
Deciphering Complex Interactions in Bioactive Lipid Signaling


1
Department of Biotechnological and Applied Clinical Sciences, University of L’Aquila, 67100 L’Aquila, Italy
2
European Center for Brain Research (CERC), Santa Lucia Foundation IRCCS, 00143 Rome, Italy
Molecules 2023, 28(6), 2622; https://doi.org/10.3390/molecules28062622
Submission received: 23 February 2023
/
Revised: 10 March 2023
/
Accepted: 11 March 2023
/
Published: 14 March 2023
(This article belongs to the Special Issue Bioactive Lipids in Inflammatory Diseases)
Abstract
:Lipids are usually viewed as metabolic fuel and structural membrane components. Yet, in recent years, different families of lipids able to act as authentic messengers between cells and/or intracellularly have been discovered. Such lipid signals have been shown to exert their biological activity via specific receptors that, by triggering distinct signal transduction pathways, regulate manifold pathophysiological processes in our body. Here, endogenous bioactive lipids produced from arachidonic acid (AA) and other poly-unsaturated fatty acids will be presented, in order to put into better perspective the relevance of their mutual interactions for health and disease conditions. To this end, metabolism and signal transduction pathways of classical eicosanoids, endocannabinoids and specialized pro-resolving mediators will be described, and the intersections and commonalities of their metabolic enzymes and binding receptors will be discussed. Moreover, the interactions of AA-derived signals with other bioactive lipids such as shingosine-1-phosphate and steroid hormones will be addressed.
1. Introduction
Lipids are usually viewed as metabolic fuel and structural membrane components. Yet, in recent years, different families of lipids able to act as authentic messengers between cells and/or intracellularly have been discovered and have been shown to exert their biological activity by binding to specific receptors, thus triggering signal transduction pathways that regulate manifold pathophysiological processes in our body.
In general, bioactive lipids in cell signaling include plant-derived compounds such as cannabinoids (terpeno-phenol compounds also known as phytocannabinoids) [1,2], terpenoids (including sterols) [3,4], carotenoids and phenolics (including flavonoids) [5,6]. In addition, our cells produce lipids endowed with biological activity in signal transduction, the best characterized being derivatives of poly-unsaturated fatty acids (PUFAs). These endogenous substances support a complex network of molecular and cellular events responsible for a plethora of physiopathological conditions such as inflammation and chronic diseases, cancer, diabetes, atherosclerosis, asthma, autoimmune and neurodegenerative disorders [7], as well as reproduction [8,9], just to mention a few.
To date, the three major classes of endogenous bioactive lipids that have been best characterized are classical eicosanoids (EICs), endocannabinoids (ECBs) and specialized pro-resolving mediators (SPMs) [7]. A PubMed search conducted over the last 30 years clearly shows that interest in these substances has always remained high for EICs, the first to be discovered in the 1930s [10], and has been continuously growing for ECBs and SPMs since their first identification in 1992 [11] and 1974 [12], respectively (Table 1).
In order to put into better perspective the relevance of the potential interactions of different bioactive lipids for health and disease conditions, the metabolism and signal transduction pathways of these compounds are summarized below.
2. Bioactive Lipids
2.1. Eicosanoids
Eicosanoids (EICs) are a family of ~120 biologically active compounds derived from unsaturated fatty acids, among which n-6 (ω-6) arachidonic acid (eicosatetraenoic acid, C20:4) is the main precursor. The history of the EICs dates back to the 1930s, when new biologically active lipids were found in human seminal plasma [10]. Since then, they have been shown to play a major role in key processes like inflammation [13] and maintenance of energy homeostasis [14,15,16,17].
Upon specific stimuli, phospholipase A2 (PLA2) releases arachidonic acid (AA) from the sn-2 position of membrane glycerophosholipids [18,19], and then EICs are generated through three main biosynthetic pathways, driven by (i) cyclooxygenases (COXs) [20], (ii) lipoxygenases (LOXs) [21], and (iii) cytochrome P450 epoxydases (CYP450s) [22].
Cyclooxygenase-1 (COX-1) and -2 (COX-2) generate EICs that have a ring in their structure and are collectively termed prostanoids; the latter include prostaglandins (PGs), prostacyclins (PGIs) and thromboxanes (TXs) that are all formed from the common precursor prostaglandin H2 (PGH2). All prostanoids bind to and activate distinct G protein-coupled receptors (GPCRs), as summarized in Figure 1, and thus, they exert well-recognized hormone-like actions on the surrounding cells or the producing cells in a paracrine or autocrine manner [20]. Consequently, prostanoids trigger signal transduction on various human cells such as neuron, retina, vessel, heart, and blood cells.
Different lipoxygenase (LOX) isozymes such as 5-LOX, 12-LOX and 15-LOX promote the conversion of AA into hydroperoxy derivatives (hydroperoxyeicosatetraenoic acids, HpETEs). 5-HpETE is a precursor of leukotrienes LTA4 and LTB4, as well as of cysteinyl-LTs (CysLTs) such as LTC4, LTD4 and LTE4; 12-HpETE is a precursor of hepoxilins (HxA and HxB); and 15-HpETE is a precursor of lipoxins (LXA4 and LXB4) [21]. Of note, due to their pro-resolving properties, lipoxins are generally also listed among SPMs, detailed in the following Section 2.3. Much like COX-generated compounds, LOX-derived EICs bind to distinct GPCRs, some of which have been identified. LTB4 binds to and activates BLT1 and BLT2 receptors, whereas CysLTs bind to and activate CyLTR1 and CysLTR2 receptors [23].
2.2. Endocannabinoids
The endocannabinoids (ECBs) bind to and activate the same cannabinoid 1 (CB1) and 2 (CB2) GPCRs activated by Δ9-tetrahydrocannabinol (THC), the main psychoactive component of cannabis (Cannabis sativa and Cannabis indica) [24,25]. Therefore, ECBs exert a wide range of pathophysiological functions, both in the central nervous system and in peripheral tissues, generally acting as pro-homeostatic modulators [26]. The main ECBs so far identified are N-arachidonoylethanolamine or anandamide (AEA), which is an amide of AA with ethanolamine [11], and 2-arachidonoylglycerol (2-AG), which is an ester of AA with glycerol [27]. AEA is produced mainly via N-acylphosphatidylethanolamines-specific phospholipase D (NAPE-PLD) [28], and 2-AG mainly via sn-1-diacylglycerol lipases (DAGLs) α and β [29]. Yet, additional biosynthetic enzymes can contribute to AEA and 2-AG production, such as cytosolic phospholipase A2γ [30], phospholipase C [31] and the αβ-hydrolase domain 4 (ABHD4) protein [32]. Then, degradation of AEA is primarily catalyzed by fatty acid amide hydrolase (FAAH), which cleaves it into AA and ethanolamine [33]. Instead, 2-AG is cleaved by monoacylglycerol lipase (MAGL) that releases AA and glycerol [34]. Much like the biosynthesis, the degradation of AEA and 2-AG also can occur via additional hydrolases or can be paralleled by alternative pathways such as oxygenation through COX-2, 12-LOX, 15-LOX, or CYP450s [35]. The latter oxygenases generate ECB-like derivatives known as PG-ethanolamides, glyceryl esters, hydroxy-AEAs and hydroxy-eicosatetraenoylglycerols. These oxidized compounds seem to have their own biological activity by activating distinct receptor targets, yet to be identified [26,36]. Indeed, both AEA and 2-AG bind to and activate Gi/o protein-coupled CB1 and CB2 receptors [37,38], along with additional ECB-binding receptors, such as the orphan G protein-coupled receptors GPR55 and GPR119, the transient receptor potential vanilloid type-1 (TRPV1) ion channel, and the nuclear peroxisome proliferator-activated receptors (PPAR) α, γ and δ [25]. Thus, AEA and 2-AG, in addition to congeners such as N-archidonoyldopamine (NADA) and N-oleoyldopamine (OLDA), are also considered bona fide endovanilloids [39]. Incidentally, there are ECB-like compounds, such as N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA), and amphiphilic molecules, such as N-acyl amino acids (e.g., N-arachidonoylglycine), that also share enzymatic pathways and/or interactions with receptors of the ECBs, thus modulating various physiological and pathological conditions [25].
The metabolic pathways and known receptors of eCBs, that together all form the so-called “endocannabinoid system” are summarized in Figure 2.
2.3. Specialized Pro-Resolving Mediators
The specialized pro-resolving mediators (SPMs) had been identified in 1974 as anti-inflammatory lipids [12]. Indeed, their name refers to their major role as drivers of resolution of inflammation. Unlike classical EICs that are produced mainly from n-6 (ω-6) AA, almost all SPMs derive from n-3 (ω-3) PUFAs, with only a few compounds sharing with EICs the n-6 (ω-6) AA precursor [40]. Depending on the PUFAs used for the synthesis, SPMs are classified as derivatives of (i) docosahexaenoic acid (DHA), (ii) docosapentaenoic acid (DPA), (iii) eicosapentaenoic acid (EPA), and (iv) AA.
The biosynthesis of SPMs resembles that of EICs, because it starts with the PLA2-catalyzed release of PUFAs from membrane phospholipids, followed by further processing via COX-2 and/or various LOX and CYP450 isozymes.
DHA-derived SPMs are the largest class of SPMs and include heterogeneous D-series resolvins 1–6 (RvD 1–6), maresins 1,2 (MaR1 and MaR2), and neuroprotectins/protectins 1, X (PD1/NPD1 and PDX). 12-LOX or 15-LOX generate the 14(S)- or 17(S)-hydroperoxides of DHA (14(S)-HpDHA and 17(S)-HpDHA), respectively. Then, maresins are obtained through further metabolism of 14(S)-HpDHA, whereas RvDs and PDs/NPDs are generated from 17(S)-HpDHA. To date, the following GPCRs have been identified as targets of DHA-derived SPMs: formyl peptide receptor 2 (FPR2)—also known as ALX (FPR2/ALX)—a target of RvD1 and RvD3; GPR18, a target of RvD2; GPR32, a target of RvD1, RvD3 and RvD5; GPR37, a target of PD1; and Leucine-rich repeat-containing G protein-coupled receptor 6 (LGR6), a target of MaR1 [40].
DPA-derived SPMs are a newly discovered class of SPMs that include 13-series resolvins (RvTs) as well as the DPA-derived counterparts of DHA SPMs. The metabolic enzymes that convert DPA into SPMs are, again, COX-2, 12-LOX and 15-LOX [41,42].
EPA-derived SPMs are a small group that includes E-series resolvins 1–3 (RvE1–3). Upon release of EPA from membrane phospholipids, COX-2 or CYP450 promote its conversion to 18(R/S)-hydroperoxyeicosapentaenoic acid (18(R/S)-HpEPE) epimers that are then further metabolized through 5/12/15-LOX, peroxidases and hydrolases. As yet, only one receptor has been shown to be activated by RvEs, and it is termed the chemerin receptor 23 (ChemR23). Yet, LTB4-binding BLT1 receptor also can be antagonized by both RvE1 and RvE2 [43].
3. Interactions in Lipid Signaling Pathways
3.1. Common Metabolic Pathways
From the metabolic pathways of EICs, ECBs and SPMs shown in Figure 1, Figure 2 and Figure 3, it is apparent that these families of bioactive lipids share key enzymes, such as the lipase responsible for the release from phospholipid precursors (PLA2) and three oxygenases able to generate from AA: (i) epoxyeicosatrienoic acids (CYP450); (ii) 12-hydroxyeicosatetraenoic acids (12-LOX); and (iii) cyclic prostanoid(-like) oxidative derivatives (COX-2). Of note, PLA2 belongs to a superfamily of enzymes that play major roles in several pathophysiological processes, e.g., mast cell biology [45], atherosclerotic cardiovascular disease [46], and gastrointestinal diseases [47]. Unsurprisingly, PLA2 enzymes are able to mobilize unique bioactive lipids in multiple ways that are spatiotemporally regulated, thus triggering distinct signaling pathways. In addition, an emerging concept in the control of the synthesis of oxygenated metabolites of AA—as well as other PUFAs—is the compartmentalization of the enzymes and lipid substrates within the cell, a clue that allows shaping the appropriate biological response in different cellular contexts [48]. Unfortunately, the molecular mechanisms that underlie the fine tuning of these enzymes, and their ability to drive signaling through one class of lipids or another, remain elusive. Yet, substrate hydrolysis has been carefully interrogated in the case of COX-2, by comparing oxygenation of AA to prostaglandins (PGs) and of the AA ester 2-AG to prostaglandin glyceryl esters (PG-Gs) [49]. The efficiency of in vitro oxygenation of these two substrates by COX-2 has been found to be similar, but competition between AA and 2-AG at the catalytic and allosteric sites of the enzyme (a dimer), combined with a differential allosteric modulation, leads to preferential oxygenation of AA in vivo [49]. This is just one example, thoroughly investigated, of the complex interplay between different bioactive lipids and their metabolic enzymes, with major functional consequences for the engagement of downstream targets that in turn trigger a biological activity [35]. Common enzymes between EICs, ECBs and SPMs are summarized in Table 2.
Besides enzymes, protein carriers [50] or microvesicles [51] that transport bioactive lipids inside and outside the cell can also be expected to contribute to fueling one metabolic route rather than another, as it is becoming apparent in the case of ECBs [52,53,54]. Unfortunately, for known lipid transporters such as heat shock protein 70 and albumin [55], fatty acid-binding proteins 1, 5 and 7 [56,57], FAAH-like anandamide transporter [58], sterol carrier protein 2 [59] and retinol-binding protein 2 [60], the mechanisms that support the choice of one substrate over another also remain to be elucidated. At any rate, crosstalk among different bioactive lipids are emerging in the literature. For instance, RvD1 has been shown to produce potent antiallodynia and antihyperalgesia in a model of bone cancer pain by also triggering spinal upregulation of ECBs, which produce additional antinociception predominantly through CB2 receptors [61]. Moreover, activation of the DHA-derived SPM-binding receptor GPR18 has been shown to drive neuroprotection against HIV-1 Tat-induced neurodegeneration by engaging FAAH, the key enzyme for AEA hydrolysis [62]. The same GPR18 receptor has been also shown to modulate ECB signaling in metabolic dysfunction, as well as in microglia [63]. Incidentally, it should be noted that more subtle interactions may also occur directly between lipid metabolic enzymes, as in the case of 5-LOX that blocks FAAH activity in human mast cells by producing AEA hydroperoxides that act as enzyme inhibitors [64].
3.2. Common Receptor Targets
It is remarkable that almost all receptors of bioactive lipids are G protein-coupled receptors (GPCRs), the exceptions being the ECB-binding TRPV1 channels and PPARα/γ/δ nuclear receptors (Figure 1, Figure 2 and Figure 3). GPCRs are one of the most important classes of membrane receptors, able to sense an extraordinary variety of biomolecules in order to activate multiple intracellular signaling cascades [65]. An unresolved issue about GPCRs is signal bias (also known as biased signaling, biased agonism or functional selectivity), whereby a ligand-dependent selectivity exists for certain signal transduction pathways compared to a reference ligand (often the endogenous agonist) at the same receptor. Biased agonists for GPCRs are of profound pharmacological interest, because they could usher in a new generation of drugs with greatly reduced side effects [66]. An interesting example of the alternative pathways that can be activated by different ligands at the same receptor, and by the same ligand at different receptors, is represented by ECB signaling [25], schematically depicted in Figure 4.
Unlike metabolic pathways that share some of the enzymes responsible for synthesis and degradation (Table 2), receptors seem to be clearly distinct for EICs, ECBs and SPMs (Figure 1, Figure 2 and Figure 3). However, emerging evidence supports the possibility that one class of bioactive lipids can bind to and activate specific receptors of another class, adding a further dimension to the complexity of the transduction pathways of these molecules. Indeed, the endogenous ECB-like compound N-arachidonoylglycine has been recently shown to activate the SPM-binding receptor GPR18 with neuroprotective effects [67], and so does the plant-derived substance cannabidiol, with an impact on Alzheimer’s disease [68] and, apparently more in general, on the balance between different lipid signals. Additionally, the endogenous ECB-like compound, N-palmitoylethanolamine (PEA), has been shown to promote a pro-resolving macrophage phenotype [69], and selective activation of CB2 improves efferocytosis, which is a typical hallmark of resolution of inflammation [70]. Therefore, GPR18 and CB2 seem to be a cross-roads for the action of ECBs and SPMs, apparently also due to the possible formation of heterodimers [71]. GPR18-CB2 heteroreceptor complexes display particular functional properties that often consist of negative crosstalk (i.e., activation of one receptor reduces signaling that arises from the partner receptor) and cross-antagonisms (i.e., the response of one of the receptors is blocked by a selective antagonist of the partner receptor). Once again, this is just one example of the complexity of the crosstalk between different classes of bioactive lipids, which might contribute to biased signaling triggered by each of them [72].
Additionally, lipoxin A4 (LXA4), an anti-inflammatory EIC/SPM, has been shown to act as an endogenous allosteric enhancer of CB1 receptor [73], able to reinforce the biological activity of AEA [74]. Moreover, the peroxidation product 12(S)-hydroxyeicosatetraenoic acid, generated by 12-LOX from AA (Figure 1), has been shown to mediate diabetes-induced endothelial dysfunction by activating TRPV1 [75], leaving open the possibility that the latter receptor channel may be a target not only of ECBs, but also of EICs and SPMs. In this context, LTB4-binding BLT1 receptor can be antagonized by both RvE1 and RvE2 [43], thus bridging together the actions of EICs and SPMs (Table 2).
On a final note, it should be stressed that a pre-requisite to design effective and selective drugs able to modulate endogenous lipid signaling is the clear understanding of the 3D structures of the molecular targets. This knowledge is largely available for the metabolic enzymes and binding receptors of EICs, a lipid class known for quite a few decades [10]. Surprisingly, for the molecular machinery that drives ECB signaling, to date, 3D structures of only 23 major components have been resolved, whereas many other receptors (e.g., GPR55, GPR119 and TRPV4), enzymes (e.g., NAT, DAGLα/β, GDE1,4,7, ABHD2,4,6,12) and putative membrane transporters still await clarification [26]. Finally, for SPMs, only the 3D structures of formyl peptide receptor 2 (FPR2) [76] and leukotriene B4 receptor 1 (BLT1) [77] have been recently elucidated, making it difficult to thoroughly investigate the mechanisms of binding, activation and regulation of these GPCRs. It is apparent that such an information gap is particularly troubling for drug discovery programs and must be urgently filled.
3.3. Interaction of EICs, ECBs and SPMs with AA/PUFA-Unrelated Bioactive Lipids
In addition to the above-discussed interactions among AA/PUFA-derived EICs, ECBs and SPMs, additional bioactive lipids can crosstalk with them. For instance, sphingolipids are a relevant class of bioactive lipids that play key roles in the regulation of several cellular processes, including growth, differentiation, inflammatory responses, and apoptosis. Recent studies implicated a regulatory function of sphingolipids in prostaglandin production, and sphingosine-1-phosphate (S1P) has been shown to induce COX-2 expression [78]. In line with this, S1P has been recently demonstrated to activate TRPV1, with an impact on AEA-dependent regulation of mitochondrial activity [79]; conversely, RvD1 activates the S1P signaling pathway [80]. Moreover, steroid hormones also can contribute to modulating AA/PUFA-derived lipid signaling, for instance, at the level of gene expression. Such a regulation has been indeed shown for estrogen that enhances FAAH expression via an epigenetic mechanism mediated by histone demethylase LSD1 [81], and for progesterone that activates the promoter of the same AEA hydrolase through the transcription factor Ikaros [82]. These findings open new perspectives for the development of epigenetic drugs (epidrugs) able to modulate bioactive lipid signaling [83]. Overall, these data support a further layer of complexity in bioactive lipid crosstalk, well beyond the family of AA/PUFA-derived molecules.
4. Conclusions and Future Directions
In recent years, a diverse set of signaling lipids has been demonstrated to interact with specific receptors under a tight metabolic control, with an impact on many pathophysiological processes. Unlike protein signals that can be stored in vesicles, lipid messengers are often synthesized “on demand”, i.e., when and where needed, and their endogenous content is tightly regulated by distinct biosynthetic and hydrolytic enzymes. Understanding mutual regulations of these enzymes, possibly through post-translational modifications, protein–protein and protein–lipid interactions, as well as via membrane and subcellular location, now seems urgent in order to exploit lipid signals as potential next-generation therapeutics. In particular, it should be recalled that location is a critical element to modulating metabolic enzymes and receptors, and hence to driving signaling. In line with this, the possibility of visualizing intracellular trafficking of lipid-interacting proteins seems relevant, making necessary and urgent the development and use of chemical probes. Only recently have the development of small-molecule inhibitors and activity-based probes, along with the use of advanced techniques like activity-based protein profiling and chemical proteomics, been essential to guide the drug discovery and development of compounds able to target lipid signaling, for instance allowing to visualize key elements of ECB signaling, such as FAAH and MAGL hydrolases [84] and CB2 receptor [85]. Now that tools for more accurately tracking lipid metabolism, location, and action have been provided, and that innovations in bioimaging techniques have been made, it can be anticipated that the underlying mechanisms of lipid signaling and of the crosstalk among different lipid classes will be uncovered at nanoscale resolution, thus allowing to interrogate lipid networks in real life.
Funding
This research was funded by the Italian Ministry of Health under the competitive grant RF-2018-12365391.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
I wish to thank Annamaria Tisi (University of L’Aquila, Italy) for kindly preparing the artwork.
Conflicts of Interest
The author declares no conflict of interest.
References
- Abd-Nikfarjam, B.; Dolati-Somarin, A.; Baradaran Rahimi, V.; Askari, V.R. Cannabinoids in neuroinflammatory disorders: Focusing on multiple sclerosis, Parkinson’s, and Alzheimer’s diseases. Biofactors, 2023; in press. [Google Scholar] [CrossRef] [PubMed]
- Ang, S.P.; Sidharthan, S.; Lai, W.; Hussain, N.; Patel, K.V.; Gulati, A.; Henry, O.; Kaye, A.D.; Orhurhu, V. Cannabinoids as a potential alternative to opioids in the management of various pain subtypes: Benefits, limitations, and risks. Pain Ther. 2023; in press. [Google Scholar] [CrossRef]
- Bhaswant, M.; Shanmugam, D.K.; Miyazawa, T.; Abe, C.; Miyazawa, T. Microgreens-A comprehensive review of bioactive molecules and health benefits. Molecules 2023, 28, 867. [Google Scholar] [CrossRef] [PubMed]
- Mulyani, Y.; Sinaga, S.E.; Supratman, U. Phytochemistry and biological activities of endophytic fungi from the meliaceae family. Molecules 2023, 28, 778. [Google Scholar] [CrossRef] [PubMed]
- Duan, N.; Hu, X.; Zhou, R.; Li, Y.; Wu, W.; Liu, N. A review on dietary flavonoids as modulators of the tumor microenvironment. Mol. Nutr. Food Res. 2023, 2023, e2200435. [Google Scholar] [CrossRef]
- Kumar, A.P.N.; Kumar, M.; Jose, A.; Tomer, V.; Oz, E.; Proestos, C.; Zeng, M.; Elobeid, T.K.S.; Oz, F. Major phytochemicals: Recent advances in health benefits and extraction method. Molecules 2023, 28, 887. [Google Scholar] [CrossRef]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive lipids, inflammation and chronic diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef]
- Cecconi, S.; Rapino, C.; Di Nisio, V.; Rossi, G.; Maccarrone, M. The (endo)cannabinoid signaling in female reproduction: What are the latest advances? Prog. Lipid Res. 2020, 77, 101019. [Google Scholar] [CrossRef]
- Maccarrone, M.; Rapino, C.; Francavilla, F.; Barbonetti, A. Cannabinoid signalling and effects of cannabis on the male reproductive system. Nat. Rev. Urol. 2021, 18, 19–32. [Google Scholar] [CrossRef]
- Baker, R.R. The eicosanoids: A historical overview. Clin. Biochem. 1990, 23, 455–458. [Google Scholar] [CrossRef]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Systems approach to inflammation resolution: Identification of novel anti-inflammatory and pro-resolving mediators. J. Thromb. Haemost. 2009, 7, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 158, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzawa, H.; Kohno, D.; Koga, T.; Sasaki, T.; Fukunaka, A.; Okuno, T.; Jo-Watanabe, A.; Kazuno, S.; Miyatsuka, T.; Kitamura, T. Leukotriene A4 hydrolase deficiency protects mice from diet-induced obesity by increasing energy expenditure through neuroendocrine axis. FASEB J. 2020, 34, 13949–13958. [Google Scholar] [CrossRef]
- Félix-Soriano, E.; Sáinz, N.; Gil-Iturbe, E.; Collantes, M.; Fernández-Galilea, M.; Castilla-Madrigal, R.; Ly, L.; Dalli, J.; Moreno-Aliaga, M.J. Changes in brown adipose tissue lipid mediator signatures with aging, obesity, and DHA supplementation in female mice. FASEB J. 2021, 35, e21592. [Google Scholar] [CrossRef]
- Fujimori, K.; Uno, S.; Kuroda, K.; Matsumoto, C.; Maehara, T. Leukotriene C4 synthase is a novel PPARγ target gene, and leukotriene C4 and D4 activate adipogenesis through cysteinyl LT1 receptors in adipocytes. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119203. [Google Scholar] [CrossRef]
- Eckenstaler, R.; Ripperger, A.; Hauke, M.; Petermann, M.; Hemkemeyer, S.A.; Schwedhelm, E.; Ergün, S.; Frye, M.; Werz, O.; Koeberle, A. A thromboxane A2 receptor-driven COX-2-dependent feedback loop that affects endothelial homeostasis and angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 444–461. [Google Scholar] [CrossRef]
- Leslie, C.C. Cytosolic phospholipase A2: Physiological function and role in disease. J. Lipid Res. 2015, 56, 1386–1402. [Google Scholar] [CrossRef] [Green Version]
- Mouchlis, V.D.; Dennis, E.A. Phospholipase A2 catalysis and lipid mediator lipidomics. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 766–771. [Google Scholar] [CrossRef]
- Smith, W.L.; Urade, Y.; Jakobsson, P. Enzymes of the cyclooxygenase pathways of prostanoid biosynthesis. Chem. Rev. 2011, 111, 5821–5865. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.R.; Guy, C.J.; Scurr, M.J.; Taylor, P.R.; Kift-Morgan, A.P.; Hammond, V.J.; Thomas, C.P.; Coles, B.; Roberts, G.W.; Eberl, M.; et al. Esterified eicosanoids are acutely generated by 5-lipoxygenase in primary human neutrophils and in human and murine infection. Blood 2011, 117, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Roman, R.J. Effect of cytochrome P450 metabolites of arachidonic acid in nephrology. J. Am. Soc. Nephrol. 2017, 28, 2845–2855. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, T.; Nakamura, M.; Shimizu, T. Leukotriene receptors as potential therapeutic targets. J. Clin. Invest. 2018, 128, 2691–2701. [Google Scholar] [CrossRef]
- El Sohly, M.A.; Radwan, M.M.; Gul, W. Phytochemistry of Cannabis sativa L. Prog. Chem. Org. Nat. Prod. 2017, 103, 1–36. [Google Scholar] [PubMed]
- Gómez-Cañas, M.; Rodríguez-Cueto, C.; Satta, V.; Hernández-Fisac, I.; Navarro, E.; Fernández-Ruiz, J. Endocannabinoid-binding receptors as drug targets. Methods Mol. Biol. 2023, 2576, 67–94. [Google Scholar] [PubMed]
- Maccarrone, M. Missing pieces to the endocannabinoid puzzle. Trends Mol. Med. 2020, 26, 263–272. [Google Scholar] [CrossRef]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular characterization of a phospholipase D generating anandamide and its congeners. J. Biol. Chem. 2004, 279, 5298–5305. [Google Scholar] [CrossRef] [Green Version]
- Bisogno, T.; Howell, F.; Williams, G.; Minassi, A.; Cascio, M.G.; Ligresti, A.; Matias, I.; Schiano-Moriello, A.; Paul, P.; Williams, E.J.; et al. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. J. Cell Biol. 2003, 163, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Uyama, T.; Rahman, S.M.K.; Sikder, M.M.; Hussain, Z.; Tsuboi, K.; Miyake, M.; Ueda, N. Involvement of the γ isoform of cPLA2 in the biosynthesis of bioactive N-acylethanolamines. Molecules 2021, 26, 5213. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Harvey-White, J.; Osei-Hyiaman, D.; Razdan, R.; Gong, Q.; Chan, A.C.; Zhou, Z.; Huang, B.X.; Kim, H.Y. A biosynthetic pathway for anandamide. Proc. Natl. Acad. Sci. USA 2006, 103, 13345–13350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, G.M.; Cravatt, B.F. Endocannabinoid biosynthesis proceeding through glycerophospho-N-acyl ethanolamine and a role for alpha/beta-hydrolase 4 in this pathway. J. Biol. Chem. 2006, 281, 26465–26472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cravatt, B.F.; Giang, D.K.; Mayfield, S.P.; Boger, D.L.; Lerner, R.A.; Gilula, N.B. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996, 384, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.P.; Carpenter, D.; Leslie, F.M.; Freund, T.F.; Katona, I.; Sensi, S.L.; Kathuria, S.; Piomelli, D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA 2002, 99, 10819–10824. [Google Scholar] [CrossRef] [Green Version]
- Rouzer, C.A.; Marnett, L.J. Structural and chemical biology of the interaction of cyclooxygenase with substrates and non-steroidal anti-inflammatory drugs. Chem. Rev. 2020, 120, 7592–7641. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Joffre, J.; Wong, E.; Lawton, S.; Lloyd, E.; Nguyen, N.; Xu, F.; Sempio, C.; Kobzik, L.; Zlatanova, I.; Schumacher, M.; et al. N-Oleoyl dopamine induces IL-10 via central nervous system TRPV1 and improves endotoxemia and sepsis outcomes. J. Neuroinflamm. 2022, 19, 118. [Google Scholar] [CrossRef]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Colas, R.A.; Serhan, C.N. Novel n-3 immunoresolvents: Structures and actions. Sci. Rep. 2013, 3, 1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalli, J.; Chiang, N.; Serhan, C.N. Elucidation of novel 13-series resolvins that increase with atorvastatin and clear infections. Nat. Med. 2015, 21, 1071–1075. [Google Scholar] [CrossRef] [Green Version]
- López-Vicario, C.; Rius, B.; Alcaraz-Quiles, J.; García-Alonso, V.; Lopategi, A.; Titos, E.; Clària, J. Pro-resolving mediators produced from EPA and DHA: Overview of the pathways involved and their mechanisms in metabolic syndrome and related liver diseases. Eur. J. Pharmacol. 2016, 785, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Chiang, N.; Serhan, C.N. Structural elucidation and physiologic functions of specialized pro-resolving mediators and their receptors. Mol. Aspects Med. 2017, 58, 114–129. [Google Scholar] [CrossRef] [PubMed]
- Taketomi, Y.; Murakami, M. Regulatory roles of phospholipase A2 enzymes and bioactive lipids in mast cell biology. Front. Immunol. 2022, 13, 923265. [Google Scholar] [CrossRef]
- Pantazi, D.; Tellis, C.; Tselepis, A.D. Oxidized phospholipids and lipoprotein-associated phospholipase A2 (Lp-PLA2) in atherosclerotic cardiovascular disease: An update. Biofactors 2022, 48, 1257–1270. [Google Scholar] [CrossRef]
- Wu, W.; Li, W.X.; Huang, C.H. Phospholipase A2, a nonnegligible enzyme superfamily in gastrointestinal diseases. Biochimie 2022, 194, 79–95. [Google Scholar] [CrossRef]
- Astudillo, A.M.; Balboa, M.A.; Balsinde, J. Compartmentalized regulation of lipid signaling in oxidative stress and inflammation: Plasmalogens, oxidized lipids and ferroptosis as new paradigms of bioactive lipid research. Prog. Lipid Res. 2023, 89, 101207. [Google Scholar] [CrossRef]
- Mitchener, M.M.; Hermanson, D.J.; Shockley, E.M.; Brown, H.A.; Lindsley, C.W.; Reese, J.; Rouzer, C.A.; Lopez, C.F.; Marnett, L.J. Competition and allostery govern substrate selectivity of cyclooxygenase-2. Proc. Natl. Acad. Sci. USA 2015, 112, 12366–12371. [Google Scholar] [CrossRef] [Green Version]
- Verderio, C.; Gabrielli, M.; Giussani, P. Role of sphingolipids in the biogenesis and biological activity of extracellular vesicles. J. Lipid Res. 2018, 59, 1325–1340. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, M.; Battista, N.; Riganti, L.; Prada, I.; Antonucci, F.; Cantone, L.; Matteoli, M.; Maccarrone, M.; Verderio, C. Active endocannabinoids are secreted on extracellular membrane vesicles. EMBO Rep. 2015, 16, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Li, H.; Tang, C.K. Sterol carrier protein 2 in lipid metabolism and non-alcoholic fatty liver disease: Pathophysiology, molecular biology, and potential clinical implications. Metabolism 2022, 131, 155180. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Dryanovski, D.I.; Kimura, Y.; Jackson, S.N.; Woods, A.S.; Yasui, Y.; Tsai, S.Y.; Patel, S.; Covey, D.P.; Su, T.P.; et al. Cocaine-induced endocannabinoid signaling mediated by sigma-1 receptors and extracellular vesicle secretion. eLife 2019, 8, e47209. [Google Scholar] [CrossRef]
- Marion, M.; Hamilton, J.; Richardson, B.; Roeder, N.; Figueiredo, A.; Nubelo, A.; Hetelekides, E.; Penman, S.; Owada, Y.; Kagawa, Y.; et al. Environmental enrichment sex-dependently rescues memory impairment in FABP5 KO mice not mediated by brain-derived neurotrophic factor. Behav. Brain Res. 2022, 425, 113814. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Fezza, F.; Pasquariello, N.; D’Agostino, A.; Catanzaro, G.; De Simone, C.; Rapino, C.; Finazzi-Agrò, A.; Maccarrone, M. Molecular identification of albumin and Hsp70 as cytosolic anandamide-binding proteins. Chem. Biol. 2009, 16, 624–632. [Google Scholar] [CrossRef]
- Kaczocha, M.; Glaser, S.T.; Deutsch, D.G. Identification of intracellular carriers for the endocannabinoid anandamide. Proc. Natl. Acad. Sci. USA 2009, 106, 6375–6380. [Google Scholar] [CrossRef] [Green Version]
- Elmes, M.W.; Prentis, L.E.; McGoldrick, L.L.; Giuliano, C.J.; Sweeney, J.M.; Joseph, O.M.; Che, J.; Carbonetti, G.S.; Studholme, K.; Deutsch, D.G.; et al. FABP1 controls hepatic transport and biotransformation of D9-THC. Sci. Rep. 2019, 9, 7588. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Bottegoni, G.; Sasso, O.; Bertorelli, R.; Rocchia, W.; Masetti, M.; Guijarro, A.; Lodola, A.; Armirotti, A.; Garau, G.; et al. A catalytically silent FAAH-1 variant drives anandamide transport in neurons. Nat. Neurosci. 2011, 15, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Hillard, C.J.; Huang, H.; Vogt, C.D.; Rodrigues, B.E.; Neumann, T.S.; Sem, D.S.; Schroeder, F.; Cunningham, C.W. Endocannabinoid transport proteins: Discovery of tools to study sterol carrier protein-2. Methods Enzymol. 2017, 593, 99–121. [Google Scholar]
- Plau, J.; Golczak, M.; Paik, J.; Calderon, R.M.; Blaner, W.S. Retinol-binding protein 2 (RBP2): More than just dietary retinoid uptake. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159179. [Google Scholar] [CrossRef]
- Khasabova, I.A.; Golovko, M.Y.; Golovko, S.A.; Simone, D.A.; Khasabov, S.G. Intrathecal administration of resolvin D1 and E1 decreases hyperalgesia in mice with bone cancer pain: Involvement of endocannabinoid signaling. Prostaglandins Other Lipid Mediat. 2020, 151, 106479. [Google Scholar] [CrossRef] [PubMed]
- Hermes, D.J.; Yadav-Samudrala, B.J.; Xu, C.; Paniccia, J.E.; Meeker, R.B.; Armstrong, M.L.; Reisdorph, N.; Cravatt, B.F.; Mackie, K.; Lichtman, A.H.; et al. GPR18 drives FAAH inhibition-induced neuroprotection against HIV-1 Tat-induced neurodegeneration. Exp. Neurol. 2021, 341, 113699. [Google Scholar] [CrossRef]
- McHugh, D. GPR18 in microglia: Implications for the CNS and endocannabinoid system signalling. Br. J. Pharmacol. 2012, 167, 1575–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccarrone, M.; Fiorucci, L.; Erba, F.; Bari, M.; Finazzi-Agrò, M.; Ascoli, F. Human mast cells take up and hydrolyze anandamide under the control of 5-lipoxygenase and do not express cannabinoid receptors. FEBS Lett. 2000, 468, 176–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti-Solano, M. A multi-dimensional view of context-dependent G protein-coupled receptor function. Biochem. Soc. Trans. 2023; in press. [Google Scholar] [CrossRef]
- Chen, Q.; Tesmer, J.J.G. G protein-coupled receptor interactions with arrestins and GPCR kinases: The unresolved issue of signal bias. J. Biol. Chem. 2022, 298, 102279. [Google Scholar] [CrossRef]
- Grabiec, U.; Hohmann, T.; Ghadban, C.; Rothgänger, C.; Wong, D.; Antonietti, A.; Growth, T.; Mackie, K.; Dehghani, F. Protective effect of N-arachidonoyl glycine-GPR18 signaling after excitotoxical lesion in murine organotypic hippocampal slice cultures. Int. J. Mol. Sci. 2019, 20, 1266. [Google Scholar] [CrossRef] [Green Version]
- Patricio, F.; Morales-Andrade, A.A.; Patricio-Martínez, A.; Limón, I.D. Cannabidiol as a therapeutic target: Evidence of its neuroprotective and neuromodulatory function in Parkinson’s disease. Front. Pharmacol. 2020, 11, 595635. [Google Scholar] [CrossRef]
- Rinne, P.; Guillamat-Prats, R.; Rami, M.; Bindila, L.; Ring, L.; Lyytikäinen, L.P.; Raitoharju, E.; Oksala, N.; Lehtimäki, T.; Weber, C.; et al. Palmitoylethanolamide promotes a proresolving macrophage phenotype and attenuates atherosclerotic plaque formation. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2562–2575. [Google Scholar] [CrossRef]
- Jiang, L.; Chen, Y.; Huang, X.; Yuan, A.; Shao, Q.; Pu, J.; He, B. Selective activation of CB2 receptor improves efferocytosis in cultured macrophages. Life Sci. 2016, 161, 10–18. [Google Scholar] [CrossRef]
- Reyes-Resina, I.; Navarro, G.; Aguinaga, D.; Canela, E.I.; Schoeder, C.T.; Załuski, M.; Kieć-Kononowicz, K.; Saura, C.A.; Müller, C.E.; Franco, R. Molecular and functional interaction between GPR18 and cannabinoid CB2 G-protein-coupled receptors. Relevance in neurodegenerative diseases. Biochem. Pharmacol. 2018, 157, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Langmead, C.J.; Riddy, D.M. New advances in targeting the resolution of inflammation: Implications for specialized pro-resolving mediator GPCR drug discovery. ACS Pharmacol. Transl. Sci. 2020, 3, 88–106. [Google Scholar] [CrossRef]
- Pamplona, F.A.; Ferreira, J.; Menezes de Lima, O., Jr.; Duarte, F.S.; Bento, A.F.; Forner, S.; Villarinho, J.G.; Bellocchio, L.; Wotjak, C.T.; Lerner, R.; et al. Anti-inflammatory lipoxin A4 is an endogenous allosteric enhancer of CB1 cannabinoid receptor. Proc. Natl. Acad. Sci. USA 2012, 109, 21134–21139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G. Lipoxin A4 is an allosteric endocannabinoid that strengthens anandamide-induced CB1 receptor activation. Proc. Natl. Acad. Sci. USA 2012, 109, 20781–206782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M.; Bucher, C.; Liu, W.; Müller, M.; Schmidt, T.; Kardell, M.; Driessen, M.N.; Rossaint, J.; Gross, E.R.; Wagner, N.M. 12(S)-HETE mediates diabetes-induced endothelial dysfunction by activating intracellular endothelial cell TRPV1. J. Clin. Investig. 2020, 130, 4999–5010. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xiong, M.; Zong, X.; Ge, Y.; Hui Zhang, H.; Wang, M.; Han, G.W.; Yi, C.; Ma, L.; Ye, R.D.; et al. Structural basis of ligand binding modes at the human formyl peptide receptor 2. Nat. Commun. 2020, 11, 1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; He, X.; Zhao, J.; Jiang, H.; Cheng, X.; Xia, Y.; Xu, H.E.; He, Y. Structural basis of leukotriene B4 receptor 1 activation. Nat. Commun. 2022, 13, 1156. [Google Scholar] [CrossRef]
- Nakamura, H.; Murayama, T. Role of sphingolipids in arachidonic acid metabolism. J. Pharmacol. Sci. 2014, 124, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Standoli, S.; Pecchioli, S.; Tortolani, D.; Di Meo, C.; Fanti, F.; Sergi, M.; Bacci, M.; Seidita, I.; Bernacchioni, C.; Donati, C.; et al. The TRPV1 receptor is up-regulated by sphingosine 1-phosphate and is implicated in the anandamide-dependent regulation of mitochondrial activity in C2C12 myoblasts. Int. J. Mol. Sci. 2022, 23, 11103. [Google Scholar] [CrossRef]
- Kang, J.W.; Choi, H.S.; Shin, J.K.; Lee, S.M. Resolvin D1 activates the sphingosine-1-phosphate signaling pathway in murine livers with ischemia/reperfusion injury. Biochem. Biophys. Res. Commun. 2019, 514, 1058–1065. [Google Scholar] [CrossRef]
- Grimaldi, P.; Pucci, M.; Di Siena, S.; Di Giacomo, D.; Pirazzi, V.; Geremia, R.; Maccarrone, M. The faah gene is the first direct target of estrogen in the testis: Role of histone demethylase LSD1. Cell. Mol. Life Sci. 2012, 69, 4177–4190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccarrone, M.; Bari, M.; Di Rienzo, M.; Finazzi-Agrò, A.; Rossi, A. Progesterone activates fatty acid amide hydrolase (FAAH) promoter in human T lymphocytes through the transcription factor Ikaros. Evidence for a synergistic effect of leptin. J. Biol. Chem. 2003, 278, 32726–32732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Muñoz, L.J.; Ulloa, E.V.; Sahlgren, C.; Lizano, M.; De La Cruz-Hernández, E.; Contreras-Paredes, A. Modulating epigenetic modifications for cancer therapy. Oncol. Rep. 2023, 49, 59. [Google Scholar] [CrossRef] [PubMed]
- Punt, J.M.; van der Vliet, D.; van der Stelt, M. Chemical probes to control and visualize lipid metabolism in the brain. Acc. Chem. Res. 2022, 55, 3205–3217. [Google Scholar] [CrossRef]
- Gazzi, T.; Brennecke, B.; Atz, K.; Korn, C.; Sykes, D.; Forn-Cuni, G.; Pfaff, P.; Sarott, R.C.; Westphal, M.V.; Mostinski, Y.; et al. Detection of cannabinoid receptor type 2 in native cells and zebrafish with a highly potent, cell-permeable fluorescent probe. Chem. Sci. 2022, 13, 5539–5545. [Google Scholar] [CrossRef]
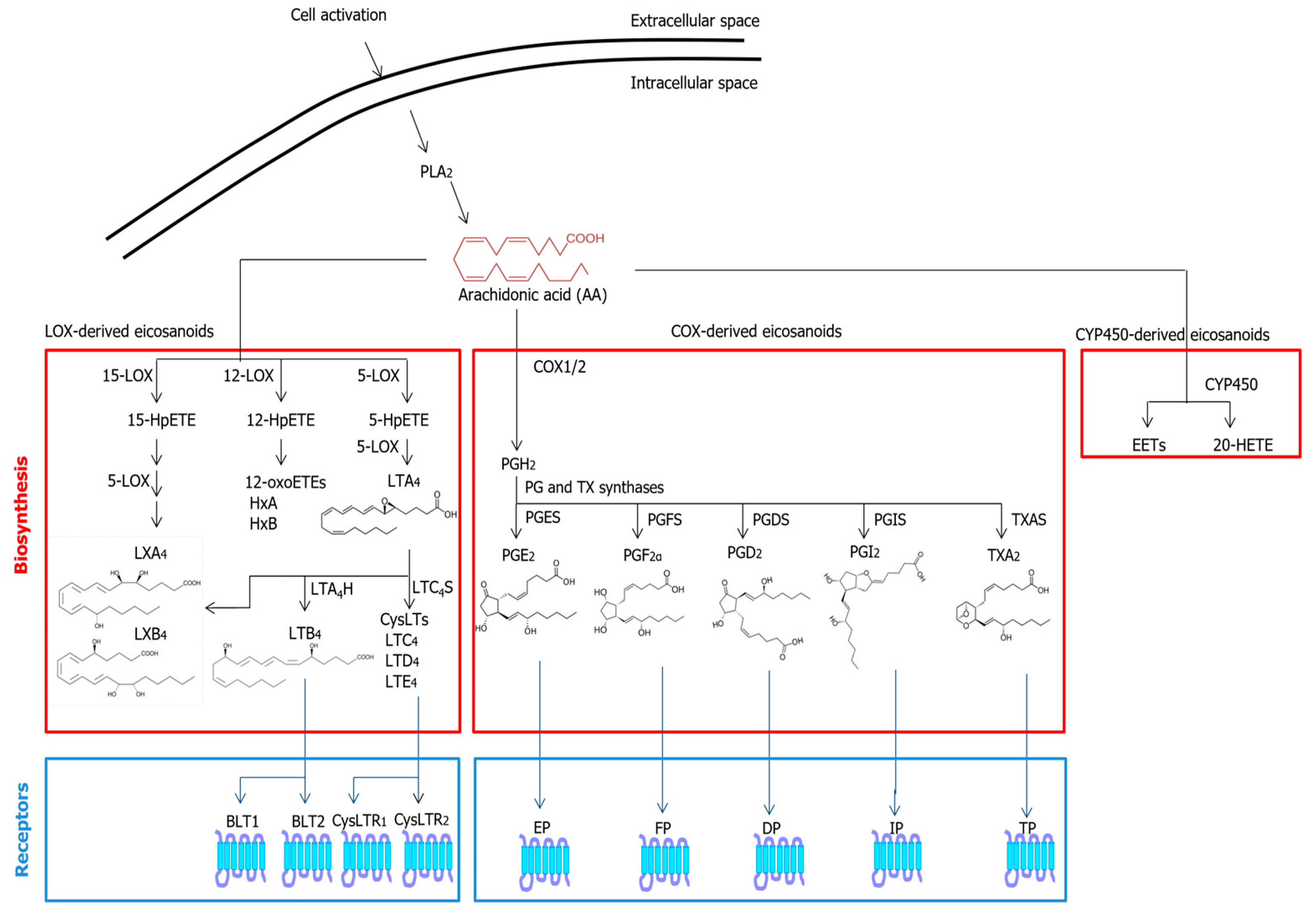
Figure 1.
Metabolic pathways and known receptors of classical EICs. Abbreviations: 20-HETE—20-hydroxyeicosatetraenoic acid; BLT—leukotriene B4 receptor; COX—cyclooxygenase; CYP450—cytochrome P450; DP—prostaglandin D receptor; EET—epoxyeicosatrienoic acid; EP—prostaglandin E receptor; FP—prostaglandin F receptor; HpETE—hydroperoxyeicosatetraenoic acid; IP—prostacyclin I receptor; LOX—lipoxygenase; LT—leukotriene; LX—lipoxin; PG—prostaglandin; PGDS—prostaglandin D2 synthase; PGES—prostaglandin E synthase; PGFS—prostaglandin F synthase; PGI2—prostacyclin 2; PGIS—prostacyclin I synthase; PLA2—phospholipase A2; TP—thromboxane receptor; TXAS—thromboxane synthase.
Figure 1.
Metabolic pathways and known receptors of classical EICs. Abbreviations: 20-HETE—20-hydroxyeicosatetraenoic acid; BLT—leukotriene B4 receptor; COX—cyclooxygenase; CYP450—cytochrome P450; DP—prostaglandin D receptor; EET—epoxyeicosatrienoic acid; EP—prostaglandin E receptor; FP—prostaglandin F receptor; HpETE—hydroperoxyeicosatetraenoic acid; IP—prostacyclin I receptor; LOX—lipoxygenase; LT—leukotriene; LX—lipoxin; PG—prostaglandin; PGDS—prostaglandin D2 synthase; PGES—prostaglandin E synthase; PGFS—prostaglandin F synthase; PGI2—prostacyclin 2; PGIS—prostacyclin I synthase; PLA2—phospholipase A2; TP—thromboxane receptor; TXAS—thromboxane synthase.

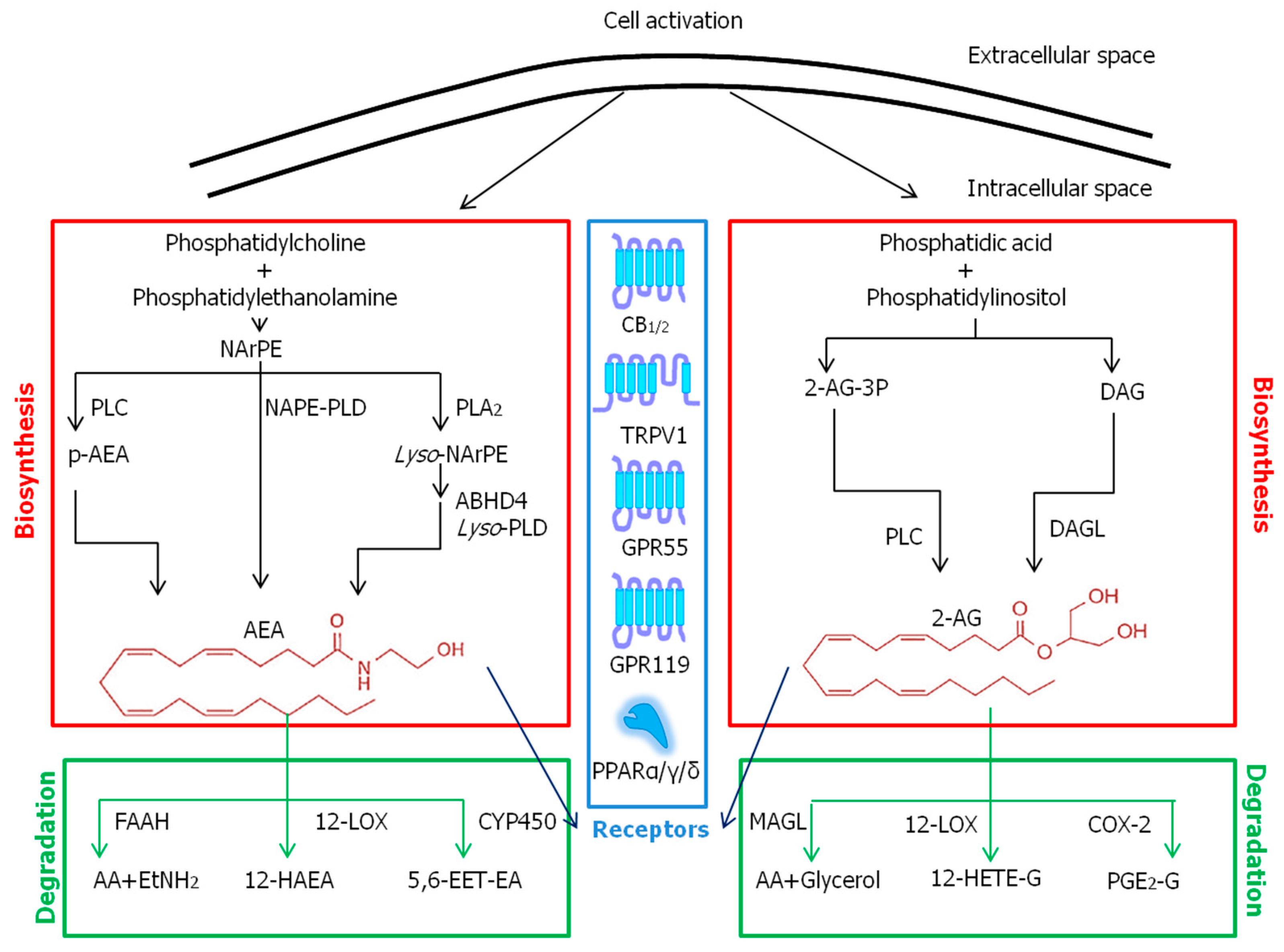
Figure 2.
Metabolic pathways and known receptors of ECBs. Abbreviations: 12-HAEA—12-hydroxy-N-arachidoylethanolamine; 12-HETE-G—12-hydroxy eicosatetraenoylglycerol; 2-AG—2-arachidonoylglycerol; 5,6-EET-EA—5,6-epoxyeicosatrienoyl ethanolamide; AA—arachidonic acid; ABHD4—αβ-hydrolase domain 4 protein; AEA—N-arachidonoylethanolamine; CB1/2—cannabinoid receptor 1/2; COX-2—cyclooxygenase-2; CYP450—cytochrome P450; DAG—diacylglycerol; DAGL—diacylglycerol lipase; EtNH2—ethanolamine; FAAH—fatty acid amide hydrolase; GPR—G protein-coupled receptor; LOX—lipoxygenase; Lyso-NArPE—lyso-N-arachidonoyl phosphatidylethanolamine; Lyso-PLD—lysophospholipase D; MAGL—monoacylglycerol lipase; NAPE-PLD—N-acylphosphatidylethanolamines-specific phospholipase D; NArPE—N-arachidonoyl phosphatidylethanolamine; p-AEA—phospho-N-arachidonoylethanolamine; PGE2-G—prostaglandin E2 glycerol; PLA2—phospholipase A2; PLC—phospholipase C; PPARα/γ/δ—peroxisome proliferator activated receptor α/γ/δ; TRPV1—transient receptor potential vanilloid type-1.
Figure 2.
Metabolic pathways and known receptors of ECBs. Abbreviations: 12-HAEA—12-hydroxy-N-arachidoylethanolamine; 12-HETE-G—12-hydroxy eicosatetraenoylglycerol; 2-AG—2-arachidonoylglycerol; 5,6-EET-EA—5,6-epoxyeicosatrienoyl ethanolamide; AA—arachidonic acid; ABHD4—αβ-hydrolase domain 4 protein; AEA—N-arachidonoylethanolamine; CB1/2—cannabinoid receptor 1/2; COX-2—cyclooxygenase-2; CYP450—cytochrome P450; DAG—diacylglycerol; DAGL—diacylglycerol lipase; EtNH2—ethanolamine; FAAH—fatty acid amide hydrolase; GPR—G protein-coupled receptor; LOX—lipoxygenase; Lyso-NArPE—lyso-N-arachidonoyl phosphatidylethanolamine; Lyso-PLD—lysophospholipase D; MAGL—monoacylglycerol lipase; NAPE-PLD—N-acylphosphatidylethanolamines-specific phospholipase D; NArPE—N-arachidonoyl phosphatidylethanolamine; p-AEA—phospho-N-arachidonoylethanolamine; PGE2-G—prostaglandin E2 glycerol; PLA2—phospholipase A2; PLC—phospholipase C; PPARα/γ/δ—peroxisome proliferator activated receptor α/γ/δ; TRPV1—transient receptor potential vanilloid type-1.

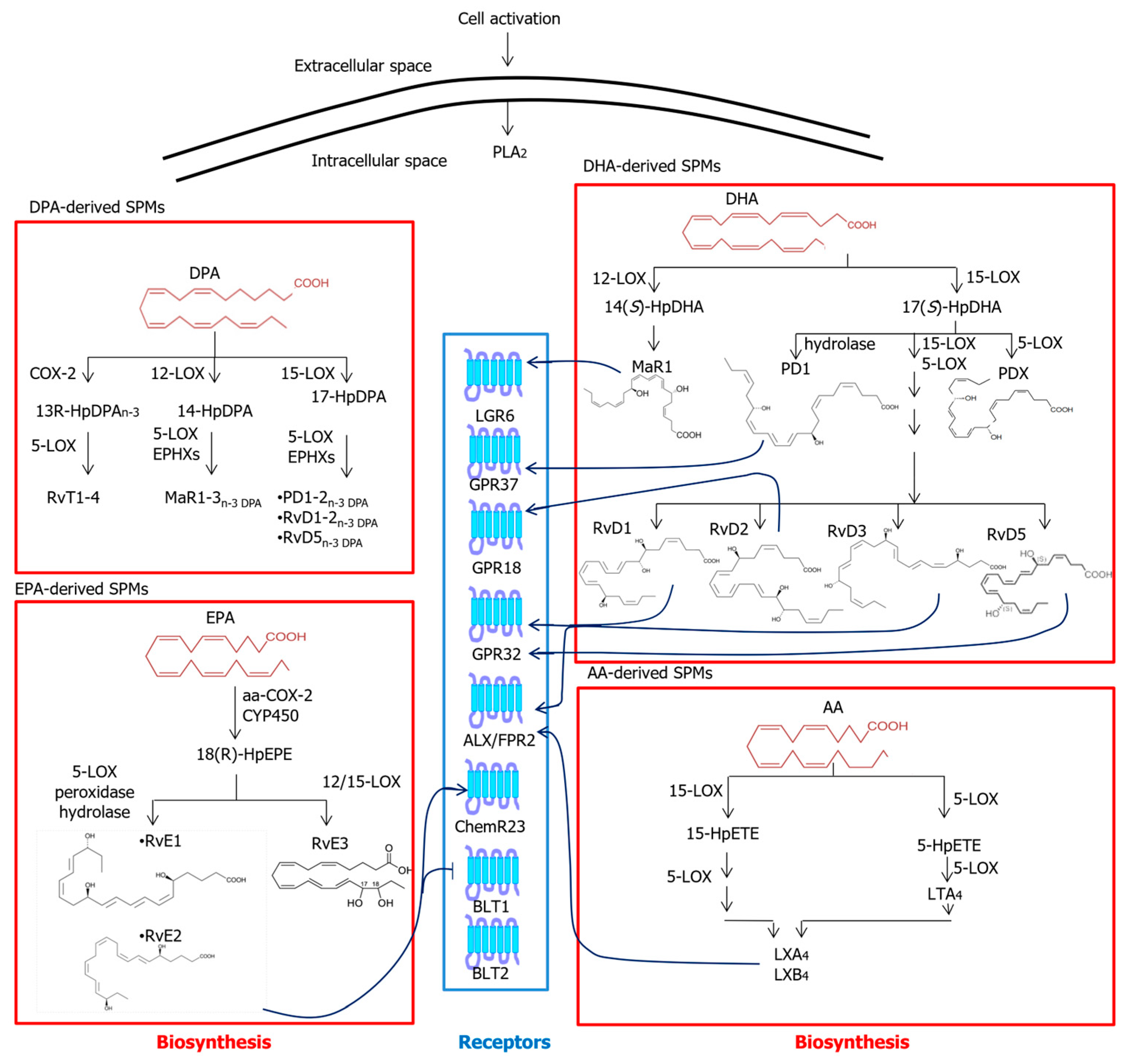
Figure 3.
Metabolic pathways and known receptors of SPMs. Abbreviations: AA—arachidonic acid; ALX/FPR2—formyl peptide receptor 2; BLT—leukotriene B4 receptor; ChemR23—chemerin receptor 23; COX—cyclooxygenase; CYP450—cytochrome P450; DHA—docosahexaenoic acid; EPA—eicosapentaenoic acid; EPHX—epoxide hydrolase; GPR—G protein-coupled receptor; HpDHA—hydroperoxy-docosahexaenoic acid; HpDPA—hydroperoxydocosapentaenoic acid; HpEPE—hydroperoxyeicosapentaenoic acid; HpETE—hydroperoxyeicosatetraenoic acid; LGR6—leucine-rich repeat-containing G protein-coupled receptor 6; LOX—lipoxygenase; LT—leukotriene; LX—lipoxin; MaR1—maresin 1; n-3 DPA—n-3 docosapentaenoic acid; PD1—protectin D1; PDX—protectin DX; PLA2—phospholipase A2; RvD—D-series resolvins; RvE—E-series resolvins; RvTs—13-series resolvins.
Figure 3.
Metabolic pathways and known receptors of SPMs. Abbreviations: AA—arachidonic acid; ALX/FPR2—formyl peptide receptor 2; BLT—leukotriene B4 receptor; ChemR23—chemerin receptor 23; COX—cyclooxygenase; CYP450—cytochrome P450; DHA—docosahexaenoic acid; EPA—eicosapentaenoic acid; EPHX—epoxide hydrolase; GPR—G protein-coupled receptor; HpDHA—hydroperoxy-docosahexaenoic acid; HpDPA—hydroperoxydocosapentaenoic acid; HpEPE—hydroperoxyeicosapentaenoic acid; HpETE—hydroperoxyeicosatetraenoic acid; LGR6—leucine-rich repeat-containing G protein-coupled receptor 6; LOX—lipoxygenase; LT—leukotriene; LX—lipoxin; MaR1—maresin 1; n-3 DPA—n-3 docosapentaenoic acid; PD1—protectin D1; PDX—protectin DX; PLA2—phospholipase A2; RvD—D-series resolvins; RvE—E-series resolvins; RvTs—13-series resolvins.

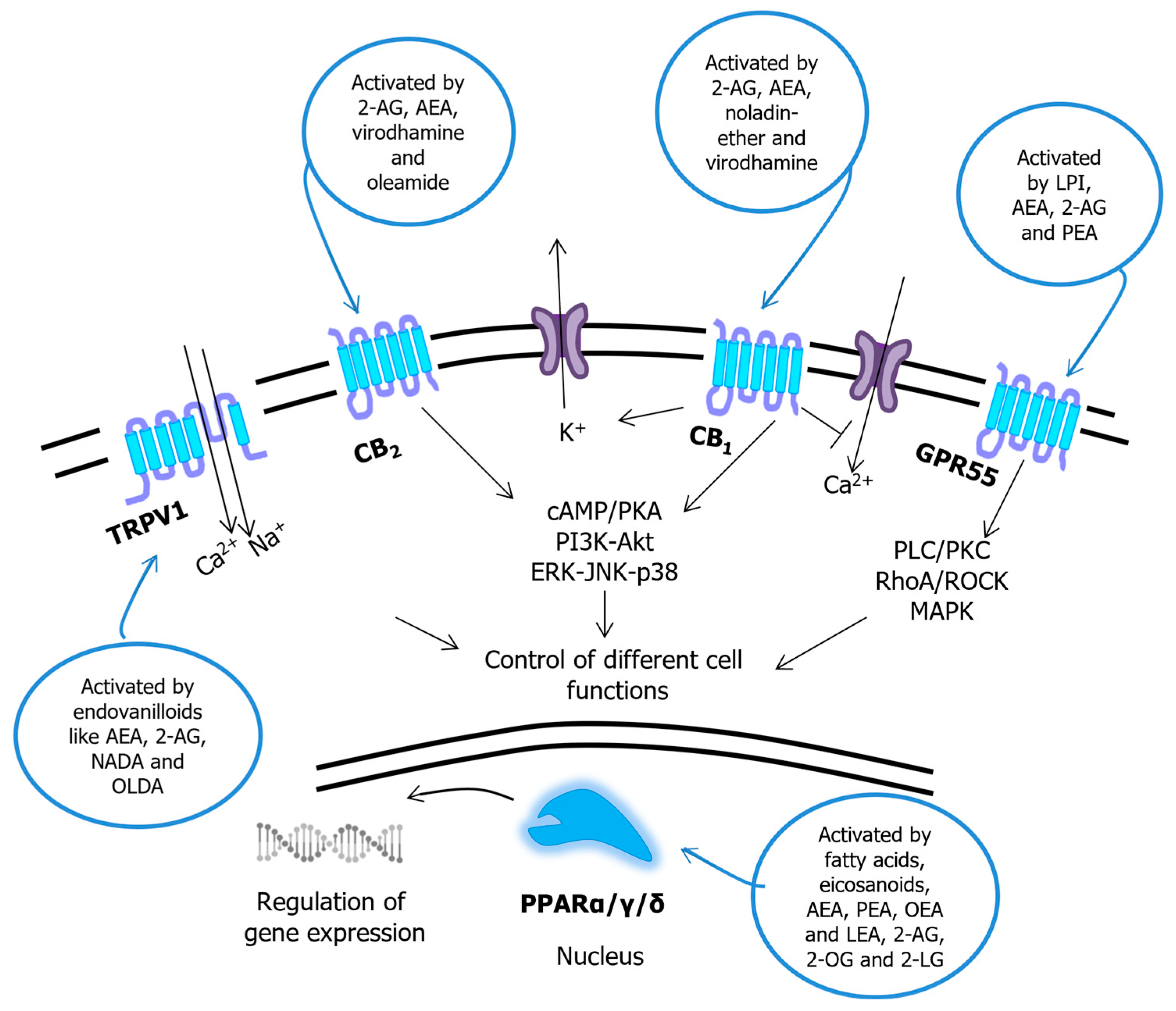
Figure 4.
Signal transduction pathways triggered by endocannabinoids and congeners. Abbreviations: AEA—N-arachidonoylethanolamine (anandamide); 2-AG—2-arachidonoylglycerol; 2-LG—2-linoleoylglycerol; PEA—N-palmitoylethanolamine; LEA—N-linoleoylethanolamine; OEA—N-oleoylethanolamine; 2-OG—2-oleoyglycerol; LPI—lysophosphatidic acid; NADA—N-arachidonoyldopamine; OLDA—N-oleoyldopamine; PLC—phospholipase C; PKC—protein kinase C; ROCK—Rho-associated protein kinase; MAPK—mitogen-activated protein kinase; cAMP—cyclic AMP; PKA—protein kinase A; PI3K—phosphatidylinositol (phosphoinositide) 3-kinase; Akt—α-serine/threonine-protein kinase; ERK—extracellular signal-regulated kinase; JNK—c-Jun N-terminal kinase; GPR55—G protein-coupled receptor 55; CB1/2—cannabinoid receptor 1/2; TRPV1—transient receptor potential vanilloid 1. Modified from ref. [25].
Figure 4.
Signal transduction pathways triggered by endocannabinoids and congeners. Abbreviations: AEA—N-arachidonoylethanolamine (anandamide); 2-AG—2-arachidonoylglycerol; 2-LG—2-linoleoylglycerol; PEA—N-palmitoylethanolamine; LEA—N-linoleoylethanolamine; OEA—N-oleoylethanolamine; 2-OG—2-oleoyglycerol; LPI—lysophosphatidic acid; NADA—N-arachidonoyldopamine; OLDA—N-oleoyldopamine; PLC—phospholipase C; PKC—protein kinase C; ROCK—Rho-associated protein kinase; MAPK—mitogen-activated protein kinase; cAMP—cyclic AMP; PKA—protein kinase A; PI3K—phosphatidylinositol (phosphoinositide) 3-kinase; Akt—α-serine/threonine-protein kinase; ERK—extracellular signal-regulated kinase; JNK—c-Jun N-terminal kinase; GPR55—G protein-coupled receptor 55; CB1/2—cannabinoid receptor 1/2; TRPV1—transient receptor potential vanilloid 1. Modified from ref. [25].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of entries of a PubMed search with “eicosanoids”, “endocannabinoids” and “specialized pro-resolving mediators” over the last 30 years.
Table 1.
Number of entries of a PubMed search with “eicosanoids”, “endocannabinoids” and “specialized pro-resolving mediators” over the last 30 years.
| Time Range | EICs | ECBs | SPMs |
|---|---|---|---|
| 1992–1997 | 22,258 | 200 | 3 |
| 1998–2002 | 16,534 | 648 | 5 |
| 2003–2007 | 17,065 | 1791 | 18 |
| 2008–2012 | 16,128 | 2866 | 85 |
| 2013–2017 | 14,402 | 3271 | 316 |
| 2018–2022 | 10,713 | 3949 | 847 |
Abbreviations: EICs, eicosanoids; ECBs, endocannabinoids; SPMs, specialized pro-resolving mediators.
Table 2.
Common metabolic enzymes and receptors of different classes of bioactive lipids.
| Enzyme/Receptor | EICs | ECBs | SPMs |
|---|---|---|---|
| PLA2 | + | + | + |
| CYP450 | + | + | + |
| 12-LOX | + | + | + |
| COX-2 | + | + | + |
| CB1 | + | + | + |
| CB2 | + | + | |
| GPR18 | + | + | |
| TRPV1 | + | + | + |
| BLT1 | + | + |
Abbreviations: EICs—eicosanoids; ECBs—endocannabinoids; SPMs—specialized pro-resolving mediators; PLA2—phospholipase A2; CYP450—cytochrome P450; 12-LOX—12-lipoxygenase; COX-2: cyclooxygenase-2; CB1/2—cannabinoid receptor 1/2; GPR18—G protein-coupled receptor 18; TRPV1—transient receptor potential vanilloid-1; BLT1—leukotriene B4 receptor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maccarrone, M. Deciphering Complex Interactions in Bioactive Lipid Signaling. Molecules 2023, 28, 2622. https://doi.org/10.3390/molecules28062622
AMA Style
Maccarrone M. Deciphering Complex Interactions in Bioactive Lipid Signaling. Molecules. 2023; 28(6):2622. https://doi.org/10.3390/molecules28062622
Chicago/Turabian StyleMaccarrone, Mauro. 2023. "Deciphering Complex Interactions in Bioactive Lipid Signaling" Molecules 28, no. 6: 2622. https://doi.org/10.3390/molecules28062622