
A Review Concerning the Polysaccharides Found in Edible and Medicinal Plants in Xinjiang
Abstract
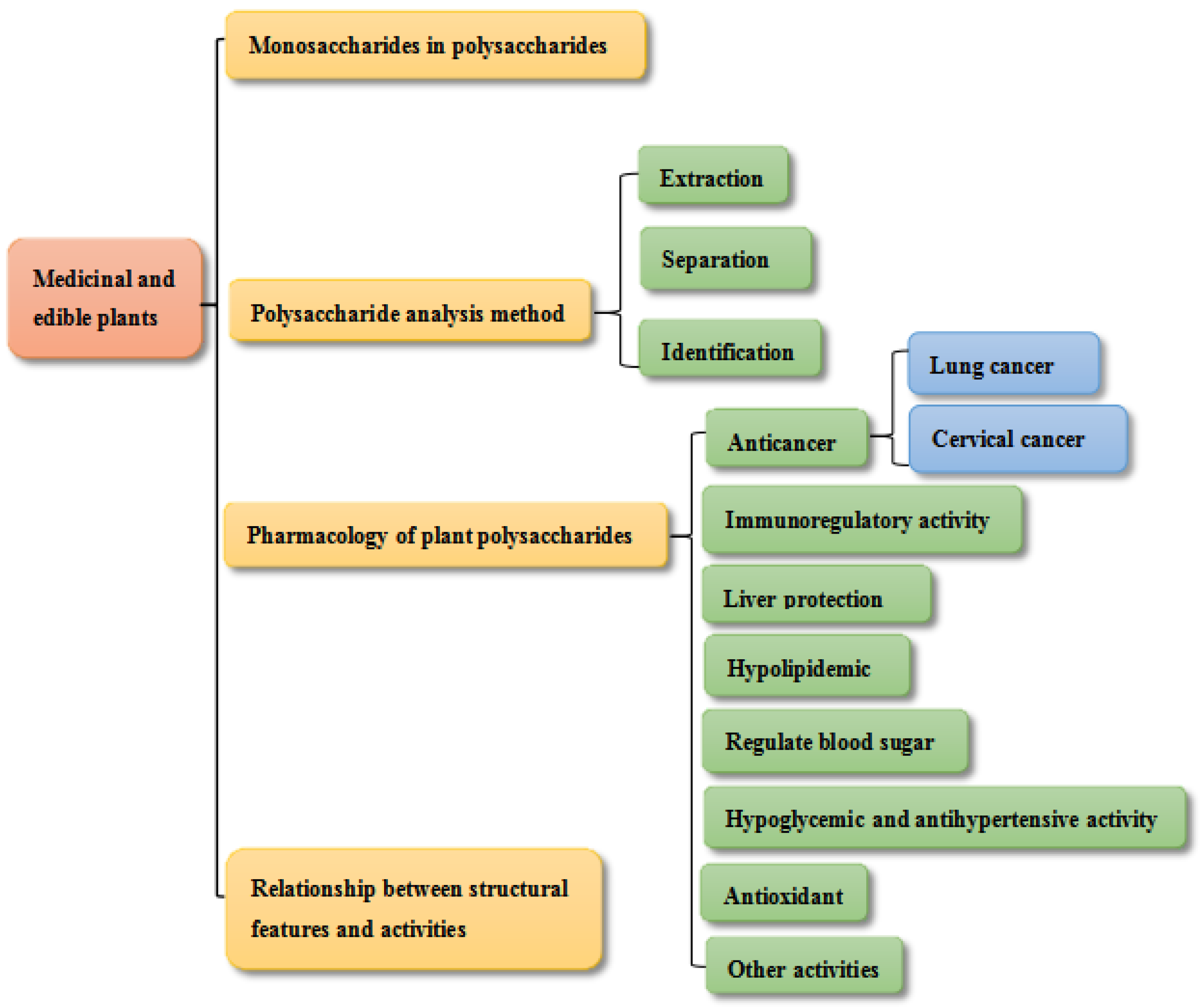
:1. Introduction
2. Monosaccharides in Polysaccharides
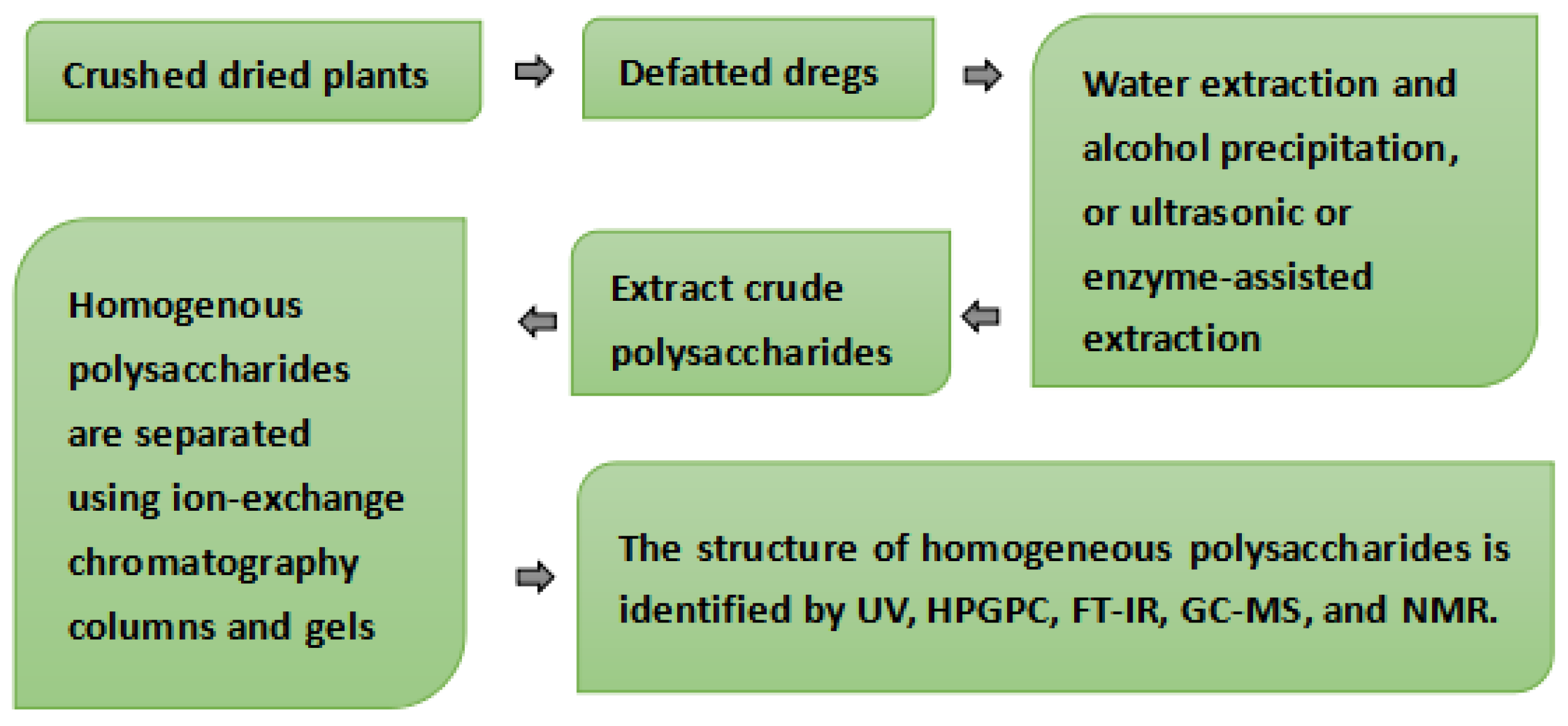
3. Polysaccharide Analysis Method
4. Pharmacological Activity
4.1. Anticancer
4.1.1. Lung Cancer
4.1.2. Cervical Cancer
4.1.3. Other Cancers
4.2. Immunoregulatory Activity
4.3. Liver Protection
4.4. Hypolipidemic Activity
4.5. Hypoglycemic and Antihypertensive Activity
4.6. Antioxidant
4.7. Other Activities
5. Relationship between Structural Features and Activities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Molecular Weight of Polysaccharides | Polysaccharides | Disease and Function | Experimental Subject | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Turnip | 7590~6080 Da and 4751~6873 Da | 8.99% | Inflammation | RAW 264.7 macrophage | Regulate NF-κB, TNF-α proteins | [81] |
| Regulate blood sugar level | SD rat | Promote hepatic glycogen synthesis and suppress its decomposition | [82] | |||
| Asthma | Wistar rat | Control TNF-α, IL-6, IL-2, and CRP level | [75] | |||
| Lung cancer | Lewis lung cancer mice | - | [56] | |||
| Kunlun snow chrysanthemum | 8200~8700 Da and 6100~6500 Da. | 8.96% | Acute liver injury | KM mouse | Affects tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) level | [83,84] |
| Anti-colon cancer | HCT116 cells, CACO-2 cells | Apoptosis | [58,85] | |||
| Pomegranate | 14~1400 kDa | Skin: 27.3% Leaf: 1.809% Seed: 2.65% | Lipid lowering | - | Inhibition of cholesterol esterase and pancreatic lipase | [9,18] |
| Immunoregulation | RAW 264.7 macrophage | Inhibition of NO release | [86] | |||
| Regulate blood sugar and blood lipids | SD rat | Lower blood glucose and serum TC, TG, and LDL-C, and increase serum HDL-C | [87] | |||
| Mulberry | 80~250 kDa | 9.95% | Myocardial inflammatory response | H9C2 cells | Apoptosis | [88,89,90,91,92,93] |
| Cancer | MCF-7 cells, Kupffer cells, HeLa cell, MKN-45 cells, NCI-H1650 cells | Affects the content of MDA, NO, IL-6, IL-1 β, and NF-κB, and the viability values of TNF-α | [24,60] | |||
| Alcoholic liver injury | SPF mouse | Intervention of linoleic acid, α-linoleic acid, and glycerophospholipid metabolism | [68] | |||
| Jujube | 45,900, 69,860, and 195,100 Da | 3.68% | Diabetes | Mouse | Lower the FBG, TC, and TG in diabetic mice, and increase the HDL content | [53] |
| Lipid lowering | Rat | Reduce TG, CHO, LDL-C, and increase the levels of HDL-C. Improve T-AOC, SOD level, and reduce the MDA content | [54,55] | |||
| Cancer | MKN-45 | Apoptosis | [55,56] |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, X.L.; Hou, C.Y.; Shi, M.M.; Yan, Y.Z.; Liu, Y.Q. An insight into the research concerning Panax ginseng C. A. Meyer polysaccharides: A review. Food Rev. Int. 2020, 38, 1771363. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, X.R.; Guo, Q.S. Comparative analysis of main chemical constituents of Prunella wilwilt from nine habitats. J. Chengdu Med. Coll. 2017, 12, 551–566. [Google Scholar] [CrossRef]
- Li, Y.; Han, J.; Chen, Y. p-Coumaric acid as a prophylactic measure against normobaric hypoxia induced pulmonary edema in mice. Life Sci. 2018, 211, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Lin, C.Y.; Bian, Z.X. An insight into anti-inflammatory effects of fungal beta-glucans. Trends Food Sci. Technol. 2015, 41, 49–59. [Google Scholar] [CrossRef]
- Falco, A.; Frost, P.; Miest, J. Reduced inflammatory response to Aeromonas salmonicida infection in common carp (Cyprinus carpio L.) fed with β-glucan supplements. Fish Shellfish Immunol. 2012, 32, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.L.; Li, Y.X.; Cui, Y.S. Isolation, purification and structural characterization of two pectin-type polysaccharides from Coreopsis tinctoria Nutt. and their proliferation activities on RAW264.7 cells. Glycoconj. J. 2021, 38, 251–259. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Ye, H.; Hu, B.; Zhou, L.; Jabbar, S.; Zeng, X.; Shen, W. Optimization of extraction, characterization and antioxidant activity of polysaccharides from Brassica rapa L. Int. J. Biol. Macromol. 2016, 82, 979–988. [Google Scholar] [CrossRef]
- Yang, Y.M. Study on Extraction and Purification, Structure Characterization and Antioxidant Activity of Polysaccharides from Hami Jujube. Master’s Thesis, Shandong Agriculture University, Shandong, China, 2021. [Google Scholar]
- Ji, X.L.; Guo, J.H.; Ding, D.; Gao, J.; Hao, L.; Guo, X.; Liu, Y. Structural characterization and antioxidant activity of a novel high-molecular-weight polysaccharide from Ziziphus Jujuba cv. Muzao. J. Food Meas. Charact. 2022, 16, 2191–2200. [Google Scholar] [CrossRef]
- Ren, A.N.; Lu, Y.; Zou, Y.F. Isolation and purification of polysaccharides from Carthamus sativa L. and analysis of monosaccharide composition. Chin. J. Pharm. Anal. 2013, 237, 1190–1196. [Google Scholar] [CrossRef]
- Hu, Y. Structural Analysis and Antioxidant Activities of Polysaccharide Isolated from Carthamus tinctorius L. Master’s Thesis, Shihezi University, Shihezi, China, 2020. [Google Scholar]
- Yao, Y.; Yao, J.; Du, Z.; Wang, P.; Ding, K. Structural Elucidation and Immune-Enhancing Activity of an Arabinogalactan from Flowers of Carthamus tinctorius L. Carbohydr. Polym. 2018, 202, 134–142. [Google Scholar] [CrossRef]
- Wang, W.; Buligen, J.; Chen, K. The xinjiang turnip pectic polysaccharide composition analysis and the evaluation of the immune activity. Food Ind. Sci. Technol. 2019, 40, 62–67. [Google Scholar] [CrossRef]
- Gong, G.; Fan, J.; Sun, Y.; Wu, Y.; Liu, Y.; Sun, W.; Zhang, Y.; Wang, Z. Isolation, structural characterization, and antioxidativity of polysaccharide LBLP5-a from lycium barbarum leaves. Process Biochem. 2016, 51, 314–324. [Google Scholar] [CrossRef]
- Redgwell, R.J.; Curti, D.; Wang, J.; Dobruchowska, J.M.; Gerwig, G.J.; Kamerling, J.P.; Bucheli, P. Cell wall polysaccharides of Chinese Wolfberry (Lycium barbarum): Part 2. Characterisation of arabinogalactan-proteins. Carbohydr. Polym. 2011, 84, 1075–1083. [Google Scholar] [CrossRef]
- Eradan, M.; Li, Y.; Yao, J.; Yuan, H. Isolation and immunoactivity of polysaccharides from Cistanola tubulosa. Chin. J. Pharm. 2021, 32, 1479–1484. [Google Scholar]
- Wang, R.Q.; Yang, X.G.; Xu, W.T. Research progress of sea-buckthorn polysaccharide. Food Res. Dev. 2017, 38, 221–224. [Google Scholar] [CrossRef]
- Shen, C.; Wang, T.; Guo, F.; Sun, K.; Wang, B.; Wang, J.; Zhang, Z.; Zhang, X.; Zhao, Y.; Chen, Y. Structural characterization and intestinal protection activity of polysaccharides from Sea buckthorn (Hippophae rhamnoides L.) berries. Carbohydr. Polym. 2021, 274, 118648. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Zheng, W.H.; Zhang, K.X. The chemical constituents, pharmacological effects of seabuckthorn research progress and its forecast analysis quality markers. Chin. J. Tradit. Chin. Med. 2021, 11, 1001–5302. [Google Scholar] [CrossRef]
- Wufuer, R.; Bai, J.; Liu, Z.; Zhou, K.; Taoerdahong, H. Biological activity of Brassica rapa L. polysaccharides on RAW264.7 macrophages and on tumor cells. Bioorganic Med. Chem. 2020, 28, 115330. [Google Scholar] [CrossRef]
- Chen, W. Isolation, Purification, Structure Identification and Pharmacological Activity of Schinulin Polymers. Master’s Thesis, Guangdong Pharmaceutical University, Guangdong, China, 2018. [Google Scholar]
- Yang, B. Honey Tree Purification and Structure Analysis of Polysaccharides and Its Fall Blood Sugar Active Research. Ph.D. Thesis, Southwest University, Chongqing, China, 2020. [Google Scholar]
- Li, R. Study on the Immunomodulatory Effect and Antitumor Activity of Mulberry Polysaccharides and Their Derivatives. Master’s Thesis, Guizhou Normal University, Guizhou, China, 2020. [Google Scholar]
- Wang, M.; Liu, Z.H.; Zhang, Y. Analysis of flavonoids, polyphenols and their antioxidant activities in two seabuckthorn species from Xinjiang. Sci. Technol. Food Ind. 2020, 41, 51–57. [Google Scholar] [CrossRef]
- Chen, X.X. Hops Polysaccharide Extraction, Isolation, Purification and Structure Identification. Master’s Thesis, Guangdong Pharmaceutical University, Guangdong, China, 2020. [Google Scholar]
- Zhang, P.L.; Chen, X.F.; Wang, Q. Isolation and anti-aging activity of Polysaccharide T-3 from Mulberry and Caenorhabditis elegans. J. Tianjin Univ. Tradit. Chin. Med. 2017, 36, 136–141. [Google Scholar] [CrossRef]
- Adilijiang, S.; Ainivar, W.; Mardan, Y. Network pharmacology study on the active components of volatile Oil from Basil in Xinjiang. Chin. Pharm. Bull. 2019, 35, 701–706. [Google Scholar] [CrossRef]
- Akoto, C.O.; Acheampong, A.; Boakye, Y.D. Anti-inflammatory, antioxidant, and anthelmintic activities of ocimum basilicum (sweet basil) fruits. J. Chem. 2020, 10, 1155. [Google Scholar] [CrossRef]
- Wang, S.P. Alfalfa Polysaccharide Extraction, Purification and Its Impact on Laying Hens Liver Cell Biological Activity. Doctor Dissertation, Chinese Academy of Agricultural Sciences, Beijing, China, 2014. [Google Scholar]
- Chen, L.; Liu, J.; Zhang, Y. Structural, thermal and anti-inflammatory properties of a novel pectic polysaccharide from alfalfa (Medicago sativa L.) stem. J. Agric. Food Chem. 2015, 63, 3219–3228. [Google Scholar] [CrossRef] [PubMed]
- Israeli, A.; Grinblat, G.; Shochat, I.; Sarid, M.; Dudkiewicz, M.; Braverman, I. Clinical Efficacy of Topical Nasal Pomegranate Fruit Extract for Chronic Rhinitis and Chronic Rhinosinusitis. Ear Nose Throat J. 2021, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.L.; Zhang, M. Preparation of polysaccharides from pomegranate peel and comparison of their biological activities. Lihizhen Med. Mater. Med. Res. 2019, 30, 2377–2380. [Google Scholar] [CrossRef]
- Chen, S.; Xu, C.J.; Li, Y.B. Structural analysis of Polysaccharides from Averagella mushroom. Mod. Food Sci. Technol. 2015, 3, 29–37. [Google Scholar] [CrossRef]
- Taoerdahong, H.; Zhou, K.; Yang, F.; Dong, C.X. Structure, immunostimulatory activity, and the effect of ameliorating airway inflammation of polysaccharides from Pyrus sinkiangensis Yu. Int. J. Biol. Macromol. 2022, 195, 246–254. [Google Scholar] [CrossRef]
- Hairengulii, M.; Ayixiaguli, B.; Hailiqian, T. Determination of polysaccharide content in peel of Korla fragrant pear in Xinjiang. J. Xinjiang Med. Univ. 2018, 9, 1157–1159. [Google Scholar]
- Tsumuraya, Y.; Ozeki, E.; Ooki, Y.; Yoshimi, Y.; Hashizume, K.; Kotake, T. Properties of arabinogalactan-proteins in European pear (Pyrus communis L.) fruits. Carbohydr. Res. 2019, 485, 107816. [Google Scholar] [CrossRef]
- Zhou, K.; Taoerdahong, H.; Bai, J.; Bakasi, A.; Wang, X.J.; Dong, C.X. Structural characterization and immunostimulatory activity of polysaccharides from Pyrus sinkiangensis Yu. Int. J. Biol. Macromol. 2020, 157, 444–451. [Google Scholar] [CrossRef]
- Ayitusun, A.; Parhati, R.; Gao, Y.H. Optimization of extraction technology and structural characterization of Polysaccharides from Fritillaria fritillaria L. Chin. Pharm. 2020, 31, 276–281. [Google Scholar] [CrossRef]
- Guo, H.J. The Dandelion Polysaccharide Extraction, Isolation, Purification, Identification and the Preliminary Research of the Biological Activity. Master’s Thesis, Shihezi University, Shihezi, China, 2019. [Google Scholar]
- Cai, L.; Chen, B.; Yi, F. Optimization of extraction of polysaccharide from dandelion root by response surface methodology: Structural characterization and antioxidant activity. Int. J. Biol. Macromol. 2019, 140, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.G.; Zhu, Q.; Wang, F. Purification and molecular weight determination of lilium. Polysaccharide 2013, 134, 1–4. [Google Scholar] [CrossRef]
- Song, S.; Liu, X.; Zhao, B. Effects of Lactobacillus plantarum Fermentation on the Chemical Structure and Antioxidant Activity of Polysaccharides from Bulbs of Lanzhou Lily. ACS Omega 2021, 6, 29839–29851. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Geng, C.A.; Yang, T.H. Phytochemical and Health-Beneficial Progress of Turnip (Brassica rapa). J. Food Sci. 2019, 84, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Ninomiya, M.; Efdi, M.; Inuzuka, T. Chalcone glycosides from aerial parts of Brassica rapa L. ‘hidabeni’, turnip. Phytochem. Lett. 2010, 3, 96. [Google Scholar] [CrossRef]
- Zhang, H.; Zhong, J.; Zhang, Q. Structural elucidation and bioactivities of a novel arabinogalactan from Coreopsis tinctoria. Carbohydr. Polym. 2019, 219, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.Q.; Ma, F.J.; Pu, Y.F. Study on antioxidant activity of saponins from Turnips in Xinjiang in vivo. Food Res Dev. 2016, 37, 6–10. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, S.Q.; Pan, G.Y. The kunlun snow chrysanthemum research progress on chemical composition and functional activity. Chin. Fruit 2019, 9, 44–48. [Google Scholar] [CrossRef]
- Nair, M.; Saxena, A.; Kaur, C. Characterization and Antifungal Activity of Pomegranate Peel Extract and its Use in Polysaccharide-Based Edible Coatings to Extend the Shelf-Life of Capsicum (Capsicum annuum L.). Food Bioprocess Technol. 2018, 11, 1317–1327. [Google Scholar] [CrossRef]
- Malek Mahdavi, A.; Seyedsadjadi, N.; Javadivala, Z. Potential effects of pomegranate (Punica granatum) on rheumatoid arthritis: A systematic review. Int. J. Clin. Pract. 2021, 75, e13999. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.H.; Jin, C.; Zhao, Y.H. Study on chemical constituents of pomegranate flowers. Chin. Pat. Med. 2020, 42, 102–106. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, C.; Zhang, Y.; Li, Y.; Sun, J.-R. Immunomodulatory and antioxidant effects of pomegranate peel polysaccharides on immunosuppressed mice. Int. J. Biol. Macromol. 2019, 137, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.S.; Chen, H.; Chen, G. Study on the composition of Mulberry polysaccharide and its inducing apoptosis of tumor cells. J. Nutr. 2017, 39, 287–293. [Google Scholar] [CrossRef]
- Ji, X.L.; Hou, C.; Gao, Y.; Xue, Y.; Yan, Y.; Guo, X. Metagenomic analysis of gut microbiota modulatory effects of jujube (Ziziphus jujuba Mill.) polysaccharides in a colorectal cancer mouse model. Food Func. 2020, 11, 163–173. [Google Scholar] [CrossRef]
- Ye, W.; Fan, L.; Wang, Y. Effect of jujube polysaccharide on blood glucose and blood lipids in diabetic mice. Mod. Food Technol. 2016, 1, 6–12 + 5. [Google Scholar] [CrossRef]
- Xie, Y.T.; Luo, W.; Yang, J. Hypoglycemic effects of Jujube polysaccharides on streptozotocin induced diabetic mice. Food Sci. Technol. 2018, 9, 244–250. [Google Scholar]
- Ji, X.L.; Peng, B.X.; Ding, H.; Cui, B.; Nie, H.; Yan, Y.Z. Purification, structure and biological activity of pumpkin (Cucurbita moschata) polysaccharides: A review. Food Rev. Int. 2021, 11, 1904973. [Google Scholar] [CrossRef]
- Ayixiaguli, B.; Hu, S.; Cheng, L. Anti-lewis lung cancer activity of acidic polysaccharides BRAP-2 from turnip in vivo. J. Food Saf. Qual. 2019, 10, 5111–5116. [Google Scholar]
- Gunana, D. Isolation, purification and structure analysis of Polysaccharides from the Ancient Tibetan medicine. Master’s Thesis, Xinjiang Medical University, Xinjiang, China, 2014. [Google Scholar]
- Paerhati, M.; Ling, H.; Zhu, Q.; Ayitulun, S. Study on antitumor effect of Kunlun on colon cancer cell lines in vitro. Anhui Agr. Sci. 2015, 20, 146–148. [Google Scholar] [CrossRef]
- Wu, Z.; Li, H.; Wang, Y. Optimization extraction, structural features and antitumor activity of polysaccharides from Z. jujuba cv. Ruoqiangzao seeds. Int. J. Biol. Macromol. 2019, 135, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Lin, Q. Research progress of polysaccharides from Mulberry. J. Beijing Univ. 2016, 4, 63–66. [Google Scholar] [CrossRef]
- Xie, Y.; Jiang, S.; Su, D. Composition analysis and anti-hypoxia activity of polysaccharide from Brassica rapa L. Int. J. Biol. Macromol. 2010, 47, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.X. Mulberry Leaf Polysaccharide Separation and Purification, Structural Analysis and Biological Activity Research. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Zhao, W.H.; Ceng, C.; Qin, D.M. Study on the preventive effect and mechanism of Polysaccharide from Chamaejasme kunlun on acute liver injury induced by carbon tetrachloride in mice. Ethn. Med. 2017, 28, 4407–4410. [Google Scholar] [CrossRef]
- Shu, G.W.; Qiu, Y.H.; Fu, Q. Protective effect of Total Polysaccharides from Mulberry on acute liver injury induced by paracetamol in mice. J. S Cent. Univ. Natl. 2019, 38, 377–382. [Google Scholar]
- Liu, Y.; Tan, L.; Lan, L. Research progress on chemical constituents, pharmacological effects and quality markers of Mulberry. Chongqing Med. Sci. 2021, 50, 1063–1067. [Google Scholar] [CrossRef]
- Li, R.; Xue, Z.; Jia, Y. Polysaccharides from mulberry (Morus alba L.) leaf prevents obesity by inhibiting pancreatic lipase in high-fat diet induced mice. Int. J. Biol. Macromol. 2021, 192, 452–460. [Google Scholar] [CrossRef]
- Bian, L.; Chen, H.G.; Gong, X.J.; Zhao, C.; Zhou, X. Mori Fructus Polysaccharides Attenuate Alcohol-Induced Liver Damage by Regulating Fatty Acid Synthesis, Degradation and Glycerophospholipid Metabolism in Mice. Front. Pharm. 2021, 12, 766737. [Google Scholar] [CrossRef]
- Cheng, K.C.; Wang, C.J.; Chang, Y.C.; Hung, T.-W.; Lai, C.-J.; Kuo, C.-W.; Huang, H.-P. Mulberry fruits extracts induce apoptosis and autophagy of liver cancer cell and prevent hepatocarcinogenesis in vivo. J. Food Drug Anal. 2020, 28, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Cheng, W.; Bai, S.; Ye, L.; Du, J.; Zhong, M.; Liu, J.; Zhao, R.; Shen, B. White mulberry fruit polysaccharides enhance endothelial nitric oxide production to relax arteries in vitro and reduce blood pressure in vivo. Biomed. Pharmacother. 2019, 116, 109022. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.X.; Luo, X. Mulberry polysaccharide anti-fatigue effect and its mechanism. Chin. J. Exp. Pharm. 2012, 17, 234–236. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Han, X.; Dilnaz, K. Xinjiang kunlun snow chrysanthemum water extract on mice intestinal short chain fatty acid adjustment. Chin. J. Microecol. 2020, 37, 1134–1136. [Google Scholar] [CrossRef]
- Chen, W.; Lu, Y.; Hu, D.; Mo, J.; Ni, J. Black mulberry (Morus nigra L.) polysaccharide ameliorates palmitate-induced lipotoxicity in hepatocytes by activating nrf2 signaling pathway. Int. J. Biol. Macromol. 2021, 172, 394–407. [Google Scholar] [CrossRef]
- Tang, W.M.; Jin, L.; Xie, L.H. Isolation, purification, chemical structure and anti-fatigue animal study of polysaccharides from Turnip. Chin. J. Food Sci. 2018, 18, 22–31. [Google Scholar] [CrossRef]
- Tang, X. Effect of turnip polysaccharides on the inflammatory response in asthmatic rats. Chin. J. Med. Dr. 2012, 12, 1615–1617. [Google Scholar]
- Wu, S. Mulberry leaf polysaccharides suppress renal fibrosis. Int. J. Biol. Macromol. 2019, 124, 1090–1093. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.; Ye, B.P.; Xi, T. Progress in the polysaccharide of gray tree flowers. Pharm. Biotech. 2004, 11, 60–63. [Google Scholar]
- Wang, Z.Y.; Li, Z.W.; Bi, H.D.; Wang, F.; Wang, J.L.; Xiang, L. Optimization of synthesis of pomegranate seed polysaccharide selenate and its anti-lipid oxidation ability. Sci. Technol. Food Ind. 2019, 40, 149–162. [Google Scholar] [CrossRef]
- Gong, X.P. Structural Characterization and Biological Activity of Sulfation-Modified Jujube Polysaccharide. Ph.D. Thesis, Xinjiang Shihezi University, Xinjiang, China, 2021. [Google Scholar]
- Kimura, Y.; Sumiyoshi, M.; Suzuki, T. Anti tumor and antimetastatic activity of a novel water soluble low molecular weight beta-1,3-D-glucan (branch beta-1,6) isolated from Aureobasidium pullulans 1A1 strain black yeast (6B). Anticancer Res. 2006, 26, 4131–4141. [Google Scholar]
- Arken, T.; Huang, R.F.; Liu, X.F. Kunlun snow chrysanthemum genetic toxicity studies. Chin. J. Food Hyg. 2018, 30, 18–21. [Google Scholar] [CrossRef]
- Hailiqian, T.; Zulipiyan, A.; Li, Y.T. Determination of polysaccharide content and optimization of extraction process of Turnips in Xinjiang. J. Food Saf. Qual. 2021, 5, 1960–1965. [Google Scholar]
- Hairenguli, M.; Zulipiyan, A.; Hailiqian, T. Preliminary study on the hypoglycemic effect of turtups. J. Food Saf. Qual. 2020, 2, 387–392. [Google Scholar] [CrossRef]
- Zeng, C.; Yang, X.; Zhao, W. Kunlun snow chrysanthemum polysaccharide prevents acute and immunologically induced liver injury. Lishizhen Med. Mater. Med. Res. 2017, 11, 2604–2607. [Google Scholar]
- Hou, Y.; Li, G.; Wang, J. Okanin, effective constituent of the flower tea Coreopsis tinctoria, attenuates LPS-induced microglial activation through inhibition of the TLR4/NF-κB signaling pathways. Sci. Rep. 2017, 7, 45705. [Google Scholar] [CrossRef] [Green Version]
- Asaiguli. The Kunlun Snow Chrysanthemum Chemical Composition Analysis and Polysaccharide Extract and Active Research. Ph.D. Thesis, Gansu Agricultural University, Gansu, China, 2014. [Google Scholar]
- Li, Y.; Zhu, C.P.; Zhai, X.C.; Zhang, Y.; Duan, Z.; Sun, J.-R. Optimization of enzyme assisted extraction of polysaccharides from pomegranate peel by response surface methodology and their anti-oxidant potential. Chin. Herb. Med. 2018, 10, 416–423. [Google Scholar] [CrossRef]
- Liu, X.W.; Feng, H.Y. Effects of polysaccharides from pomegranate fruit on blood glucose and blood lipids in diabetic rats. J. Shandong Med. Sci. 2016, 38, 40–41. [Google Scholar] [CrossRef]
- Su, C.; Sun, S.; Wu, M. Effect of mulberry extract on LPS-induced inflammatory response and apoptosis in cardiomyocytes. Chin. J. Arterosclerosis 2022, 2, 125–129+146. [Google Scholar]
- Zhu, X.Y. Extraction and Purification of Polysaccharides from Mulberry and Its Protective Effect on Mice with Ulcerative Colitis. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2020. [Google Scholar]
- Mahemuti, K. Study on the antioxidant activity of turnip seed polysaccharide in vitro. Chin. J. Exp. 2014, 7, 118–121. [Google Scholar] [CrossRef]
- Geng, T.H. Studies on the content, structure and biological activity of polysaccharide in Chrysanthemum kunkunlun. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Chen, Z.E.; Wufuer, R.; Ji, J.H.; Li, J.F.; Cheng, Y.F.; Dong, C.X.; Taoerdahong, H. Structural Characterization and Immunostimulatory Activity of Polysaccharides from Brassica rapa L. J. Agric. Food Chem. 2017, 65, 9685–9692. [Google Scholar] [CrossRef] [PubMed]







| Name | Latin Name | Polysaccharide Components and Structures | Reference |
|---|---|---|---|
| Kunlun snow chrysanthemum | Coreopsis tinctoria Nutt. | KSCP1 is composed of Glc, Ara, Gal, and Xyl with a molar ratio of 10.53∶5.02∶4.96∶1 and a molecular weight range of 8200~8700 Da. KSCP2 is mainly composed of Glc, Ara, and Gal, with a molar ratio of 1∶2.78∶5.07 and a molecular weight range of 6100~6500 Da. Pectin polysaccharides: CTAP-1: [→4)GalpA(1→]n [→2)Rhap(1→]m m:n = 1:7 (n and m are numbers of monosaccharides, the same is said below) CTAP-2: [→4)GalpA(1→]n [→2)Rhap(1→]m m:n = 1:27 | [6] |
| Jujube | Ziziphus jujuba Mill. | Crude polysaccharide-1a (HJP-1a) is mainly composed of Ara and Gal with a ratio of 56.93 to 19.99. The average molecular weights of HJP-2, HJP-3, and HJP-4 are 4.590 × 104, 6.986 × 104, and 1.951 × 105 Da, respectively | [1,7,8,9] |
| Safflower | Carthamus tinctorius L. | At present, there are 11 kinds of polysaccharides isolated from safflower: SPS1, SPS2, SPS3, CTP, CTLP-1, CTLP-2, HH1-1, SPSa, SPAW, SF1, and SF2. SPS2 is composed of Rha, Ara, Xyl, Man, Glc, and Gal with a molar ratio of 4.44:1.46:4.51:5.82:8.23:19.38. The relative molecular weight of SPS3 is 5.86 × 103, and it is composed of Rha, Ara, Glc, and Gal, with a molar ratio of 2.93:11.19:33.68:3.48. CTP: Glc, Gal in a ratio of 6.08:1; CTLP-1: Ara, Glc, Gal in a ratio of 6.7:4.2:1; CTLP-2: Ara, Glc, Gal in a ratio of 16.76:4.28:1; HH1-1: Gal, Ara in a ratio of 54, 9:45.1; SPSa and SPAW: Glc; SF-1: Rha, Ara, Xyl, Man, Glc, Gal in a ratio of 2.9:7.5:3.8:1:11.6:8.9; SF-2: Rha, Ara, Xyl, Man, Glc, Gal in a rato of 2.9:10.3:4.2:1:5.1:8.5. | [10,11,12] |
| Chinese wolfberry | Lycium dasystemum Pojark. | Lycium barbarum polysaccharides (LBP) are composed of Man, Rha, GalA, Glc, Gal, and Ara, with a content ratio of 4.98∶2.93∶8.38∶22.44∶25.38∶35.89. There are 20 kinds of Chinese wolfberry polysaccharides currently found: LbGp2: 68,200 Da, Ara:Gal = 4:5. Backbone composed of (1→6)-β-Gal. Branches composed of (1→3)-β-Ara and (1→3)-β-Gal terminated with (1→3)/(1→5)-α-Ara. LbGp3: 92,500 Da, Ara:Gal = 1:1. Backbone composed of (1→4)-β-Gal. Branches composed of (1→3)-β-Ara and (1→3)-α-Gal terminated with (1→3)/(1→5)-α-Ara. LbGp4: 214,800 Da, Ara:Gal:Rha:Glc = 1.5:2.5:0.43:0.23. Backbone composed of (1→4)-β-Gal. Branches composed of (1→3)-β-Gal terminated with (1→3)-α-Ara and (1→3)-β-Rha. LBPA3: 66,000 Da, Ara:Gal = 1.2:1. Heteropolysaccharide with (1→4), (1→6). LBPB1: 18,000 Da, Ara:Glc = 1:3.1. Heteropolysaccharide with (1→4), (1→6) β-glycosidic bond. LBP-a4: 10,200 kDa, Fuc:gal = 0.41:1. LBPC2: 12,000 kDa, Xyl:Rha:Man = 8.8:2.3:1. Heteropolysaccharide with (1→4), (1→6) β-glycosidic bond. LBPC4: 10,000 kDa, Glc. Heteropolysaccharide with (1→4), (1→6) α-glycosidic bond. LBP1a-1: 115,000 Da, Glc, α-(1→6)-D-glucan. LBP1a-2: 94,000 Da, Glc, α-(1→6)-D-glucan. LBP3a-1: 103,000 Da, Polygalacturonan with (1→4)-α-glycosidic bond. LBP3a-1: 103,000 Da, Polygalacturonan with (1→4)-α-glycosidic bond. LBP3a-2: 82,000 Da, GalA, composed of a small amount of Gal and Ara Polygalacturonan with (1→4)-α-glycosidic bond. LBLP5-A: 113,300 Da, (1→3)-linked Gal, (1→4)-linked Gal, (1→3)-linked Ara, (1→5)-linked Ara, and (1→2, 4)-linked Rha. WSP: Rha:Fuc:Ara:Xyl:Man:Gal:Glc = 1.6:0.2:51.4:4.8:1.2:25.9:7.3. Backbone composed of (1→2)-linked-Rha and (1→4)-linked-Gal. Branches composed of (1→5)-linked-Ara terminated with Ara residues, and (1→4)-linked-Xyl terminated with Man residues. AGP: Rha:Ara:Xyl:Gal:Glc:GalA:GlcA = 3.3:42.9:0.3:44.3:2.4:7. Backbone composed of linear homogalacturonan fragments and rhamnogalacturonan fragments. Side chains mainly composed of β-1,6- and β-1,4-galactopyranan and α-1,5-arabinofuranan. LBP-IV: 41,800 Da, Rha:Ara:Xyl:Glc:Gal = 1.61:3.82:3.44: 7.54:1. Backbone composed of both α- and β-anomeric configurations of Ara and Glc. Rha was located at terminal of polysaccharide chain. LbGp1: 49,100 Da, Ara:Gal = 5.6:1. Backbone composed of (1→6)-Gal. Side chains mainly composed of (1→3)-Gal/(1→4)-Gal and (1→3)-Ara/(1→4)-Ara. Ara was located at terminal of branch. P-LBP: 64,000 Da, Fuc:Rha:Ara:Gal:Glc:Xyl:GalA:GlcA = 1.00:6.44:54.84:22.98:4.05: 2.95:136.98:3.35. Backbone composed of (1→4)-α-GalA. Side chains mainly composed of α-1,2- and α−1,4-Rha and α-1,5-Ara. LBP1B-S-2: 80,000 Da, Rha:Ara:Gal:Glu = 3.13:53.55:39.37:3.95. Backbone composed of 1, 3-linked β-D-Gal and 1, 6-linked β-D-Gal. Branches contain 1, 4-linked β-D-GlcA; T-linked β-D-Gal; 1, 6-linked β-D-Gal; T-linked α-L-Ara; T-linked β-L-Ara; 1, 5-linked α-L-Ara; and T-linked β-L-Rha. LRGP1: 56,200 Da, Rha:Ara:Xyl:Man:Glu:Gal = 0.65:10.71:0.33:0.67:1:10.41. Backbone composed of (1→3)-linked Gal. The branches were composed of (1→5)-linked Ara, (1→2)-linked Ara, (1→6)-linked Gal, (1→3)-linked Gal, (1→4)-linked Gal, and (1→2,4)-linked Rha. (1→3)-β-Gal terminated with (1→3)-α-Ara and (1→3)-β-Rha. LBP-a4: 10,200 Da, Fuc:Gal = 0.41:1 LBPC2: 12,000 Da, Xyl:Rha:Man = 8.8:2.3:1. Heteropolysaccharide with (1→4), (1→6) β-glycosidic bond. LBPC4: 10,000 Da, Glc. Heteropolysaccharide with (1→4), (1→6) α-glycosidic bond. | [13,14,15] |
| Herba cistanche | Cistanche deserticola Ma. | Herba cistanche is composed of one neutral polysaccharide (CTZ) and five acidic polysaccharides (CT1, CT2, CT3, CT4, and CT5), with contents of 299.2, 168.0, 123.2, 121.6, 54.4 and 11.2 mg/g, respectively. | [16] |
| Sea-buckthorn | Hippophae rhamnoides Linn. | Sea-buckthorn polysaccharides contain the neutral polysaccharide SBP-I and the acidic polysaccharides SBP-II, SBP-III, and SP0.1-1. SBP-I is composed of Ara, Xyl, Man, Glc, and Gal with a molar ratio of 1.18∶1∶2.20∶32.17∶1.45. SBP-II is composed of Xyl, Man, Glc, and Gal with a molar ratio of 1∶0.28∶1.02∶0.20. SBP-III consists of Xyl, Glc, and Gal with a molar ratio of 1∶2.15∶0.28. SP0.1-1 is composed of Man, Glc, Gal, and Ara in a molar ratio of 1:2.3:1.9:11.2, with a core structure containing 1,4-linked-α-D-Glc; 1,4, 6-linked-α-D-Glc; and 1,4-linked-α-D-Man residues as the backbone. The side-chains are composed of 1,3,5-linked-α-L-Ara; 1,5-linked-α-L-Ara; terminal α-Ara; and 1,4-linked-β-D-Gal. | [17,18,19] |
| Turnip | Brassica rapa L. | Acidic polysaccharides in turnips consist of D-GalA, D-Man, L-Ara, D-Gal, D-Glc, L-Rha, D-GlcA = 67.73∶17.19∶10.20∶3.19∶1.11∶0.34:0.23. Brassica rapa L. acidic polysaccharide (BRAP): BRAP-1: Ara:Glc:Gal:GalA = 2.07:4.53:2.20:1; BRAP-2: Rha:Ara:Glc:Gal:GalA = 1.06:5.03:2.22:1.5. | [7,20] |
| Chicory | Cichorium intybus L. | The main components of chicory polysaccharides are a kind of fructan with similar structure. This fructan is a straight chain polysaccharide with a β-2, 1-glycosidic bond between fructose residues (F) and a glucose residue (G) at the end. The structural formula is G-1, (2-F-1) N-1, 2-F. In addition, chicory polysaccharides also contain a small amount of inulonose, that is, fructoses without a G terminal; the structure is F-1, (2-F-1) N-2, 2-F. | [21] |
| Mulberry | Morus alba L. | Three kinds of polysugars are separated from mulberry, namely, MFP-1, MFP-2, MFP-3 and MFP-4, which are composed of Ara, Gal, Glc, Gly, Xyl, and GalA. MFP3 glycosidic bond types include:(1→6)-linked α-D-Glc, (1→2)-linked α-L-Rha, (1→3)-linked α-D-Gal, (1→3)-linked β-L-Rha and (1→)-linked α-L-Ara. | [22,23] |
| Garlic | Allium sativum L. | Garlic polysaccharides are mainly composed of Ara, Gal, Glc, Xyl, Fru, and other monosaccharides. | [24,25,26] |
| Basil | Ocimum basilicum L. | Pure basil polysaccharide (OBP) is composed of Rha, Ara, Fuc, Alo, Man, and Gal, and the relative molar ratio is 2.18:4.025:2.38:0.15:1.7:0.357. | [27,28] |
| Alfalfa | Medicago Sativa Linn. | Alfalfa polysaccharide components include APS-2a, APS-2b, APS-3a, APS-3b, and APPS. APS-3b has the most complex monosaccharide composition. The monosaccharide composition and molar ratio are Xyl:Ara:Glc:Rha:Gal:GlcA:GalA = 1.00:2.35:3.78:3.05.2.74:1.72:11.45. APS-3a has the highest content of polysaccharide and uronic acid, at 92.88% and 52.09%, respectively. Pectic polysaccharide (APPS) is characterized to be a rhamnogalacturonan I (RG-I) type pectin with a molecular weight of 2.38 × 103 Da and a radius of 123 nm. Primary structural analysis indicates that APPS is composed of a (1→2)-α-L-Rha-(1→4)-α-D-GalA-(1→2) backbone with 12% branching point at C-4 of Rha forming side chains by L-arabinosyl and D-galactosyl oligosaccharide units. | [29,30] |
| Pomegranate | Punica granatum L. | Polysaccharide components of pomegranate peel I: Ara, GalA, Gal, Rha, Glc, GlcA, Man. (Ara > GalA > Gal > Rha > Glc > GlcA > Man) Polysaccharide components of pomegranate peel II: Ara, Gal, Rha, Glc, GalA, Xyl, Man, and Fuc. (Ara > Gal > Rha > Glc > GalA > Xyl > Man > Fuc) | [31,32] |
| Resina ferulae | Ferula sinkiangensis K. M. Shen. | Total polysaccharides of Avegeron (FSPt): FSP30, FSP50, FSP70, FSP80 Pleurotus ferulae lenzi polysaccharide: PFLP1 contains four kinds of monosaccharide (Rha, Man, Glu, and Gal), in a molar ratio of 1:1.54:18.6:3.64; PFLP2 contains five monosaccharides (Rha, Xyl, Man, Glu, and Gal), with a molar ratio of 1:1.08:0.65:6.76:4.28. PFLP1:[→3)-β-D-Glu-(1→3)-β-D-Glu-(1→3)-β-D-Man p-(1→3)-β-D-Glu-(1→3)-β-D-Glu-(1→3)-β-D-Glu-( 1→3)-β-D-Glu-(1→3)-β-D-Glu-(1→2)-β-D-Rha-(1 →3)-β-D-Glu-(1→3)-β-D-Glu-(1→3)-β-D-Glu-(1→ 3)-β-D-Glu-(1→3)-β-D-Glu-(1→3)-β-D-Man-(1→3) -β-D-Glu-(1→3)-β-D-Glu-(1→]n PFLP2:[→3)-β-D-Glu-(1→3)-β-D-Glu-(1→4)-β-D-Xyl -(1→3)-β-D-Glu-(1→3)-β-D-Glu-(1→2)-β-D-Rha-(1 →3)-β-D-Glu-(1→3)-β-D-Glu-(1→3)-β-D-Man-(1 →3)-β-D-Glu-(1→]n | [33] |
| Korla pear | Pyrus sinkiangensis Yu. | Korla pear polysaccharides (PSP) are divided into Korla pear acidic polysaccharide (PSAP) and Korla pear neutral polysaccharide (PSNP). PSNP-1: Glc:Xyl:Gal = 3:1.6:1, in addition to a small amount of Ara, Fuc, and Man. PSNP-2: Glc:Xyl:Ara:Gal = 2.4:1:2.5:1.1, in addition to a small amount of Fuc and Man. PSAP-1 is mainly composed of GalA and contains a small amount of Ara and Rha. PSAP-2 is mainly composed of GalA and contains a small amount of Ara and Rha. | [34,35,36,37] |
| Hops | Humulus Lupulus L. | Hops polysaccharides: HLP50-1, HLP50-2, HLP70-2-1, HLP70-2-2, HLP70-3, HLBP-2, HLBP-3, and HLBP-4 The molecular weights are 49.13 kDa, 73.25 kDa, 11.12 kDa, 7.3 kDa, 26.48 kDa, 6.13 kDa, 35.23 kDa, and 33.12 kDa, respectively. | [25] |
| Fritillaria | Fritillaria walujewii Regel | Fritillaria polysaccharide (FPSP) is composed of Xyl, Glc, Gal, and Man, with a molar ratio of 1: 58.02: 0.73 | [38] |
| Dandelion | Taraxacum altaicum Schischk. | Dandelion polysaccharide: TMP-1 without uronic acid is neutral and its purity is 95.2%. TMP-2 contains uronic acid as an acidic polysaccharide, and its purity is 85.9%. Dandelion root polysaccharide (DRP): DRP-2b: with a molecular weight of 31.8 kDa, is composed of Rha, GlcA, Glc, Gal, and Ara; DRP-3a: with a molecular weight of 6.72 kDa, is composed of Rha, Glc, Gal, and Ara. The backbones of DRP-2b and DRP-3a are mainly composed of (1→5)-linked-α-D-Ara and (1→6)-linked-α-D-Glc, respectively. | [39,40] |
| Lilium | Lilium martagon var. pilosiusculum Freyn | Lilium polysaccharides are LLPS-1, LLPS-2, and LLPS-3. The average molecular weight of the three lilium polysaccharides are 350.5 kDa, 403.3 kDa, and 146.2 kDa, respectively. | [41,42] |
| Selfheal | Prunella vulgaris L. | Selfheal polysaccharides: PV-P1, PV-P2, PV-P3. PV-P1 Mainly constitued by →5)-Ara-(1→,→3)-Xyl-(1 and 6)-Glc-(1- residue composition, and has a molar ratio of 125:29.1:3.2. In addition, it contains the terminal mannose, terminal galactose, 3-Gal-(1→,→4,6)-Glc (1→,and→3,6) -Gal-(1→ residue composition. PV-P2 is mainly composed of terminal galacto, →3)-Xyl-(1→,→6)-Glc-(1→,→3)-Gal-(1→,→5)-Ara-(1→ with a molar ratio of 23: 8.1:29:3.5:4.1:38. PV-P3 mainly consists of →3)-Xyl-(1→,→5)-Ara-(1→ and terminal mannose, with a molar ratio of 59:26:2.7. | [2] |
| Type | Monosaccharide | Chemical Formula | Molecular Weight | Structural Formula |
|---|---|---|---|---|
| Common | Ara | C5H10O5 | 150.13 |  |
| Gal | C6H12O6 | 180.16 |  | |
| Glc | C6H12O6 | 180.16 |  | |
| Different | Xyl (Mulberry, Pomegranate, Kunlun snow chrysanthemum) | C5H10O5 | 150.13 |  |
| Glc-A (Turnip, Pomegranate) | C6H10O7 | 194.14 |  | |
| Gal-A (Mulberry) | C6H10O7 | 194.14 |  | |
| Man (Turnip, Pomegranate) | C6H12O6 | 180.16 |  | |
| Rha (Pomegranate) | C6H12O5 | 164.16 |  | |
| Fuc (Pomegranate) | C6H12O5 | 164.16 |  |
| Plants | Extraction Site | Extaction | Separation and Purification | Reference |
|---|---|---|---|---|
| Turnip | Root | Reflux extraction, Ethanol subsiding method | DEAE-cellulose, Sephadex G-100 column, Gel permeation chromatography (GPC) | [51] |
| DAED-650M, Sepharese 6B, Sephacry1S-300 | [20] | |||
| DEAE-52 cellulose, Sephadex G-100 column | [7] | |||
| HP-100 macroporous resin, DEAE-52 cellulose, Sephadex G-150 column | [52] | |||
| Seed | - | [53] | ||
| Kunlun snow chrysanthemum | Flowers | Reflux extraction, Ethanol subsiding method | DEAE-650M, Sepharose-6B column (5.0 i.d. × 100 cm), Sephacryl S-300 HR column (2.2 i.d. × 90 cm), PL aquagel-OH MIXED-H column (7.5 m × 300 mm, 8µm) on an Agilent 1200 system, | [6] |
| DEAE-52 Cellulose, Sephacryl S-100, Sephadex G-75 column | [20] | |||
| Complex enzyme-ultrasonic-assisted extraction | DEAE-52 Cellulose ion exchange, Sephadex G-100 Dextran gel column | [54] | ||
| Ultrasonic-assisted hot water extraction–alcohol precipitation method | DEAE-52 chromatography column, Sephadex G-100 gel chromatography column, UV spectroscopy, Freeze–thaw analysis, Sephadex G-100 gel column chromatography | [6] | ||
| Pomegranate | Seed | Enzymatic method (pectinase) | - | [31] |
| Hot water extraction, ultrasonic-assisted water extraction and compound enzyme method | - | [32] | ||
| Polyethylene glycol combined with ultrasonic microwave-assisted extraction technology | High-speed countercurrent chromatography(HSCCC), Gel permeation chromatography(GPC) | [50] | ||
| Skin | Reflux extraction, Ethanol subsiding method | DEAE-cellulose column | [51] | |
| Mulberry | Fruits | Reflux extraction, Ethanol subsiding method | DEAE-52 Cellulose column, Sephadex G-100 Dextran gel column | [24] |
| Reflux extraction, ultrasonic-assisted extraction, Enzyme-assisted extraction, High-speed shear-technology-assisted extraction | - | [26] | ||
| Oil ether reflux degreasing combined with water extraction and alcohol precipitation | AB-8 macroporous resin, DEAE cellulose column, SepharseCL-B gel column layer | [52] | ||
| Jujube | Fruits | Ethanol subsiding method | Anion exchange, Sepharose CL-6B column, High-performance gel permeation chromatography | [53] |
| Ultrasonic extraction | Ion exchange, Gel permeation chromatography | [54] | ||
| DEAE-52 cellulose column, Sephadex G-100 Dextran gel column | [55] | |||
| Ethanol subsiding method | DEAE-52 cellulose column, Sephadex G-100 column | [54] | ||
| Hot water extraction, Ultrasonic-assisted extraction | DEAE-52 anion exchange column, Sephadex G-100 column | [55] | ||
| Cellulase method | - | [53] | ||
| Leaf | Ultrasonic extraction | - | [8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taoerdahong, H.; Kadeer, G.; Chang, J.; Kang, J.; Ma, X.; Yang, F. A Review Concerning the Polysaccharides Found in Edible and Medicinal Plants in Xinjiang. Molecules 2023, 28, 2054. https://doi.org/10.3390/molecules28052054
Taoerdahong H, Kadeer G, Chang J, Kang J, Ma X, Yang F. A Review Concerning the Polysaccharides Found in Edible and Medicinal Plants in Xinjiang. Molecules. 2023; 28(5):2054. https://doi.org/10.3390/molecules28052054
Chicago/Turabian StyleTaoerdahong, Hailiqian, Gulimila Kadeer, Junmin Chang, Jinsen Kang, Xiaoli Ma, and Fei Yang. 2023. "A Review Concerning the Polysaccharides Found in Edible and Medicinal Plants in Xinjiang" Molecules 28, no. 5: 2054. https://doi.org/10.3390/molecules28052054