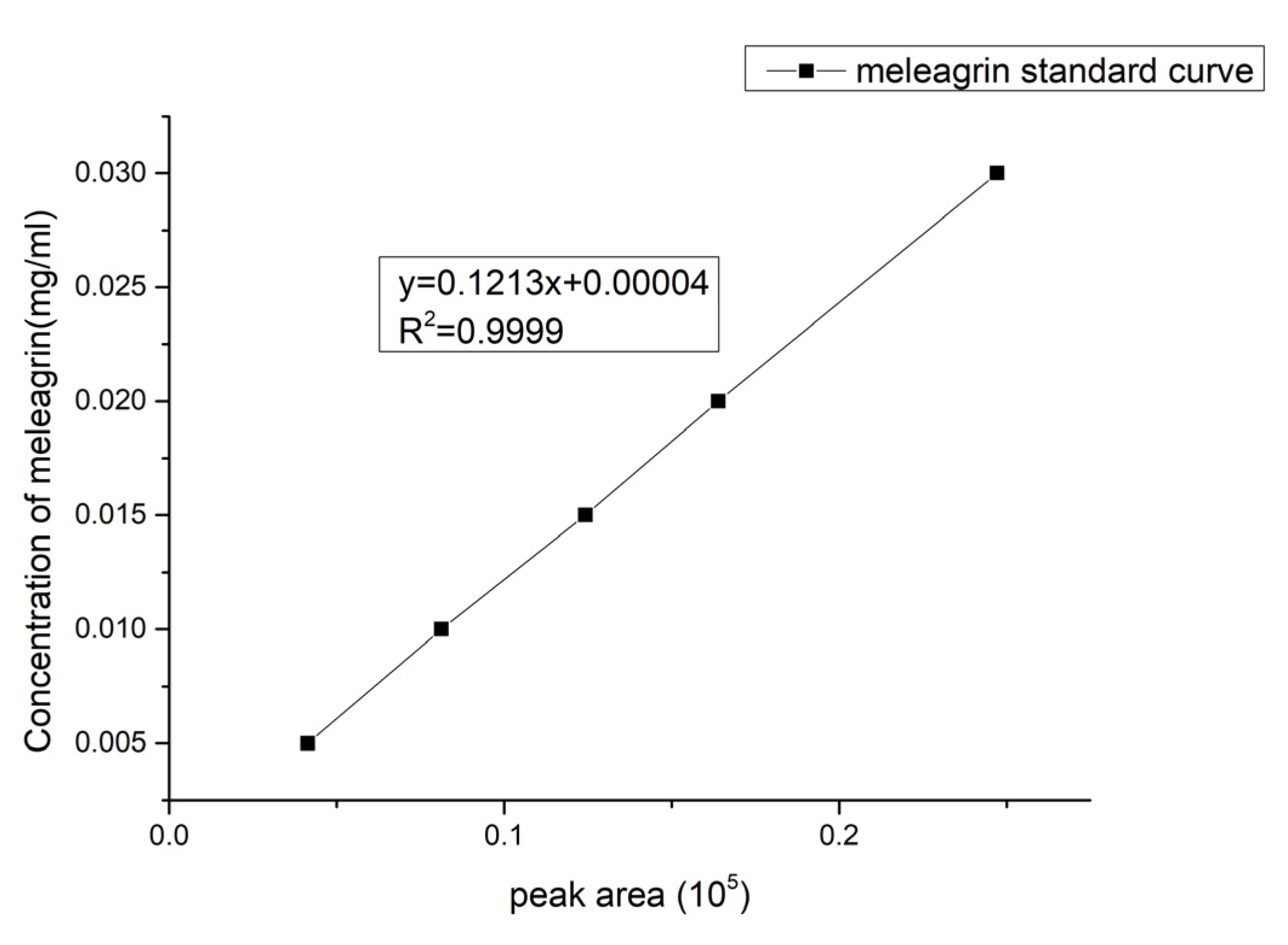
2.4. Meleagrin Standard Curve
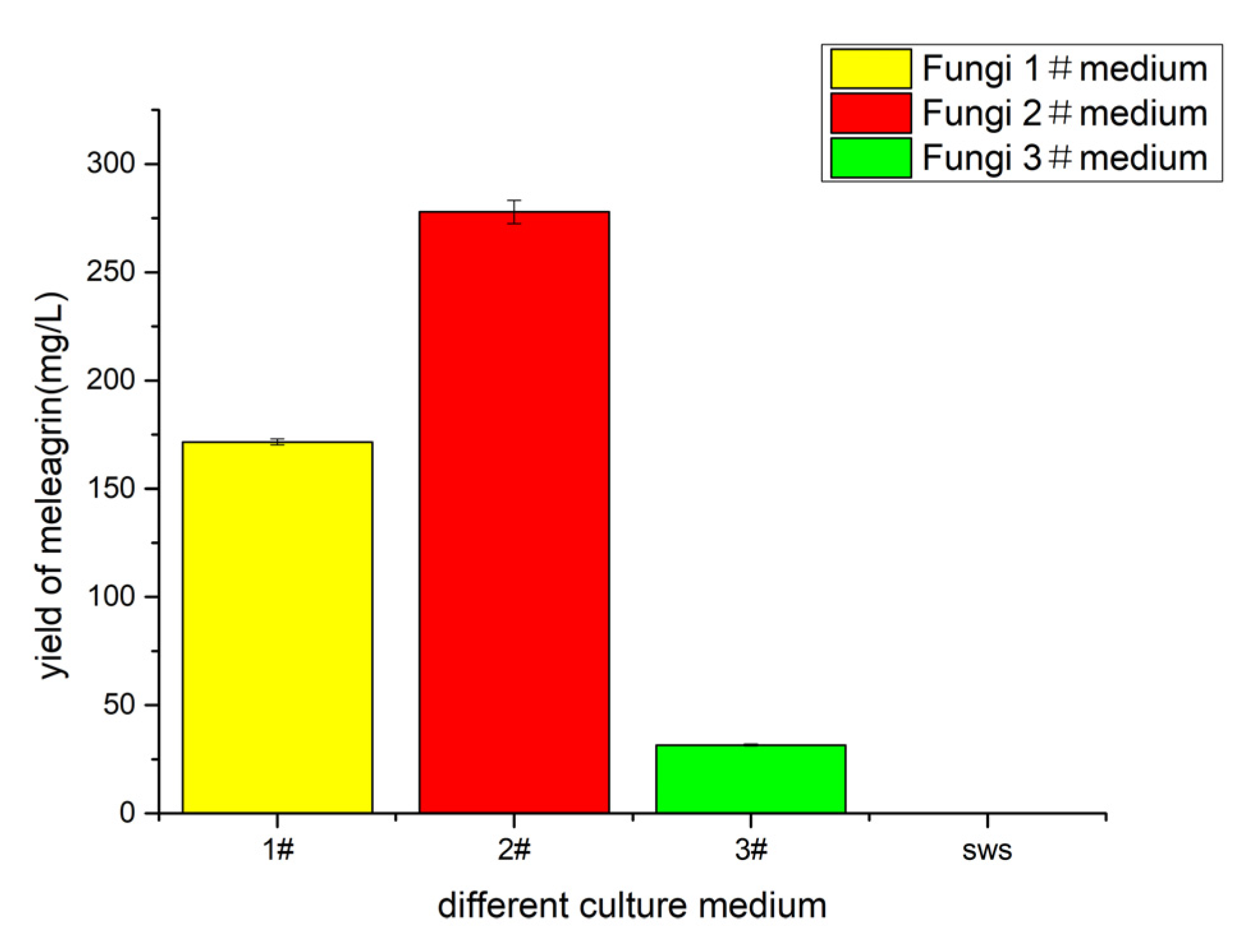
2.5. Determination of Basic Medium
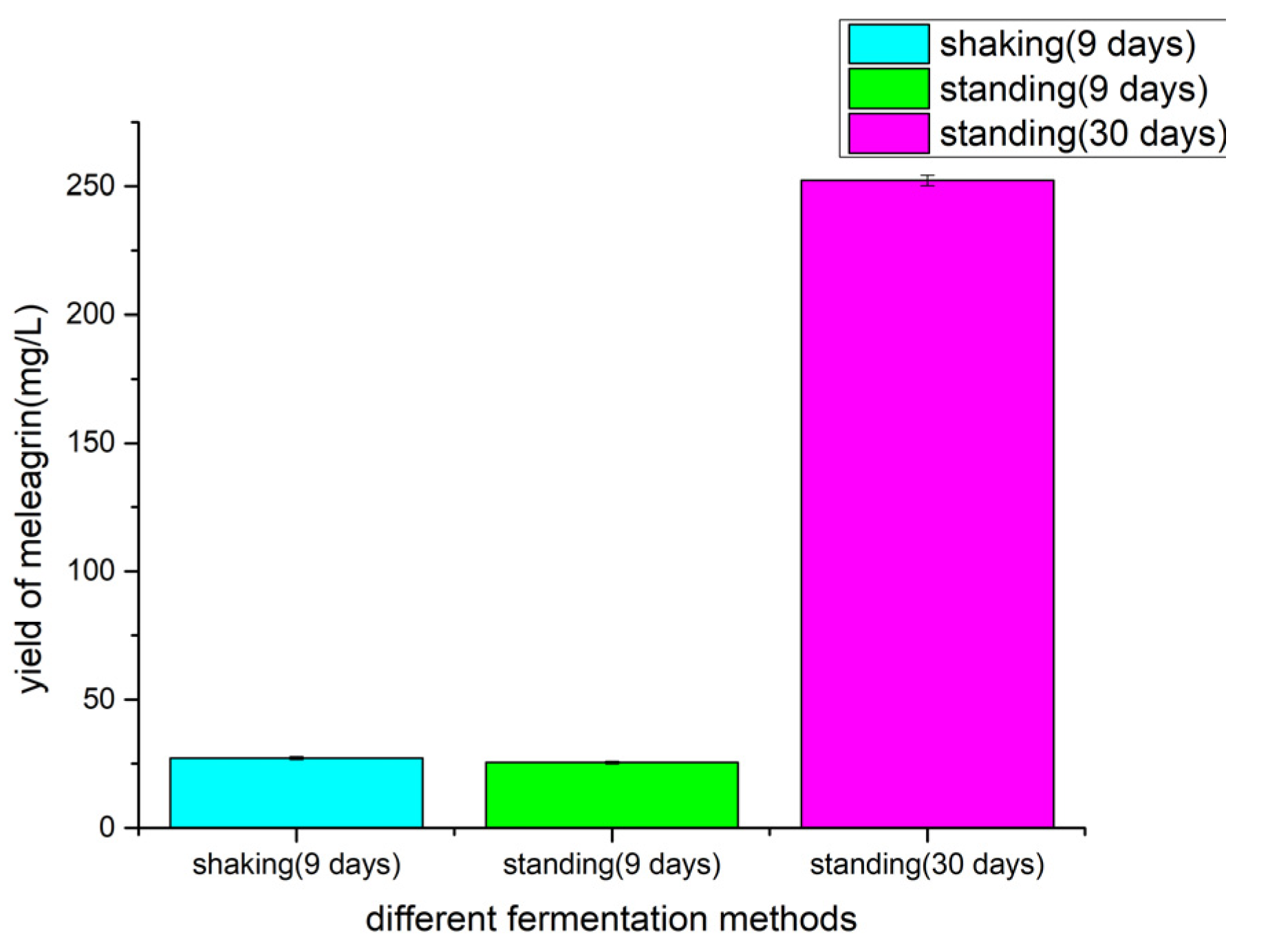
2.6. Determination of Fermentation Methods
2.7. Single-Factor Optimization
2.7.1. Effect of Fermentation Days on Meleagrin Yield
2.7.2. Effect of Initial Fermentation pH on Meleagrin Production
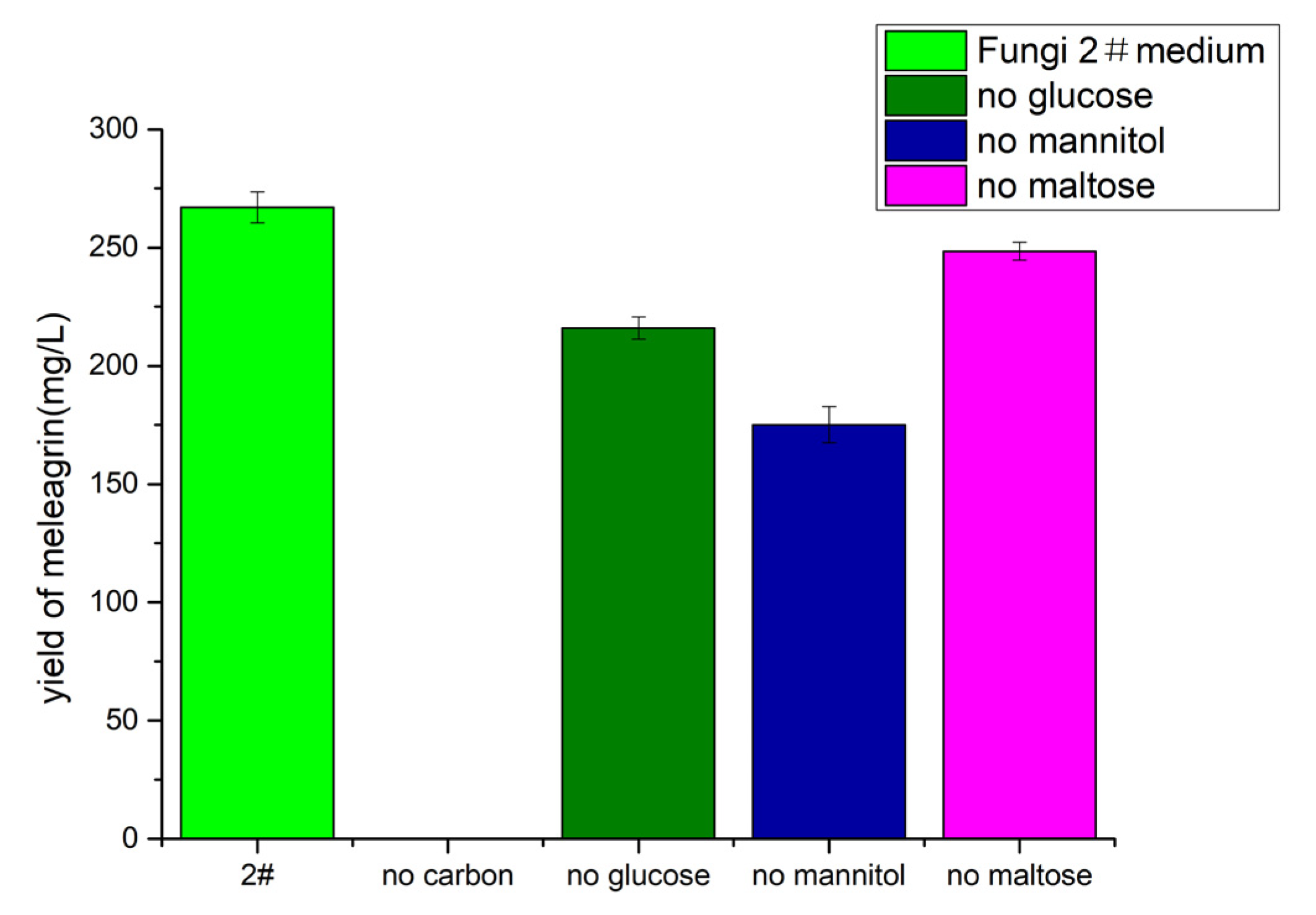
2.7.3. Effects of Different Carbon Sources on Meleagrin Production
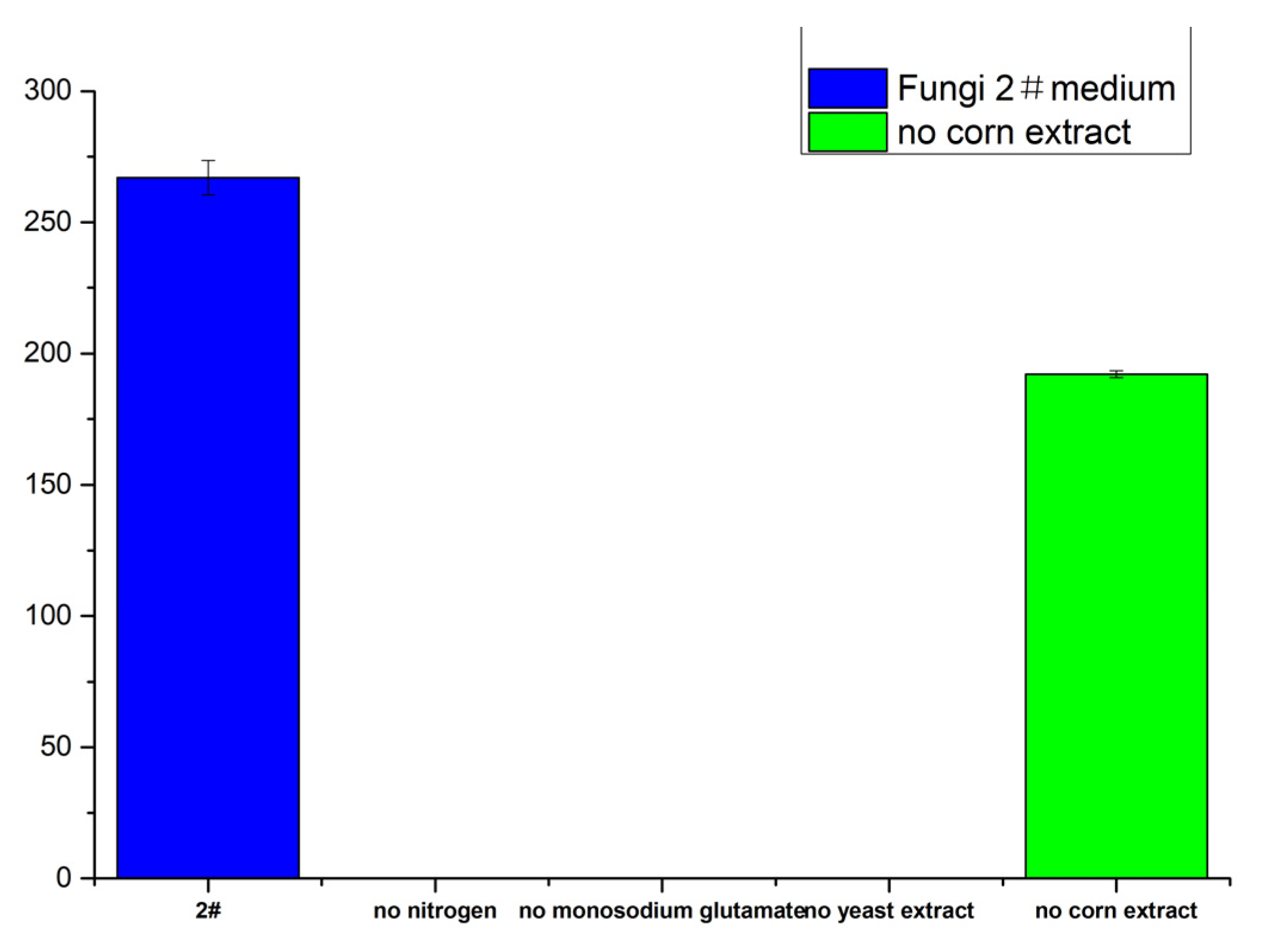
2.7.4. Effects of Different Nitrogen Sources on Meleagrin Production
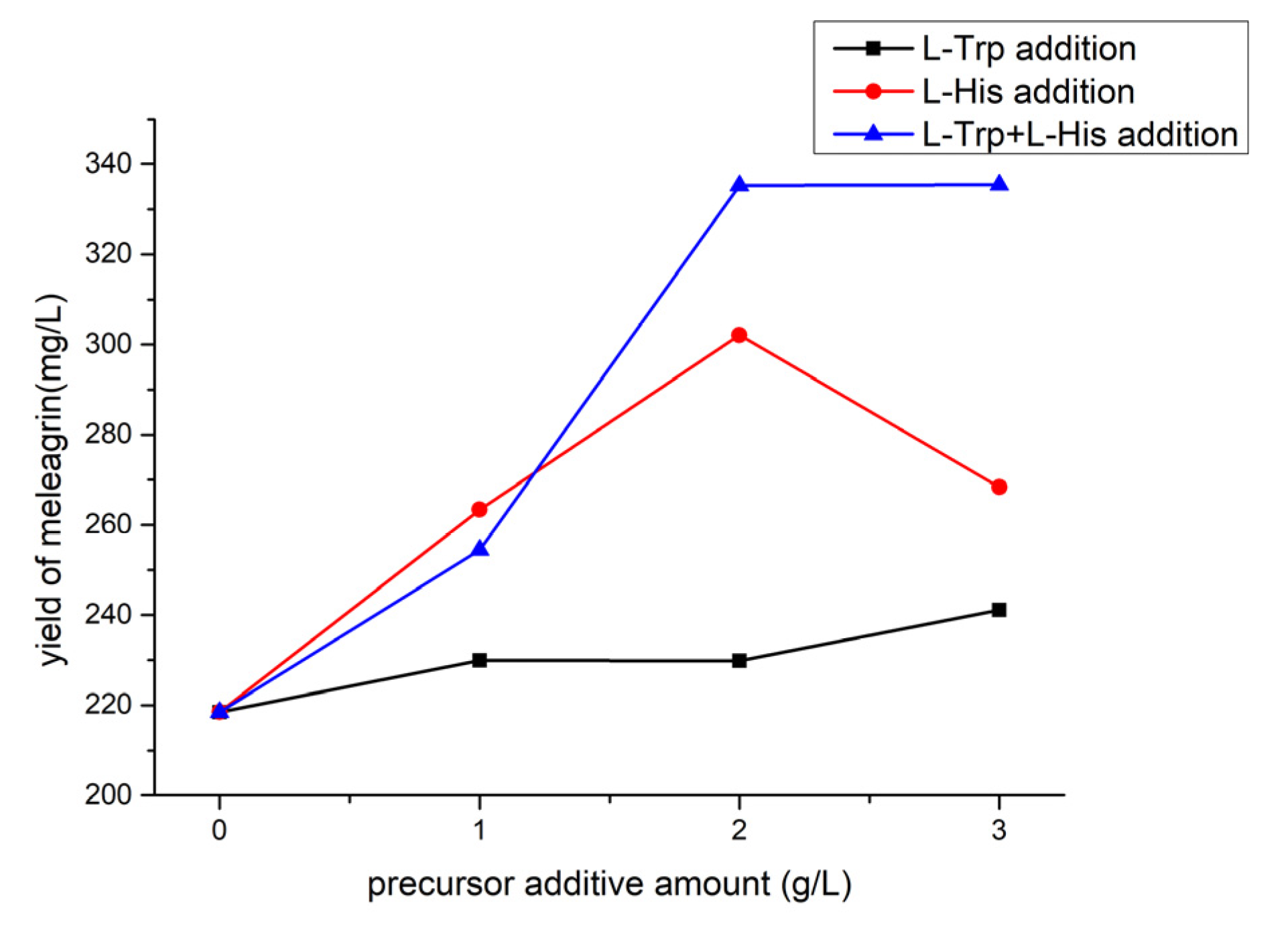
2.7.5. Effect of Precursor Addition on Meleagrin Production
3. Discussion
4. Materials and Methods
4.1. Strain
4.2. Biological Probes
4.3. Fermentation and Extraction
4.4. Isolation and Identification
4.5. Cell Culture and Cytotoxicity Assays
4.6. Proliferation and Metastasis Assay
4.7. Cell Cycle Analysis
4.8. Cell Death Assay
4.9. Quantitative Detection of Meleagrin and Establishment of the Standard Curve
4.10. Single-Factor Optimization of Fermentation Conditions
4.10.1. Determination of the Influence of Different Fermentation Days
4.10.2. Determination of the Influence of Different Initial pH
4.10.3. Determination of the Influence of Different Carbon Sources
4.10.4. Determination of the Influence of Different Nitrogen Sources
4.10.5. Determination of the Influence of Different Precursor Addition Experiments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kozlovskii, A.G.; Zhelifonova, V.P.; Antipova, T.V. Fungi of the genus Penicillium as producers of physiologically active compounds (Review). Appl. Biochem. Microbiol. 2013, 49, 5–16. [Google Scholar] [CrossRef]
- Zeilinger, S. Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites. In Fungal Biology, 1st ed.; Martín, J.F., García-Estrada, C., Zeilinger, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Chapter 14; pp. 303–316. [Google Scholar]
- Du, L.; Feng, T.; Zhao, B. Alkaloids from a deep ocean sediment-derived fungus Penicillium sp. and their antitumor activities. J. Antibiot. 2010, 63, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, B.; Capon, R. Roquefortine E, a diketopiperazine from an Australian isolate of Gymnoascus reessii. J. Nat. Prod. 2005, 68, 1661–1664. [Google Scholar] [CrossRef] [PubMed]
- Ohmomo, S.; Sato, T.; Utagawa, T. Isolation of festuclavine and three new indole alkaloids, roquefortine A, B and C from cultures of Penicillium roqueforti. Agric. Biol. Chem. 1975, 39, 1333–1334. [Google Scholar] [CrossRef] [Green Version]
- Ohmomo, S.; Oguma, K.; Ohashi, T. Isolation of a new indole alkaloid, roquefortine D, from the cultures of Penicillium roqueforti. Agric. Biol. Chem. 1978, 42, 2387–2389. [Google Scholar] [CrossRef] [Green Version]
- Ries, M.I.; Ali, H.; Lankhorst, P.P. Novel key metabolites reveal further branching of the roquefortine/meleagrin biosynthetic pathway. J. Bio. Chem. 2013, 288, 37289–37295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, S.; Wang, N.; Xie, C.L. Roquefortine J, a novel roquefortine alkaloid, from the deep-sea-derived fungus Penicillium granulatum MCCC 3A00475. J. Antibiot. 2018, 71, 658–661. [Google Scholar] [CrossRef]
- Garcia-Estrada, C.; Ullan, R.V.; Albillos, S.M.; Fernandez-Bodega, M.A. A single cluster of coregulated genes encodes the biosynthesis of the mycotoxins roquefortine C and meleagrin in Penicillium chrysogenum. Chem. Biol. 2011, 18, 1499–1512. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Carroll, P.J.; Winkler, J.D. A Transannular Rearrangement Reaction of a Pyrroloindoline Diketopiperazine. Org. Lett. 2019, 21, 6619–6623. [Google Scholar] [CrossRef] [PubMed]
- Steyn, P.S. The isolation, structure and absolute configuration of secalonic acid D, the toxic metabolite of Penicillium oxalicum. Tetrahedron 1970, 26, 51–57. [Google Scholar] [CrossRef]
- Hirano, A.; Iwai, Y.; Masuma, R. Neoxaline, a new alkaloid produced by Aspergillus japonicus. Production, isolation and properties. J. Antibiot. 1979, 32, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Takeshi, Y.; Ideguchi-Matsushita, T. Asymmetric Total Synthesis of Indole Alkaloids Containing an Indoline Spiroaminal Framework. J. Chem. Eur. 2015, 21, 11855–11864. [Google Scholar] [CrossRef]
- Koizumi, Y.; Arai, M.; Tomoda, H. Oxaline, a fungal alkaloid, arrests the cell cycle in M phase by inhibition of tubulin polymerization. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2004, 1693, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reshetilova, T.A.; Vinokurova, N.G.; Khmelenina, V.N.; Kozlovsky, A.G. The role of roquefortine in the synthesis of alkaloids meleagrin, glandicolines A and B, and oxaline in fungi Penicillium glandicola and P. atramentosum. Microbio 1995, 64, 27–29. [Google Scholar]
- Gober, M.; Carroll, J.; Joullié, M. Triazaspirocycles: Occurrence, Synthesis, and Applications. Mini-Rev. Org. Chem. 2016, 13, 126–142. [Google Scholar] [CrossRef] [Green Version]
- Newmister, S.A.; Stelamar, R.; Jennifer, J.S. Unveiling sequential late-stage methyltransferase reactions in the meleagrin/oxaline biosynthetic pathway. Org. Biomol. Chem. 2018, 16, 6450–6459. [Google Scholar] [CrossRef]
- Hazrat, A.; Ries, M.I.; Nijland, J.G. A Branched Biosynthetic Pathway Is Involved in Production of Roquefortine and Related Compounds in Penicillium chrysogenum. PLoS ONE 2013, 8, e65328. [Google Scholar] [CrossRef] [Green Version]
- Kosalková, K.; Domínguez-Santos, R. A natural short pathway synthesizes roquefortine C but not meleagrin in three different Penicillium roqueforti strains. Appl. Microbiol. Biot. 2015, 99, 7601–7612. [Google Scholar] [CrossRef]
- De Faveri, R.; Nunes, R.; Santin, J.R. The role of kinins in the proliferation of fibroblast primed with TNF in scratch wound assay: Kinins and cell proliferation. Int. Immunopharmacol. 2018, 65, 23–28. [Google Scholar] [CrossRef]
- Li, Y.; Chen, D.; Su, Z. MicroRNA-106b functions as an oncogene in renal cell carcinoma by affecting cell proliferation, migration and apoptosis. Mol. Med. Rep. 2016, 13, 1420–1426. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Li, J.; Lin, Q. Ultrasonic irradiation and SonoVue microbubbles-mediated RNA interference targeting PRR11 inhibits breast cancer cells proliferation and metastasis, but promotes apoptosis. Biosci. Rep. 2020, 40, BSR20201854. [Google Scholar] [CrossRef]
- Tang, J.J.; Zhang, Y. Natural Products as Sources of New Fungicides (III): Antifungal Activity of 2,4-Dihydroxy-5-Methylacetophenone Derivatives. Bioorg. Med. Chem. Lett. 2016, 26, 2156–2158. [Google Scholar] [CrossRef]
- Kim, J.A.; Cárcer, G.D. Correction to Low Dose of Amino-Modified Nanoparticles Induces Cell Cycle Arrest. Nano 2013, 7, 7483–7494. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, M.J.; An, R.F.; Zhou, J. Diosgenin Derivatives as Potential Antitumor Agents: Synthesis, Cytotoxicity, and Mechanism of Action. J. Nat. Prod. 2020, 84, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Katsuyama, Y. Mining novel biosynthetic machineries of secondary metabolites from actinobacteria. Biosci. Biotech. Bioch. 2019, 83, 1606–1615. [Google Scholar] [CrossRef]
- Gaytan-Graham, S. Hepatocellular carcinoma: An update. Ultrastruct. Pathol. 2001, 25, 497–516. [Google Scholar] [CrossRef]
- Mady, M.S.; Mohyeldin, M.M.; Ebrahim, H.Y. The indole alkaloid meleagrin, from the olive tree endophytic fungus Penicillium chrysogenum, as a novel lead for the control of c-Met-dependent breast cancer proliferation, migration and invasion. Bioorg. Med. Chem. 2016, 24, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Li, D.H.; Zhu, T.J.; Wang, F.P. GuNew alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Size (bp) | Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|
| RDS-F | 21 | AGCTATGGCTTTGATAGCAGC | 567 |
| RDS-R | 21 | CACCAGATCGCCGGTTTTATA | |
| RPT-F | 20 | AGCGTGGAACTGAGCCAGAA | 567 |
| RPT-R | 21 | CCAATGTTCTTCCACTTTCGC | |
| SRO-F | 20 | TGACCACGCGAGCATTTGTC | 501 |
| SRO-R | 20 | TCATCACGCTCTCCATGAGT |
| Cell line | A549 | MCF-7 | P6C | K562 | L-02 |
| meleagrin | >10 | 4.94 ± 0.001 | >10 | >10 | >10 |
| oxaline | 6.85 ± 0.03 | >10 | 9.41 ± 0.01 | 7.8 ± 0.02 | >10 |
| Adriamycin | 0.26 ± 0.006 | 0.82 ± 0.014 | 0.4 ± 0.001 | 0.92 ± 0.04 | 0.21 ± 0.01 |
| Cell line | HCT-116 | HePG2 | |||
| meleagrin | 5.7 ± 0.013 | 1.82 ± 0.021 | |||
| oxaline | 4.94 ± 0.001 | 4.27 ± 0.01 | |||
| Adriamycin | 0.21 ± 0.01 | 0.04 ± 0.002 |
| Medium | Medium Constituents |
|---|---|
| SWS | peptone 0.1%, soluble starch 1%, seawater |
| 1# | sorbic alcohol 2%, maltose 2%, monosodium glutamate 1%, KH2PO4 0.05%, MgSO4 7H2O 0.03%, tryptophan 0.05%, yeast extract 0.3%, seawater |
| 2# | monosodium glutamate 1%, mannitol 2%, maltose 2%, KH2PO4 0.05%, glucose 1%, corn extract 0.1%, MgSO4 7H2O 0.03%, yeast extract 0.3%, seawater |
| 5# | glucose 2.0%, peptone 1%, malt leaching powder 0.3%, yeast extract 0.3%, seawater |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Jin, Y.; Kong, F.; Yang, L.; Zhu, M.; Wang, Y. Discovery, Antitumor Activity, and Fermentation Optimization of Roquefortines from Penicillium sp. OUCMDZ-1435. Molecules 2023, 28, 3180. https://doi.org/10.3390/molecules28073180
He X, Jin Y, Kong F, Yang L, Zhu M, Wang Y. Discovery, Antitumor Activity, and Fermentation Optimization of Roquefortines from Penicillium sp. OUCMDZ-1435. Molecules. 2023; 28(7):3180. https://doi.org/10.3390/molecules28073180
Chicago/Turabian StyleHe, Xingxing, Yanzheng Jin, Fei Kong, Liyuan Yang, Mingzhuang Zhu, and Yi Wang. 2023. "Discovery, Antitumor Activity, and Fermentation Optimization of Roquefortines from Penicillium sp. OUCMDZ-1435" Molecules 28, no. 7: 3180. https://doi.org/10.3390/molecules28073180