
Gastrodin Induces Ferroptosis of Glioma Cells via Upregulation of Homeobox D10

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
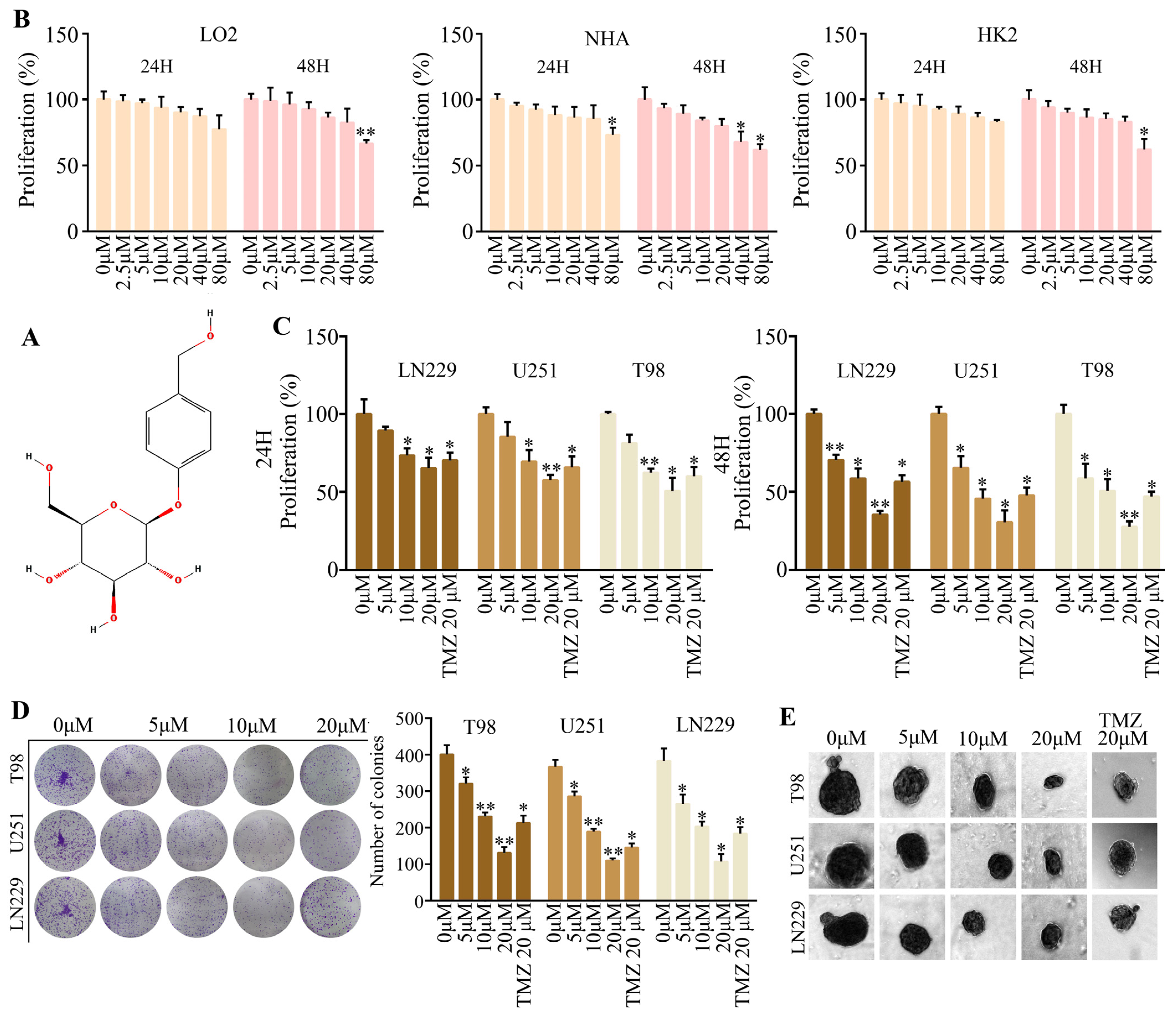
2.1. Gastrodine Exhibited Prominent Anti-Tumor Effects In Vitro
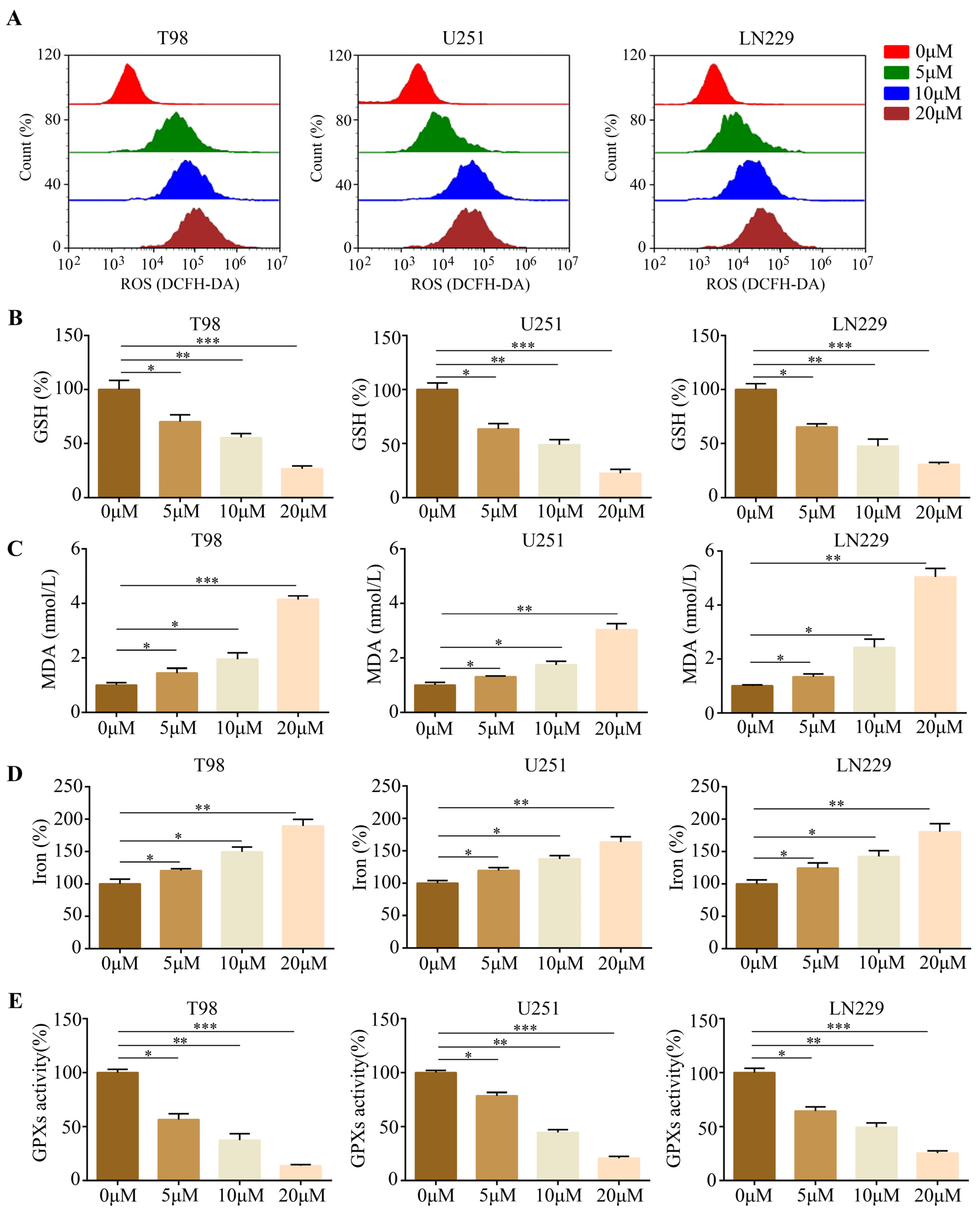
2.2. Gastrodin Induced Glioma Cell Ferroptosis In Vitro
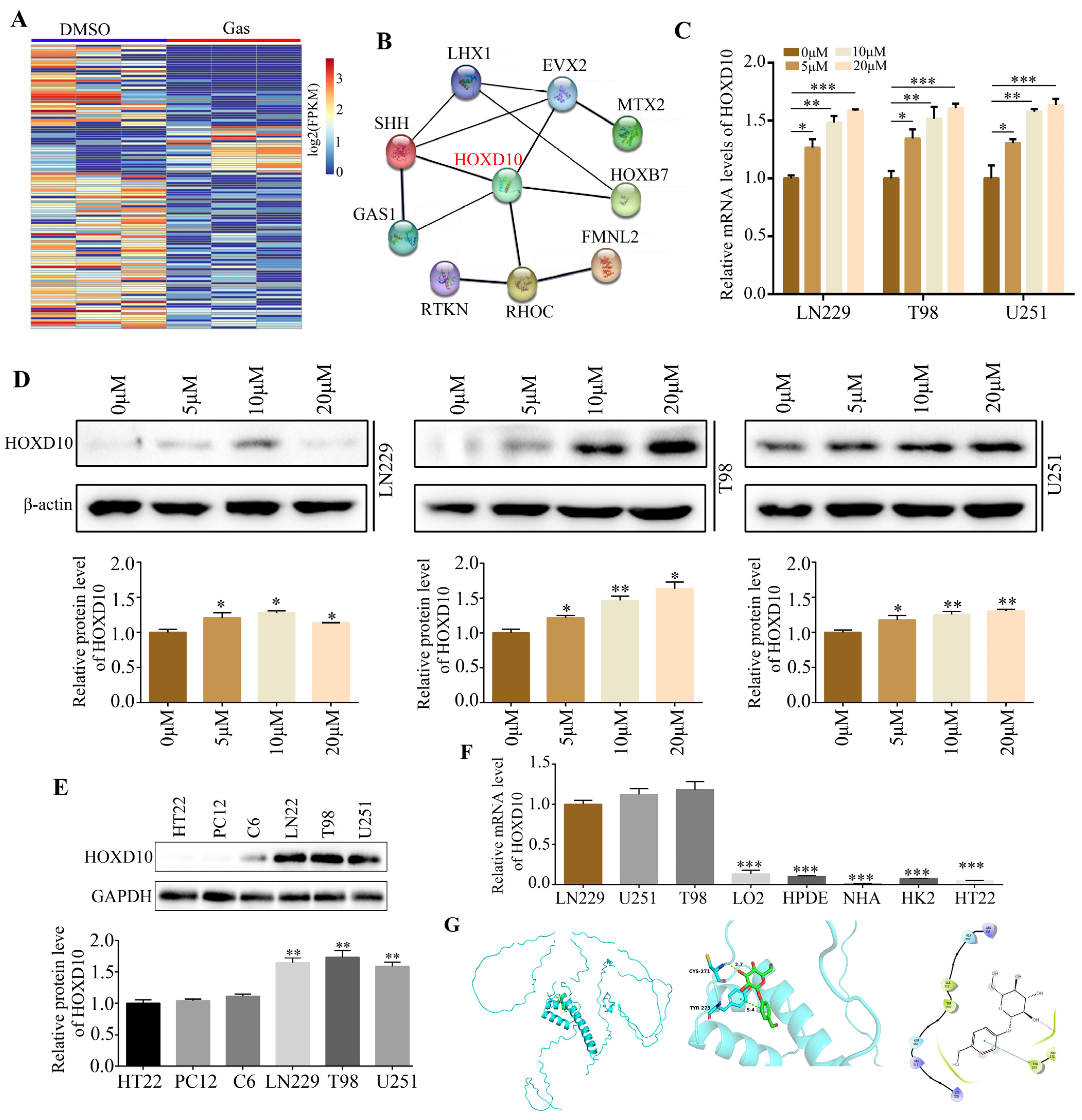
2.3. HOXD10 Was Identified as a Key Target of Gastrodin
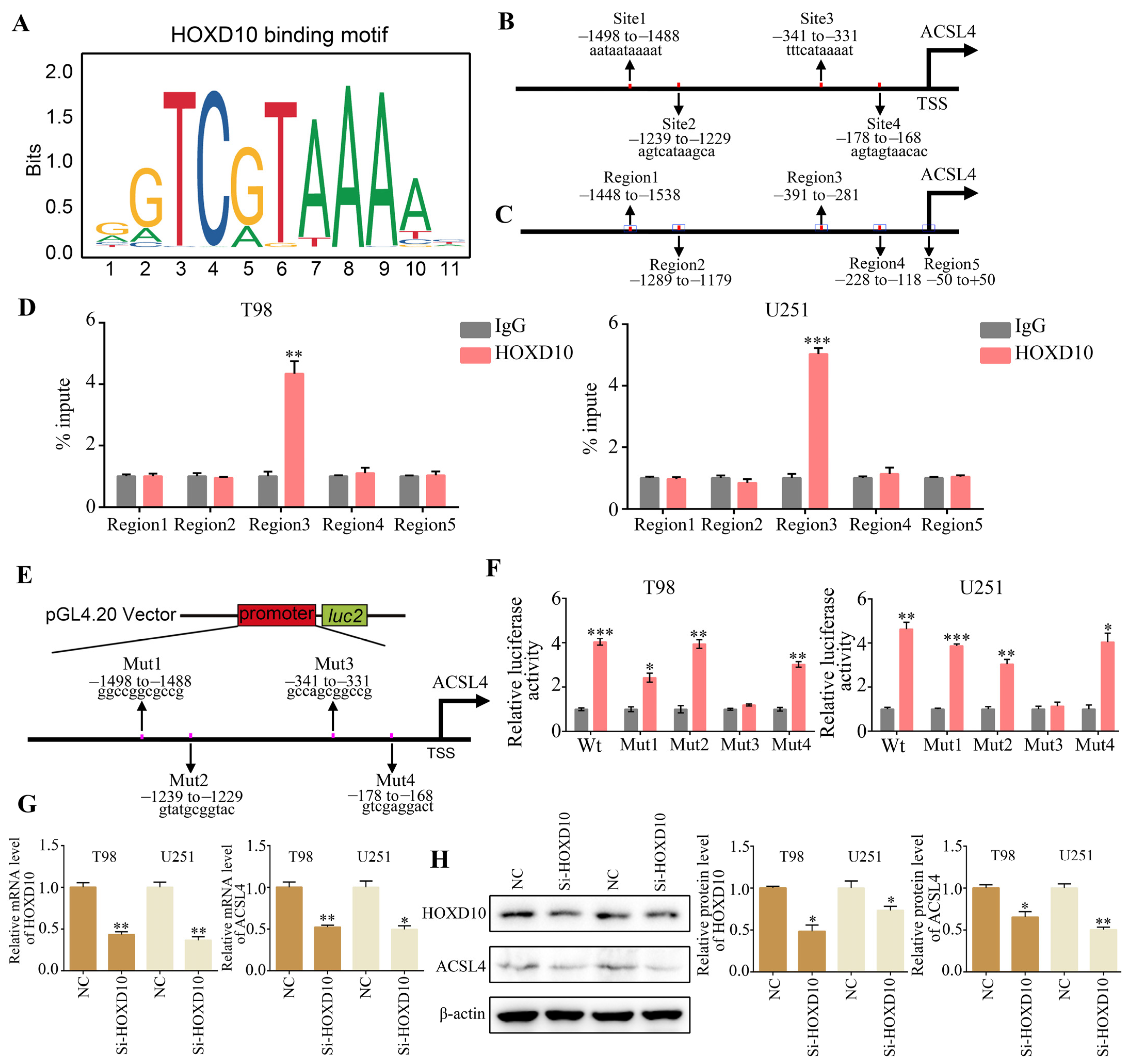
2.4. ACSL4 Is a Downstream Gene of HOXD10
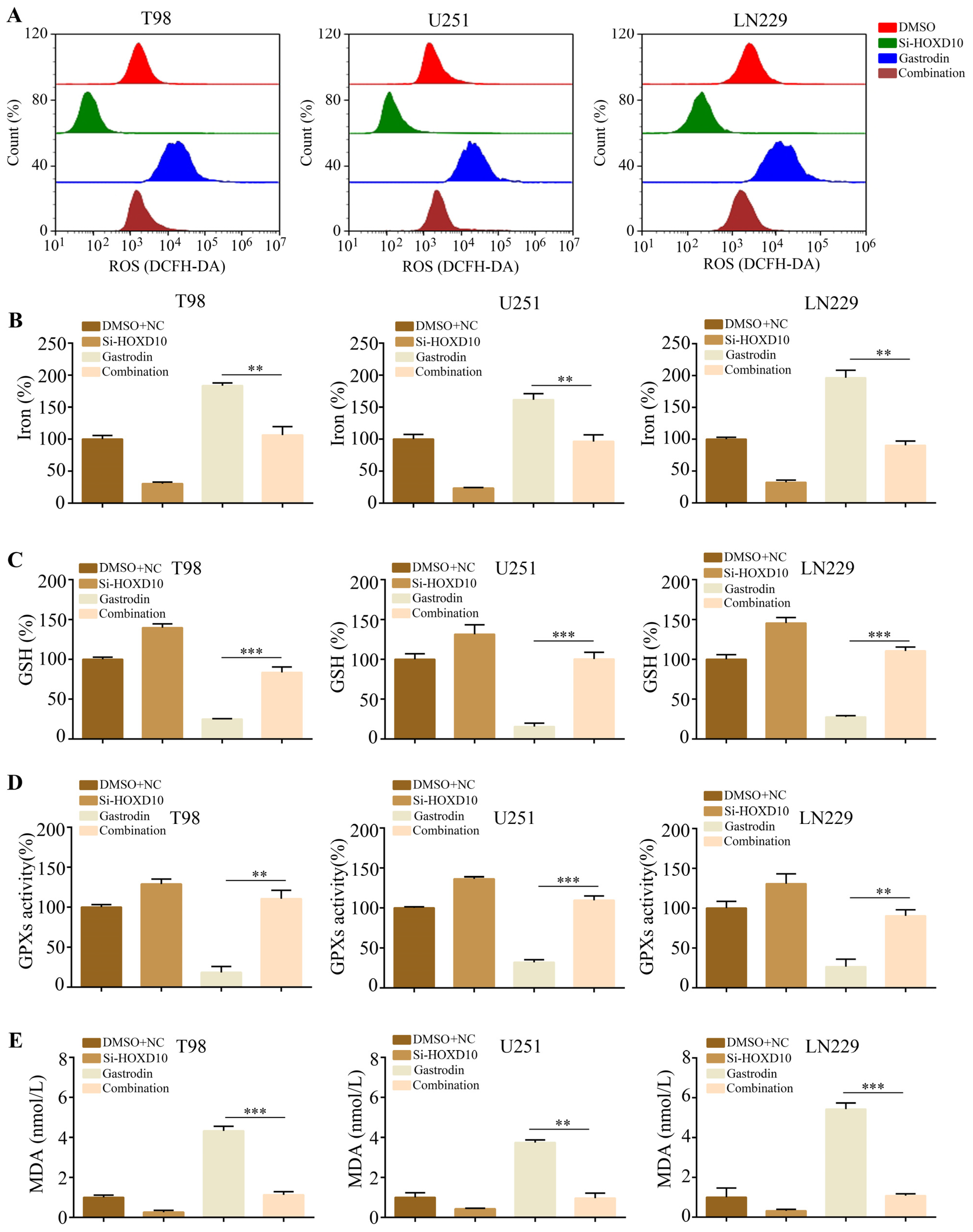
2.5. Silencing of HOXD10 Attenuated the Promotion of Ferroptosis by Gastrodin in Glioma Cells
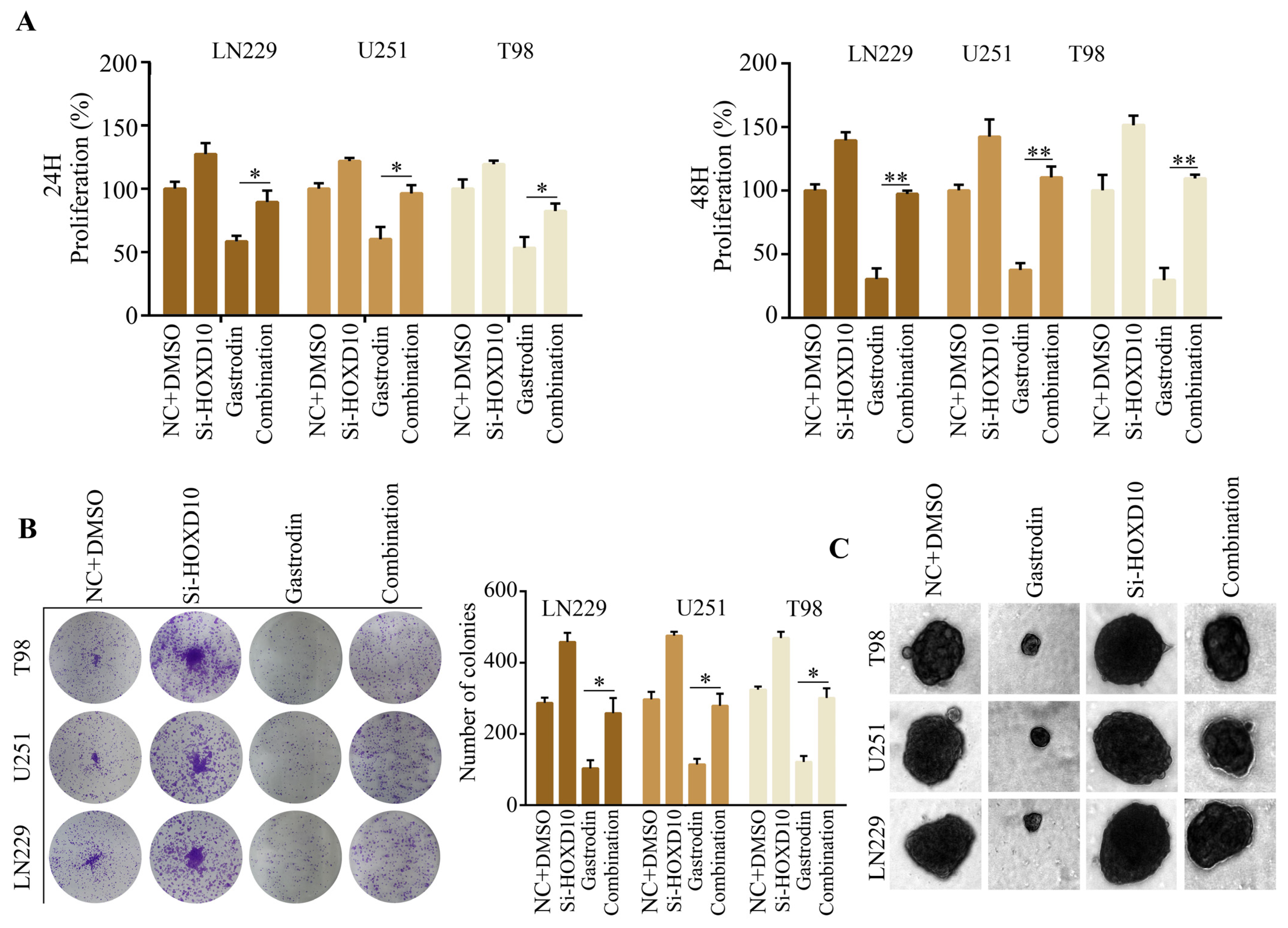
2.6. Silencing of HOXD10 Attenuated the Inhibitory Effect of Gastrodin on Glioma Cell Proliferation
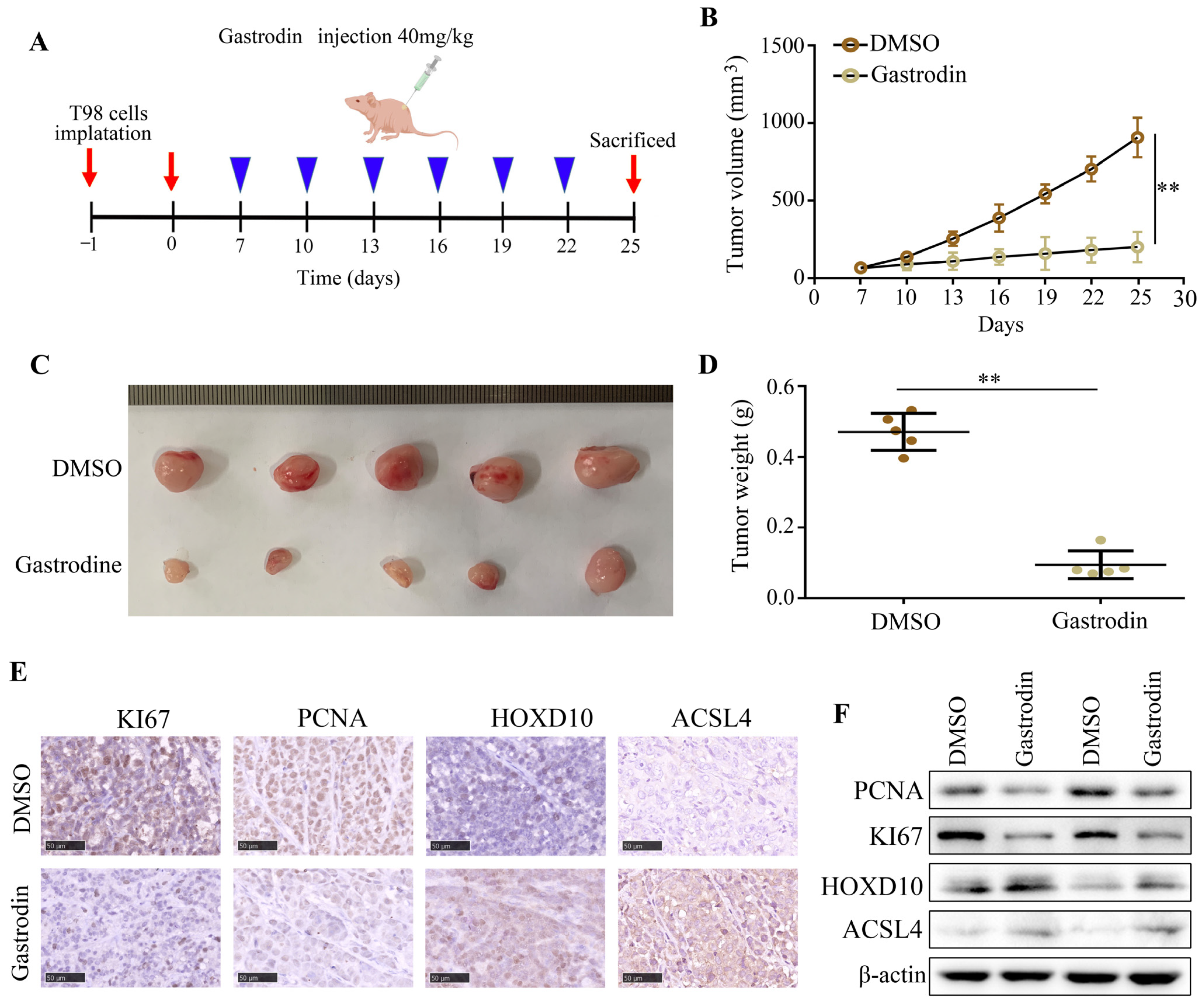
2.7. Gastrodin Suppressed the Proliferation of Glioma Cells In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. CCK-8 Assay
4.3. Spheroid Formation Experiment
4.4. Computer Molecular Docking
4.5. RNA Sequencing
4.6. Subcutaneous Tumorigenesis Experiments
4.7. Histology and Immunohistochemistry
4.8. Measurement of Cellular Ferroptosis Levels
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HOXD10 | Homeobox D10 |
| ROS | Reactive oxygen species |
| MDA | Malondialdehyde |
| GSH | Glutathione |
| ACSL4 | Acyl-CoA synthetase long-chain family member 4 |
| RT-qPCR | Real-time quantitative polymerase chain reaction |
References
- Berger, T.R.; Wen, P.Y.; Langorsini, M.; Chukwueke, U.N. World Health Organization 2021 Classification of Central Nervous System Tumors and Implications for Therapy for Adult-Type Gliomas: A Review. JAMA Oncol. 2022, 8, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wu, Z.; Zhang, H.; Zhang, N.; Wu, W.; Wang, Z.; Dai, Z.; Zhang, X.; Zhang, L.; Peng, Y.; et al. Glioma targeted therapy: Insight into future of molecular approaches. Mol. Cancer 2022, 21, 39. [Google Scholar] [CrossRef] [PubMed]
- Watowich, M.-B.; Gilbert, M.-R.; Larion, M. T cell exhaustion in malignant gliomas. Trends Cancer 2023, 9, 270–292. [Google Scholar] [CrossRef] [PubMed]
- Sauter, E.R. Cancer prevention and treatment using combination therapy with natural compounds. Expert. Rev. Clin. Pharmacol. 2020, 13, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Shafabakhsh, R.; Asemi, Z. Quercetin: A natural compound for ovarian cancer treatment. J. Ovarian Res. 2019, 12, 55. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Peng, M.; Meng, H.; Ma, H.; Cai, P.; Xu, Y.; Zhao, Q.; Si, G. A Review on Central Nervous System Effects of Gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar] [CrossRef]
- Jang, J.H.; Son, Y.; Kang, S.S.; Bae, C.S.; Kim, J.C.; Kim, S.H.; Shin, T.; Moon, C. Neuropharmacological Potential of Gastrodia elata Blume and Its Components. Evid. Based Complement. Alternat Med. 2015, 2015, 309261. [Google Scholar] [CrossRef]
- Liao, J.; Qing, X.; Deng, G.; Xiao, Y.; Fu, Y.; Han, S.; Li, X.; Gan, Y.; Li, W. Gastrodin destabilizes survivin and overcomes pemetrexed resistance. Cell Signal 2023, 110, 110851. [Google Scholar] [CrossRef]
- Shu, G.; Yang, T.; Wang, C.; Su, H.; Xiang, M. Gastrodin stimulates anticancer immune response and represses transplanted H22 hepatic ascitic tumor cell growth: Involvement of NF-κB signaling activation in CD4+ T cells. Toxicol. Appl. Pharmacol. 2013, 269, 270–279. [Google Scholar] [CrossRef]
- Cao, Y.M.; Gu, J.; Zhang, Y.S.; Wei, W.J.; Qu, N.; Wen, D.; Liao, T.; Shi, R.L.; Zhang, L.; Ji, Q.H.; et al. Aberrant hypermethylation of the HOXD10 gene in papillary thyroid cancer with BRAFV600E mutation. Oncol. Rep. 2018, 39, 338–348. [Google Scholar] [CrossRef]
- Yang, F.; Liu, D.; Deng, Y.; Wang, J.; Mei, S.; Ge, S.; Li, H.; Zhang, C.; Zhang, T. Frequent promoter methylation of HOXD10 in endometrial carcinoma and its pathological significance. Oncol. Lett. 2020, 19, 3602–3608. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, S.; Xue, M.; Zhong, J.; Wang, X.; Gan, L.; Lam, E.K.; Liu, X.; Zhang, J.; Zhou, T.; et al. Homeobox D10 gene, a candidate tumor suppressor, is downregulated through promoter hypermethylation and associated with gastric carcinogenesis. Mol. Med. 2012, 18, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yan, W.; Wang, Y.; Sun, G.; Luo, H.; Zhang, J.; Wang, X.; You, Y.; Yang, Z.; Liu, N.; et al. MicroRNA-10b induces glioma cell invasion by modulating MMP-14 and uPAR expression via HOXD10. Brain Res. 2011, 1389, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Xiong, Y.; Tu, Y.; Zhang, W.; Zhang, Q.; Nie, P.; Yan, X.; Liu, H.; Liu, R.; Xu, G. Elucidation of the Transport Mechanism of Puerarin and Gastrodin and Their Interaction on the Absorption in a Caco-2 Cell Monolayer Model. Molecules 2022, 27, 1230. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Cao, W.; Zeng, Z.; Zhang, Z.; Jin, B.; Tian, Q.; Wu, Y.; Zhang, T.; Li, D.; Hu, C. JUND/linc00976 promotes cholangiocarcinoma progression and metastasis, inhibits ferroptosis by regulating the miR-3202/GPX4 axis. Cell Death Dis. 2022, 13, 967. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qi, F.; Wang, Z.; Zhang, Z.; Pan, N.; Huai, L.; Qu, S.; Zhao, L. A review of traditional Chinese medicine for treatment of glioblastoma. Biosci. Trends. 2020, 13, 476–487. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, C.; Hou, J.; Long, H.; Wang, B.; Guo, D.; Lei, M.; Wu, W. Gastrodia elata Blume Polysaccharides: A Review of Their Acquisition, Analysis, Modification, and Pharmacological Activities. Molecules. 2019, 24, 2436. [Google Scholar] [CrossRef]
- Sun, X.; Jia, B.; Sun, J.; Lin, J.; Lu, B.; Duan, J.; Li, C.; Wang, Q.; Zhang, X.; Tan, M.; et al. Gastrodia elata Blume: A review of its mechanisms and functions on cardiovascular systems. Fitoterapia. 2023, 167, 105511. [Google Scholar] [CrossRef]
- Lai, Y.; Wang, R.; Li, W.; Zhu, H.; Fei, S.; Shi, H.; Lu, N.; Ung, C.O.L.; Hu, H.; Han, S. Clinical and economic analysis of Gastrodin injection for dizziness or vertigo: A retrospective cohort study based on electronic health records in China. Chin. Med. 2022, 17, 6. [Google Scholar] [CrossRef]
- Chen, J.; Gu, Y.T.; Xie, J.J.; Wu, C.C.; Xuan, J.; Guo, W.J.; Yan, Y.Z.; Chen, L.; Wu, Y.S.; Zhang, X.L.; et al. Gastrodin reduces IL-1β-induced apoptosis, inflammation, and matrix catabolism in osteoarthritis chondrocytes and attenuates rat cartilage degeneration in vivo. Biomed. Pharmacother. 2018, 97, 642–651. [Google Scholar] [CrossRef]
- Liang, W.Z.; Jan, C.R.; Hsu, S.S. Cytotoxic effects of gastrodin extracted from the rhizome of Gastrodia elata Blume in glioblastoma cells, but not in normal astrocytes, via the induction of oxidative stress-associated apoptosis that involved cell cycle arrest and p53 activation. Food Chem. Toxicol. 2017, 107, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Cheng, H.; Su, J.; Wang, X.; Wang, Q.; Chu, J.; Li, Q. Gastrodin protects against glutamate-induced ferroptosis in HT-22 cells through Nrf2/HO-1 signaling pathway. Toxicol. In Vitro. 2020, 62, 104715. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, E.; Yang, H.; Chen, Y.; Tao, L.; Xu, Y.; Chen, T.; Shen, X. Gastrodin Ameliorates Cognitive Dysfunction in Vascular Dementia Rats by Suppressing Ferroptosis via the Regulation of the Nrf2/Keap1-GPx4 Signaling Pathway. Molecules 2022, 227, 6311. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Chu, J.; Chen, H.; Cheng, H.; Su, J.; Wang, X.; Cao, Y.; Tian, S.; Li, Q. Gastrodin Inhibits H2O2-Induced Ferroptosis through Its Antioxidative Effect in Rat Glioma Cell Line C6. Biol. Pharm. Bull. 2020, 43, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, U.S.; Adiga, D.; Kabekkodu, S.P.; Hunter, K.D.; Radhakrishnan, R. Molecular implications of HOX genes targeting multiple signaling pathways in cancer. Cell Biol. Toxicol. 2022, 38, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.; Hunter, K.; Pandha, H.S. Downstream of the HOX genes: Explaining conflicting tumour suppressor and oncogenic functions in cancer. Int. J. Cancer. 2022, 150, 1919–1932. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sukumar, S. HOX genes: Emerging stars in cancer. Cancer Biol. Ther. 2003, 2, 524–525. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Song, G.; Zhang, X.; Gao, S.; Liu, H. HOX cluster-embedded antisense long non-coding RNAs in lung cancer. Cancer Lett. 2019, 450, 14–21. [Google Scholar] [CrossRef]
- Nakashoji, A.; Hayashida, T.; Yamaguchi, S.; Kawai, Y.; Kikuchi, M.; Yokoe, T.; Nagayama, A.; Seki, T.; Takahashi, M.; Kitagawa, Y. Comprehensive analysis of the homeobox family genes in breast cancer demonstrates their similar roles in cancer and development. Breast Cancer Res. Treat. 2021, 186, 353–361. [Google Scholar] [CrossRef]
- Gonçalves, C.S.; Leboiteux, E.; Arnaud, P.; Costa, B.M. HOX gene cluster (de)regulation in brain: From neurodevelopment to malignant glial tumours. Cell Mol. Life Sci. 2020, 77, 3797–3821. [Google Scholar] [CrossRef]
- Hakami, F.; Darda, L.; Stafford, P.; Woll, P.; Lambert, D.W.; Hunter, K.D. The roles of HOXD10 in the development and progression of head and neck squamous cell carcinoma (HNSCC). Br. J. Cancer 2014, 111, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.F.S.D.; Xavier, F.C.A.; Esteves, C.D.; Nascimento, R.B.; Nobile, J.S.; Severino, P.; deCicco, R.; Toporcov, T.N.; Tajara, E.H.; Nunes, F.D. Homeobox gene amplification and methylation in oral squamous cell carcinoma. Arch. Oral. Biol. 2021, 129, 105195. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y. The interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, Y.; Zhao, X.; Shao, L.; Liu, G.; Sun, C.; Xu, R.; Zhang, Z. ACSL4 exacerbates ischemic stroke by promoting ferroptosis-induced brain injury and neuroinflammation. Brain Behav. Immun. 2021, 93, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Hu, B.X.; Li, Z.L.; Du, T.; Shan, J.L.; Ye, Z.P.; Peng, X.D.; Li, X.; Huang, Y.; Zhu, X.Y.; et al. PKCβII phosphorylates ACSL4 to amplify lipid peroxidation to induce ferroptosis. Nat. Cell Biol. 2022, 24, 88–98. [Google Scholar] [CrossRef]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Killion, E.A.; Reeves, A.R.; ElAzzouny, M.A.; Yan, Q.W.; Surujon, D.; Griffin, J.D.; Bowman, T.A.; Wang, C.; Matthan, N.R.; Klett, E.L.; et al. A role for long-chain acyl-CoA synthetase-4 (ACSL4) in diet-induced phospholipid remodeling and obesity-associated adipocyte dysfunction. Mol. Metab. 2018, 9, 43–56. [Google Scholar] [CrossRef]
- Yuan, H.; Li, X.; Zhang, X.; Kang, R.; Tang, D. Identification of ACSL4 as a biomarker and contributor of ferroptosis. Biochem. Biophys. Res. Commun. 2016, 478, 1338–1343. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, W.; Lan, J.; Zeng, Z.; Yu, W.; Lei, S. Gastrodin Induces Ferroptosis of Glioma Cells via Upregulation of Homeobox D10. Molecules 2023, 28, 8062. https://doi.org/10.3390/molecules28248062
Cao W, Lan J, Zeng Z, Yu W, Lei S. Gastrodin Induces Ferroptosis of Glioma Cells via Upregulation of Homeobox D10. Molecules. 2023; 28(24):8062. https://doi.org/10.3390/molecules28248062
Chicago/Turabian StyleCao, Wenpeng, Jinzhi Lan, Zhirui Zeng, Wenfeng Yu, and Shan Lei. 2023. "Gastrodin Induces Ferroptosis of Glioma Cells via Upregulation of Homeobox D10" Molecules 28, no. 24: 8062. https://doi.org/10.3390/molecules28248062