
Antioxidant and Antiproliferative Activities of Kale (Brassica oleracea L. Var. acephala DC.) and Wild Cabbage (Brassica incana Ten.) Polyphenolic Extracts
 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Kale and Wild Cabbage Plants
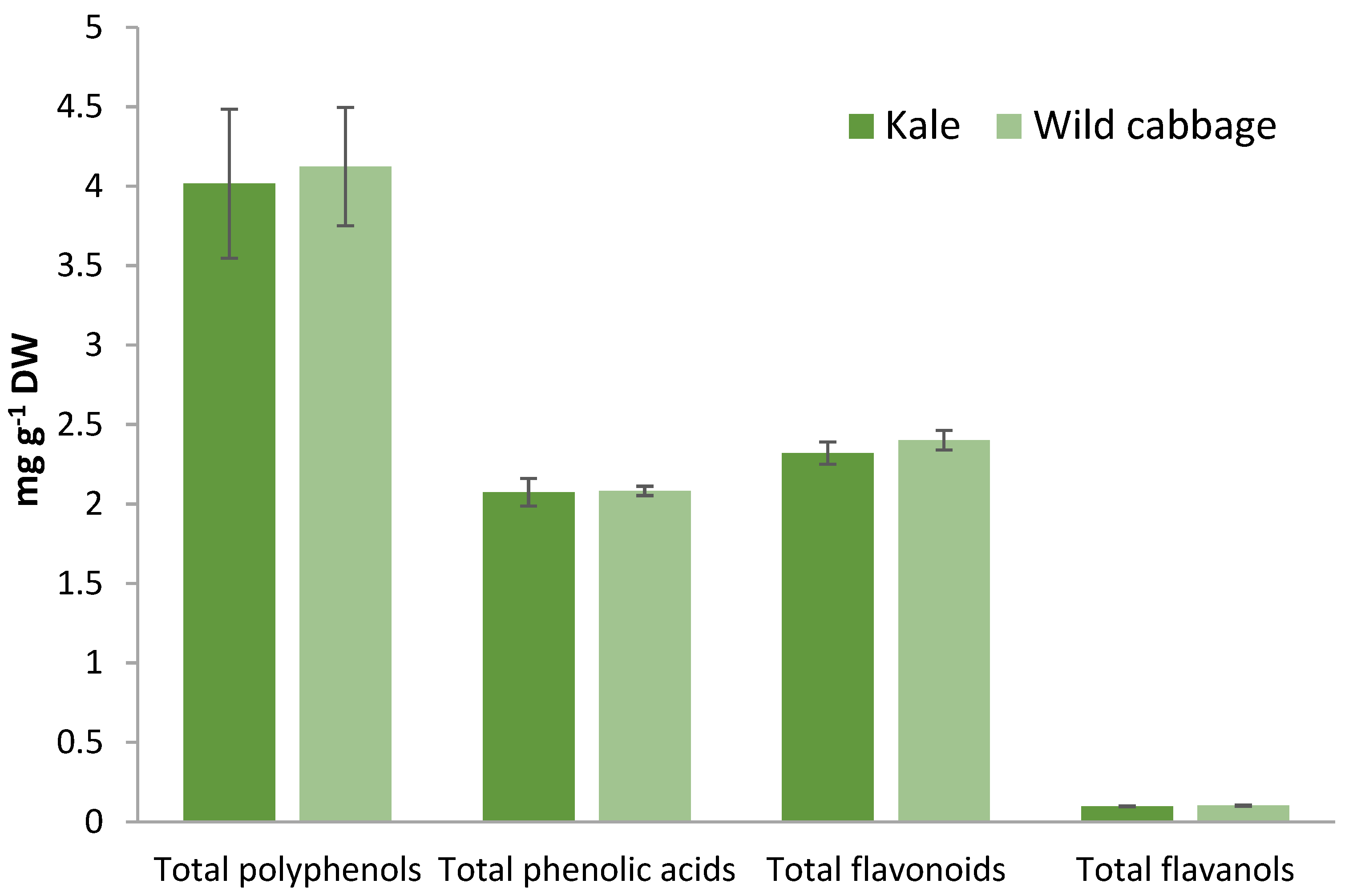
2.2. Polyphenolic Compounds
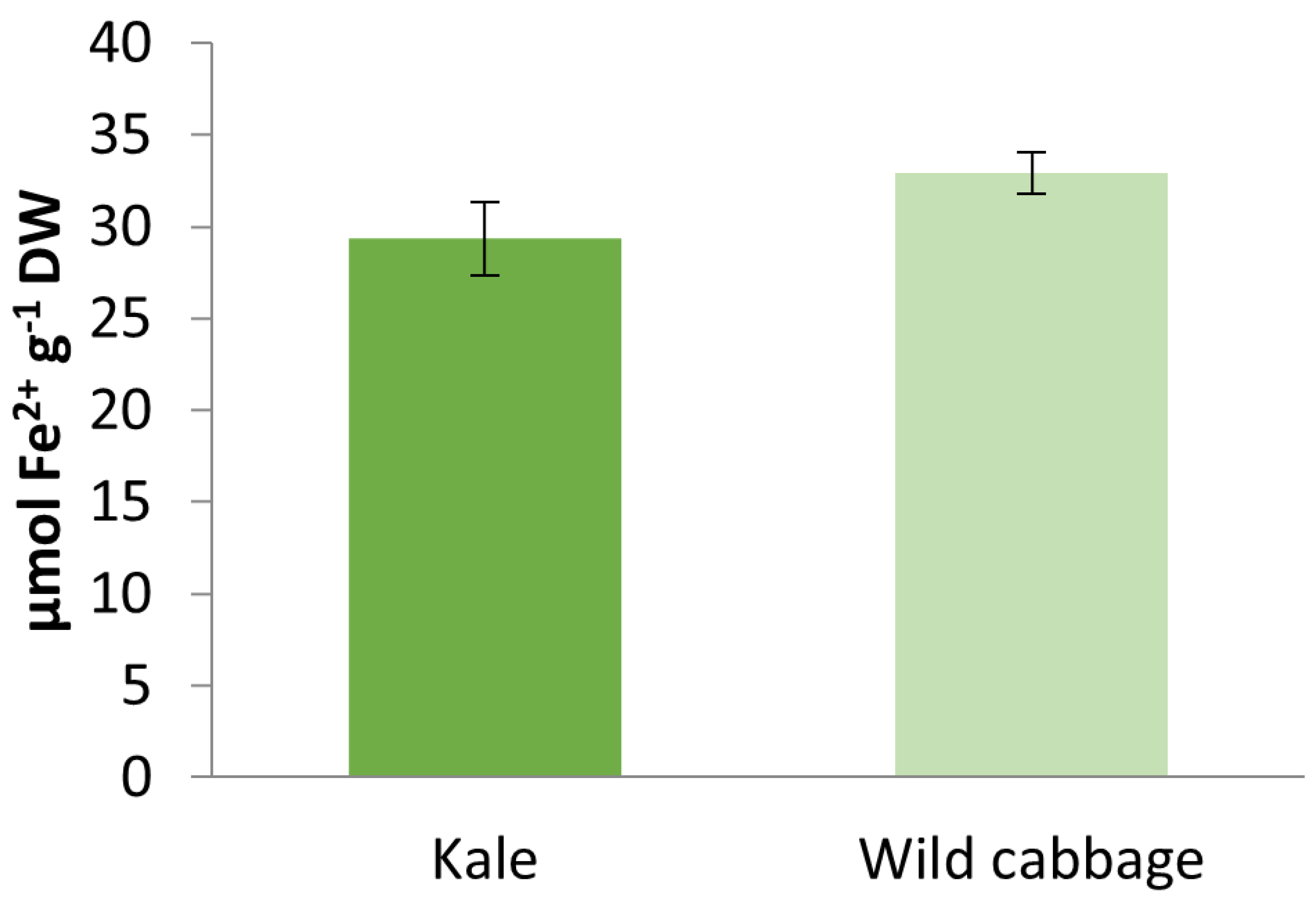
2.3. Antioxidant Activity
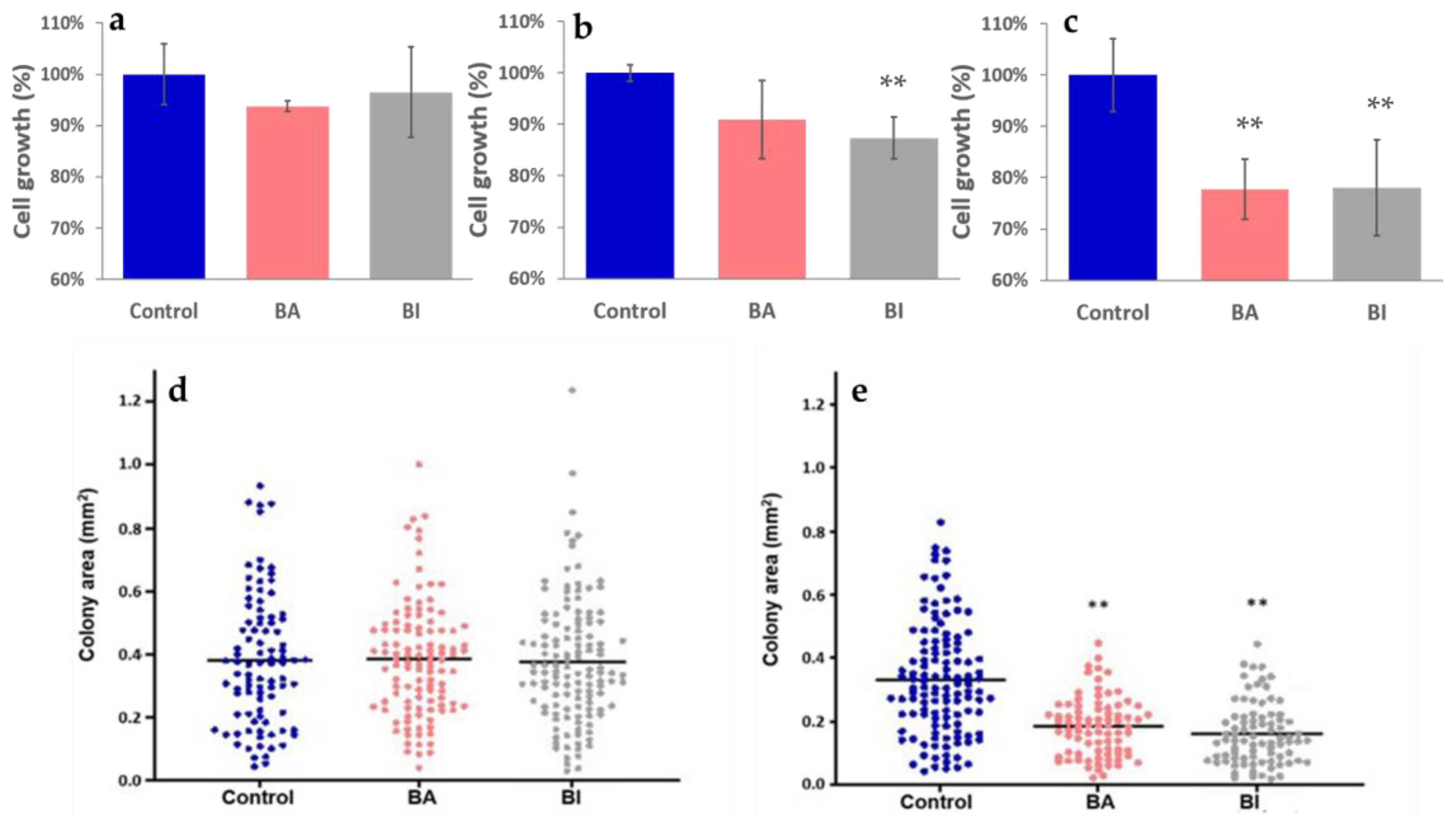
2.4. The Effect of Kale and Wild Cabbage Plant Extracts on HeLa Cells
3. Discussion
4. Materials and Methods
4.1. Plant Growth
4.2. Analysis of Polyphenolic Compounds
4.2.1. Extraction of Polyphenolic Compounds
4.2.2. Spectrophotometry of Polyphenolic Compounds and Antioxidant Activity
4.2.3. LC-MS/MS Analysis of Selected Polyphenolic Compounds
4.3. Biological Activity Analysis of Brassica Plant Extracts
4.3.1. Cell Culture and Treatments
4.3.2. Cell Growth Analysis
4.3.3. Colony-Forming Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Batelja, K.; Ban, S.G.; Žanić, K.; Miloš, B.; Dumičić, G.; Matotan, Z. Autochtonous kale populations (Brassica oleraceae L. var. acephala) in Croatian coastal region. Poljoprivreda 2009, 15, 8–14. [Google Scholar]
- Snogerup, S.; Gustafsson, M.; Von Bothmer, R. Brassica sect. Brassica (Brassicaceae) I. Taxonomy and variation. Willdenowia 1990, 19, 271–365. [Google Scholar]
- Jasprica, N. Brassica L. In Endemi u Hrvatskoj flori; Nikolić, T., Milović, M., Bogdanović, S., Jasprica, N., Eds.; Alfa: Zagreb, Croatia, 2015; pp. 107–115. [Google Scholar]
- Mabry, M.E.; Turner-Hissong, S.D.; Gallagher, E.Y.; McAlvay, A.C.; An, H.; Edger, P.P.; Moore, J.D.; Pink, D.A.C.; Teakle, G.R.; Stevens, C.J.; et al. The Evolutionary history of wild, domesticated, and feral Brassica oleracea (Brassicaceae). Mol. Biol. Evol. 2021, 38, 4419–4434. [Google Scholar] [CrossRef] [PubMed]
- Miceli, N.; Cavò, E.; Ragusa, M.; Cacciola, F.; Mondello, L.; Dugo, L.; Acquaviva, R.; Malfa, G.A.; Marino, A.; D’Arrigo, M.; et al. Brassica incana Ten. (Brassicaceae): Phenolic constituents, antioxidant and cytotoxic properties of the leaf and flowering top extracts. Molecules 2020, 25, 1461. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Pavlović, I.; Salopek-Sondi, B. White cabbage (Brassica oleracea var. capitata f. alba): Botanical, phytochemical and pharmacological overview. Phytochem. Rev. 2017, 16, 117–135. [Google Scholar] [CrossRef]
- Salehi, B.; Quispe, C.; Butnariu, M.; Sarac, I.; Marmouzi, I.; Kamle, M.; Tripathi, V.; Kumar, P.; Bouyahya, A.; Capanoglu, E.; et al. Phytotherapy and food applications from Brassica genus. Phytother. Res. 2021, 35, 3590–3609. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251. [Google Scholar] [CrossRef]
- Raiola, A.; Errico, A.; Petruk, G.; Monti, D.M.; Barone, A.; Rigano, M.M. Bioactive compounds in Brassicaceae vegetables with a role in the prevention of chronic diseases. Molecules 2018, 23, 15. [Google Scholar] [CrossRef] [Green Version]
- Sanlier, N.; Saban, M.G. The benefits of Brassica vegetables on human health. J. Human Health Res. 2018, 1, 104. [Google Scholar]
- Šamec, D.; Salopek-Sondi, B. Cruciferous (Brassicaceae) Vegetables. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Silva, T.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 195–202. [Google Scholar] [CrossRef]
- Quirante-Moya, S.; García-Ibañez, P.; Quirante-Moya, F.; Villaño, D.; Moreno, D.A. The role of Brassica bioactives on human health: Are we studying it the right way? Molecules 2020, 25, 1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favela-González, K.M.; Hernández-Almanza, A.Y.; De la Fuente-Salcido, N.M. The value of bioactive compounds of cruciferous vegetables (Brassica) as antimicrobials and antioxidants: A review. J. Food Biochem. 2020, 44, e13414. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.J.; Yang, Y.; Vogtmann, E.; Wang, J.; Han, L.H.; Li, H.L.; Xiang, Y.B. Cruciferous vegetables intake and the risk of colorectal cancer: A meta-analysis of observational studies. Ann. Oncol. 2013, 24, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.J.; Yang, Y.; Wang, J.; Han, L.H.; Xiang, Y.B. Cruciferous vegetable consumption and gastric cancer risk: A meta-analysis of epidemiological studies. Cancer Sci. 2013, 104, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Mao, Q.; Lin, Y.; Zhou, F.; Xie, L. The association of cruciferous vegetables intake and risk of bladder cancer: A meta-analysis. World J. Urol. 2013, 31, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Mao, Q.; Wang, X.; Zhou, F.; Luo, J.; Wang, C.; Lin, Y.; Zheng, X.; Xie, L. Cruciferous vegetables consumption and risk of renal cell carcinoma: A meta-analysis. Nutr. Cancer 2013, 65, 668–676. [Google Scholar] [CrossRef]
- Wu, Q.J.; Xie, L.; Zheng, W.; Vogtmann, E.; Li, H.L.; Yang, G.; Ji, B.T.; Gao, Y.T.; Shu, X.O.; Xiang, Y.B. Cruciferous vegetables consumption and the risk of female lung cancer: A prospective study and a meta-analysis. Ann. Oncol. 2013, 24, 1918–1924. [Google Scholar] [CrossRef]
- Liu, B.; Mao, Q.; Cao, M.; Xie, L. Cruciferous vegetables intake and risk of prostate cancer: A meta-analysis. Int. J. Urol. 2012, 19, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lv, K. Cruciferous vegetables intake is inversely associated with risk of breast cancer: A meta-analysis. Breast 2013, 22, 309–313. [Google Scholar] [CrossRef]
- Han, B.; Li, X.; Yu, T. Cruciferous vegetables consumption and the risk of ovarian cancer: A meta-analysis of observational studies. Diagn. Pathol. 2014, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Zirpoli, G.R.; McCann, S.E.; Moysich, K.B.; Ambrosone, C.A.; Tang, L. Trends in Cruciferous vegetable consumption and associations with breast cancer risk: A case-control study. Curr. Dev. Nutr. 2017, 1, e000448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, D.; Abellán-Victorio, A.; Beretta, V.; Camargo, A.A.; Moreno, D.A. Functional ingredients from Brassicaceae species: Overview and perspectives. Int. J. Mol. Sci. 2020, 21, 1998. [Google Scholar] [CrossRef] [PubMed]
- Velasco, L.; Becker, H.C. Variability for seed glucosinolates in a germplasm collection of the genus Brassica. Genet. Resour. Crop Evol. 2000, 47, 231–238. [Google Scholar] [CrossRef]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Linić, I.; Šamec, D.; Grúz, J.; Bok, V.V.; Strnad, M.; Salopek-Sondi, B. Involvement of phenolic acids in short-term adaptation to salinity stress is species specific among Brassicaceae. Plants 2019, 8, 155. [Google Scholar] [CrossRef] [Green Version]
- Mageney, V.; Neugart, S.; Albach, D.C. A guide to the variability of flavonoids in Brassica oleracea. Molecules 2017, 22, 252. [Google Scholar] [CrossRef] [Green Version]
- Fratianni, F.; Cardinale, F.; Cozzolino, A.; Granese, T.; Pepe, S.; Riccardi, R.; Spigno, P.P.; Coppola, R.; Nazzaro, F. Polyphenol composition and antioxidant activity of two autochthonous Brassicaceae of the Campania region, Southern Italy. Food Sci. Nutr. 2014, 5, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.; Sehwag, S.; Singh, S.P. Antioxidant activity and polyphenol content of Brassica oleracea varieties. Int. J. Veg. Sci. 2016, 22, 353–363. [Google Scholar] [CrossRef]
- Šamec, D.; Pavlović, I.; Radojčić-Redovniković, I.; Salopek-Sondi, B. Comparative analysis of phytochemicls and activity of endogenous enzymes associated with their stability, bioavaibility and food quality in five Brassicaceae sprouts. Food Chem. 2018, 269, 96–102. [Google Scholar] [CrossRef]
- Šamec, D.; Kruk, V.; Ivanišević, P. Influence of seed origin on morphological characteristics and phytochemicals levels in Brassica oleracea var. acephala. Agronomy 2019, 9, 502. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Linić, I.; Salopek-Sondi, B. Salinity stress as an elicitor for phytochemicals and minerals accumulation in selected leafy vegetables of Brassicaceae. Agronomy 2021, 11, 361. [Google Scholar] [CrossRef]
- Khalid, W.; Arshad, M.S.; Imran, M.; Ahmad, R.S.; Imran, A.; Qaisrani, T.B.; Asghar, Z.; Husain, A.; Anjum, F.M.; Suleria, H.A.R. Kale (Brassica oleracea var. sabellica) as miracle food with special reference to therapeutic and nutraceuticals perspective. Food Sci. Nutr. 2021, 10, 3175. [Google Scholar] [CrossRef]
- Linić, I.; Mlinarić, S.; Brkljačić, L.; Pavlović, I.; Smolko, A.; Salopek-Sondi, B. Ferulic Acid and Salicylic Acid Foliar Treatments Reduce Short-Term Salt Stress in Chinese Cabbage by Increasing Phenolic Compounds Accumulation and Photosynthetic Performance. Plants 2021, 10, 2346. [Google Scholar] [CrossRef]
- Tripodi, G.; Verzera, A.; Dima, G.; Condurso, C.; Ragusa, S. Brassica fruticulosa Cyr. and Brassica incana Ten. (Brassicaceae) as Mediterranean traditional wild vegetables: A valuable source of bioactive compounds. J. Essent. Oil Res. 2012, 24, 539–545. [Google Scholar] [CrossRef]
- Lukić, I.; Išić, N.; Ban, D.; Sondi, B.S.; Ban, S.G. Comprehensive Volatilome Signature of Various Brassicaceae Species. Plants 2023, 12, 177. [Google Scholar] [CrossRef]
- Horn, P.J.; Vaughan, J.G. Seed glucosinolates of fourteen wild Brassica species. Phytochemistry 1983, 22, 465–471. [Google Scholar] [CrossRef]
- Heaney, R.K.; Fenwick, G.R.; Mithen, R.F.; Lewis, B.G. Glucosinolates of wild and cultivated Brassica species. Phytochemistry 1987, 26, 1969–1973. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Bok, V.V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Anti-radical activity and polyphenol composition of local Brassicaceae ediable varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Sikora, E.; Bodziarczyk, I. Composition and antioxidant activity of kale (Brassica oleracea L. var. acephala) raw and cooked. Acta Sci. Pol. Technol. Aliment. 2012, 11, 239–248. [Google Scholar] [PubMed]
- Tajalli, F.; Saeedi, M.; Malekabadi, A.V. Anticancer and Antioxidant Effects of Red Cabbage on Three Cancerous Cell Lines and Comparison with a Normal Cell Line (HFF-3). J. Genes Cells 2020, 6, 12–20. [Google Scholar] [CrossRef]
- Mansour, K.A.; Moustafa, S.F.; Abdelkhalik, S.M. High-Resolution UPLC-MS Profiling of Anthocyanins and Flavonols of Red Cabbage (Brassica oleracea L. var. capitata f. rubra DC.) Cultivated in Egypt and Evaluation of Their Biological Activity. Molecules 2021, 26, 7567. [Google Scholar] [CrossRef] [PubMed]
- Gawlik-Dziki, U.; Jeżyna, M.; Świeca, M.; Dziki, D.; Baraniak, B.; Czyż, J. Effect of bioaccessibility of phenolic compounds on in vitro anticancer activity of broccoli sprouts. Food Res. Int. 2012, 49, 469–476. [Google Scholar] [CrossRef]
- Hwang, J.H.; Lim, S.B. Antioxidant and anticancer activities of broccoli by-products from different cultivars and maturity stages at harvest. Prev. Nutr. Food Sci. 2015, 20, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Olsen, H.; Grimmer, S.; Aaby, K.; Saha, S.; Borge, G.I.A. Antiproliferative Effects of Fresh and Thermal Processed Green and Red Cultivars of Curly Kale (Brassica oleracea L. convar. acephala var. sabellica). J. Agric. Food Chem. 2012, 60, 7375–7383. [Google Scholar] [CrossRef]
- Michalak, M.; Szwajgier, D.; Paduch, R.; Kukula-Koch, W.; Waśko, A.; Polak-Berecka, M. Fermented curly kale as a new source of gentisic and salicylic acids with antitumor potential. J. Func. Foods 2020, 67, 103866. [Google Scholar] [CrossRef]
- Nazeri, M.; Nemati, H.; Khazaei, M. Nrf2 antioxidant pathway and apoptosis induction and inhibition of NF-κB-mediated inflammatory response in human prostate cancer PC3 cells by Brassica oleracea var. acephala: An in vitro study. Mol. Biol. Rep. 2022, 49, 7251–7261. [Google Scholar] [CrossRef]
- Furdak, P.; Pieńkowska, N.; Bartosz, G.; Sadowska-Bartosz, I. Extracts of Common Vegetables Inhibit the Growth of Ovary Cancer Cells. Foods 2022, 11, 2518. [Google Scholar] [CrossRef]
- Eroğlu, C.; Avcı, E.; Vural, H.; Kurar, E. Anticancer mechanism of Sinapic acid in PC-3 and LNCaP human prostate cancer cell lines. Gene 2018, 671, 127–134. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, H.; Tan, P.; Huang, M.; Shi, H.; Sun, B.; Cheng, Y.; Li, T.; Mou, Z.; Li, Q.; et al. Sinapic acid inhibits pancreatic cancer proliferation, migration, and invasion via downregulation of the AKT/Gsk-3β signal pathway. Drug Dev. Res. 2022, 83, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yu, H.; Guo, W.; Kong, Y.; Gu, L.; Li, Q.; Yang, S.; Zhang, Y.; Wang, Y. The anticancer effects of ferulic acid is associated with induction of cell cycle arrest and autophagy in cervical cancer cells. Cancer Cell Int. 2018, 18, 102. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Kumar, A.; Ramniwas, S.; Coudhary, R.; Aggarwal, D.; Kumar, M.; Sharma, U.; Parashar, N.C.; Haque, S.; Sak, K. Ferulic Acid: A Natural Phenol That Inhibits Neoplastic Events through Modulation of Oncogenic Signaling. Molecules 2022, 27, 7653. [Google Scholar] [CrossRef] [PubMed]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and Cancer Chemoprevention. Evid. Based Complement. Altern. Med. 2011, 2011, 591356. [Google Scholar] [CrossRef] [Green Version]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Mandrich, L.; Caputo, E. Brassicaceae-Derived Anticancer Agents: Towards a Green Approach to Beat Cancer. Nutrients 2020, 12, 868. [Google Scholar] [CrossRef] [Green Version]
- Pavlović, I.; Mlinarić, S.; Tarkowská, D.; Oklestková, J.; Novák, O.; Lepeduš, H.; Bok, V.V.; Brkanac, S.R.; Strnad, M.; Salopek-Sondi, B. Early Brassica crops responses to salinity stress: A Comparative Analysis between Chinese cabbage, White cabbage and Kale. Front. Plant Sci. 2019, 10, 450. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic–phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Bartoszek, A.; Wolska, L.; Drzewiecki, J.; Gorinstein, S.; Namiésnik, J. Partial characterization of white cabbages (Brassica oleracea var. capitata f. alba) from different regions by glucosinolates, bioactive compounds, total antioxidant activities and proteins. LWT Food Sci. Technol. 2008, 41, 1–9. [Google Scholar] [CrossRef]
- European Pharmacopoeia, 4th ed.; Council of Europe: Strasbourg, France, 2004; pp. 2377–2378.
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Meth. Enzym. 1999, 299, 15–27. [Google Scholar] [PubMed]
- Ferenac, M.; Polančec, D.; Huzak, M.; Pereira-Smith, O.M.; Rubelj, I. Early-Senescing Human Skin Fibroblasts Do Not Demonstrate Accelerated Telomere Shortening. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2005, 60, 820–829. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds (µg g−1 DW) | Kale | Wild Cabbage |
|---|---|---|
| cis, trans-Sinapic acid (SiA) | 2211.19 ± 61.74 | 2809.93 ± 387.56 * |
| trans-Ferulic acid (FA) | 1247.97 ± 13.74 | 1204.123 ± 148.18 |
| Salicylic acid (SA) | 2.95 ± 0.24 | 2.82 ± 0.34 |
| Kaempferol (KAE) | 0.42 ± 0.10 | 0.24 ± 0.07 * |
| Quercetin (QUE) | 0.20 ± 0.02 | 0.17 ± 0.01 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lučić, D.; Pavlović, I.; Brkljačić, L.; Bogdanović, S.; Farkaš, V.; Cedilak, A.; Nanić, L.; Rubelj, I.; Salopek-Sondi, B. Antioxidant and Antiproliferative Activities of Kale (Brassica oleracea L. Var. acephala DC.) and Wild Cabbage (Brassica incana Ten.) Polyphenolic Extracts. Molecules 2023, 28, 1840. https://doi.org/10.3390/molecules28041840
Lučić D, Pavlović I, Brkljačić L, Bogdanović S, Farkaš V, Cedilak A, Nanić L, Rubelj I, Salopek-Sondi B. Antioxidant and Antiproliferative Activities of Kale (Brassica oleracea L. Var. acephala DC.) and Wild Cabbage (Brassica incana Ten.) Polyphenolic Extracts. Molecules. 2023; 28(4):1840. https://doi.org/10.3390/molecules28041840
Chicago/Turabian StyleLučić, Dario, Iva Pavlović, Lidija Brkljačić, Sandro Bogdanović, Vladimir Farkaš, Andrea Cedilak, Lucia Nanić, Ivica Rubelj, and Branka Salopek-Sondi. 2023. "Antioxidant and Antiproliferative Activities of Kale (Brassica oleracea L. Var. acephala DC.) and Wild Cabbage (Brassica incana Ten.) Polyphenolic Extracts" Molecules 28, no. 4: 1840. https://doi.org/10.3390/molecules28041840