


Occurrence of Luteolin in the Greek Flora, Isolation of Luteolin and Its Action for the Treatment of Periodontal Diseases

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Determination of Luteolin Content in Plants of the Greek Flora
3. Effect of Crop Management on Luteolin Content
4. Methods of Luteolin Extraction from Different Plant Species
4.1. Conventional Extraction Methods
4.2. Modern Extraction Methods
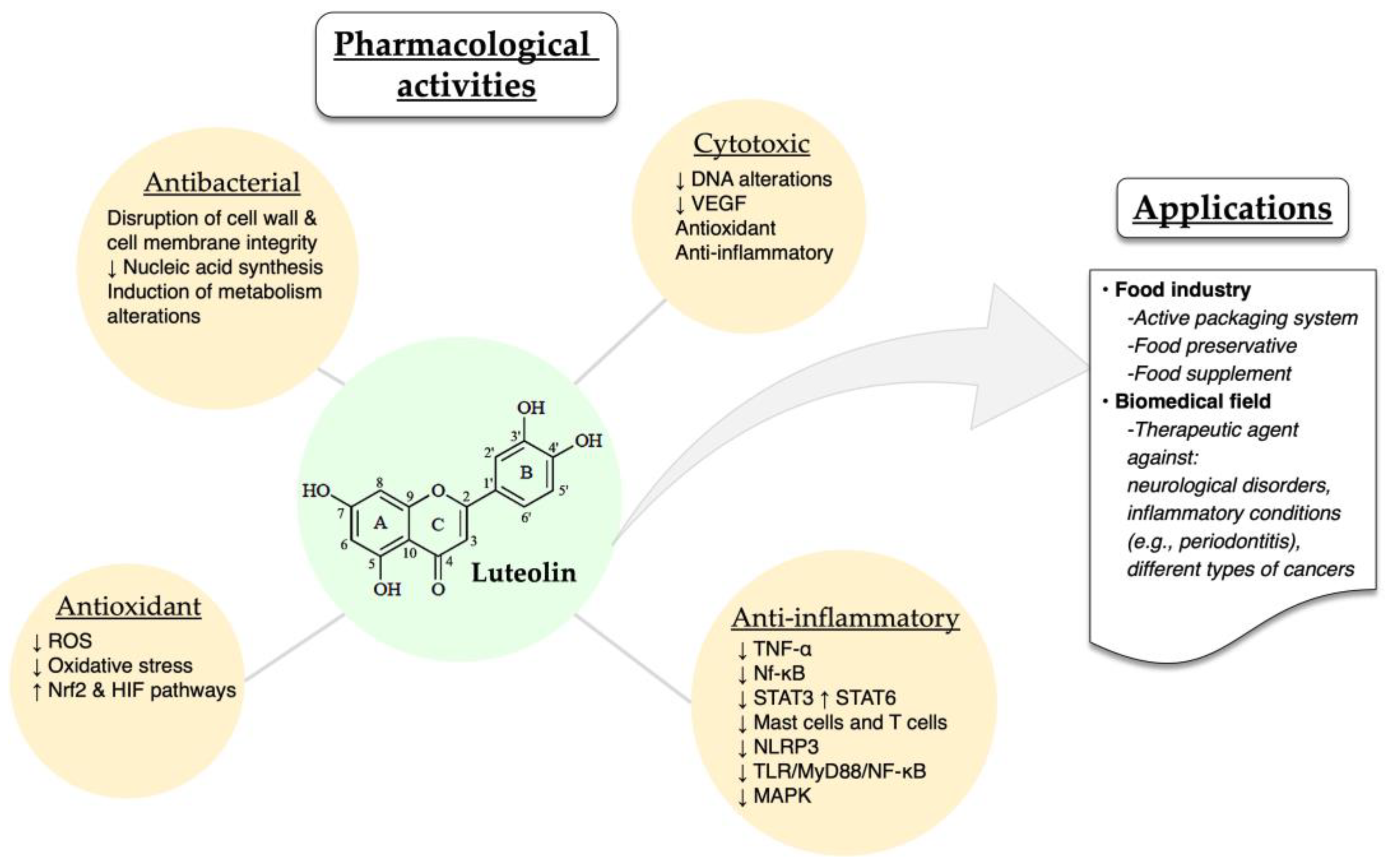
5. Pharmacological Activities and Applications of Luteolin
6. The Effect of Luteolin in the Prevention and Treatment of Periodontal Disease
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- González-Sarrías, A.; Tomás-Barberán, F.A.; García-Villalba, R. Structural Diversity of Polyphenols and Distribution in Foods. Diet. Polyphen. Their Metab. Health Eff. 2020, 1–29. [Google Scholar]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food sources, bioavailability, metabolism, and bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef]
- Gurung, R.B.; Pandey, R.P.; Sohng, J.K. Apigenin and Naringenin: Natural Sources, Pharmacology and Role in Cancer Prevention. Chapter: Role of Apigenin in Cancer Prevention. Nova Sci. 2015. [Google Scholar]
- Birt, D.F.; Hendrich, S.; Wang, W. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther. 2001, 90, 157–177. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potentials for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- Chan, T.S.; Galati, G.; Pannala, A.S.; Rice-Evans, C.; O’Brien, P.J. Simultaneous detection of the antioxidant and pro-oxidant activity of dietary polyphenolics in a peroxidase system. Free. Radic. Res. 2003, 37, 787–794. [Google Scholar] [CrossRef]
- Hempel, J.; Pforte, H.; Raab, B.; Engst, W.; Bohm, H.; Jacobasch, G. Flavonols and flavones of parsley cell suspension culture change the antioxidative capacity of plasma in rats. Nahrung 1999, 43, 201–204. [Google Scholar] [CrossRef]
- Bogers, R.J.; Craker, L.E.; Lange, D. Medicinal and Aromatic Plants: Agricultural, Commercial, Ecological, Legal, Pharmacological and Social Aspects (Wageningen UR Frontis Series); Springer: Berlin, Germany, 2006; Volume 17, 309p. [Google Scholar]
- Giannoulis, K.; Evangelopoulos, V.; Gougoulias, N.; Wogiatzi, E. Could bio-stimulators affect flower, essential oil yield, and its composition in organic lavender (Lavandula angustifolia) cultivation? Ind. Crops Prod. 2020, 154, 112611. [Google Scholar] [CrossRef]
- Leaman, D.J. The International Standard for Sustainable Wild Collection of Medicinal and Aromatic Plants (ISSC-MAP): Elements of ISSC-MAP Resource Assessment Guidance Relevant to Cites NDF; International Expert Workshop on CITES Non-Detriment Findings Perennial Plant Working Group (Ornamentals, Medicinal and Aromatic Plants Cancun, Mexico): Cancun, Mexico, 2008. [Google Scholar]
- Negi, P.S. Plant extracts for the control of bacterial growth: Efficacy, stability and safety issues for food application. Int. J. Food Microbiol. 2012, 156, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Edeoga, H.O.; Okwu, D.E.; Mbaebie, B.O. Phytochemical constituents of some Nigerian medicinal plants. Afr. J. Biotechnol. 2005, 4, 685–688. [Google Scholar] [CrossRef]
- Kadri, A.; Zarai, Z.; Chobba, I.B.; Bekir, A.; Gharsallah, N.; Damak, M.; Gdoura, R. Chemical constituents and antioxidant properties of Rosmarinus officinalis L. essential oil cultivated from South-Western Tunisia. J. Med. Plants Res. 2011, 5, 5999–6004. [Google Scholar]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Aromatic plants as a source of bioactive compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef]
- Nur, A.; Mi-Yeon, K.; Jae, Y.C. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar]
- Shimoi, K.; Okada, H.; Furugori, M.; Goda, T.; Takase, S.; Suzuki, M.; Hara, Y.; Yamamoto, H.; Kinae, N. Intestinal absorption of luteolin and luteolin 7-O-beta-glucoside in rats and humans. FEBS Lett. 1998, 438, 220–224. [Google Scholar] [CrossRef]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Kostić, Μ.; Kitić, D.; Petrović, M.B.; Jevtović-Stoimenov, T.; Jović, M.; Petrović, A.; Živanović, S. Anti-inflammatory effect of the Salvia sclarea L. ethanolic extract on lipopolysaccharide-induced periodontitis in rats. J. Ethnopharmacol. 2017, 199, 52–59. [Google Scholar] [CrossRef]
- Casili, G.; Ardizzone, A.; Lanza, M.; Gugliandolo, E.; Portelli, M.; Militi, A.; Cuzzocrea, S.; Esposito, E.; Paterniti, I. Treatment with Luteolin Improves Lipopolysaccharide-Induced Periodontal Diseases in Rats. Biomedicines 2020, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Dretcanu, G.; Stirbu, I.; Leoplold, N.; Cruceriu, D.; Danciu, C.; Stanila, A.; Farcas, A.; Borda, I.M.; Iuhas, C.; Diaconeasa, Z. Chemical structure, sources and role of bioactive flavonoids in cancer prevention: A Review. Plants 2022, 11, 1117. [Google Scholar] [CrossRef]
- Chimona, C.; Karioti, A.; Skaltsa, H.; Rhizopoulou, S. Occurrence of secondary metabolites in tepals of Asphodelus ramosus L. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 31–34. [Google Scholar]
- Barda, C.; Ciric, A.; Soković, M.; Tsoukalas, M.; Skaltsa, H. Phytochemical investigation of Crepis incana Sm. (Asteraceae) endemic to southern Greece. Biochem. Syst. Ecol. 2018, 80, 59–62. [Google Scholar] [CrossRef]
- Proestos, C.; Boziaris, I.S.; Nychas, G.J.E.; Komaitis, M. Analysis of flavonoids and phenolic acids in Greek aromatic plants: Investigation of their antioxidant capacity and antimicrobial activity. Food Chem. 2006, 95, 664–671. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Pereira, C.; Barros, L.; Ferreira, I.C.F.R. Leaf parts from Greek artichoke genotypes as a good source of bioactive compounds and antioxidants. Food Funct. 2017, 8, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Chinou, I.; Harvala, C. Polyphenolic Constituents from the Leaves of two Cynara Species Growing in Greece. Planta Medica 1997, 63, 469–470. [Google Scholar] [CrossRef]
- Kalogiouri, N.P.; Samanidou, V.F. A validated ultrasound-assisted extraction coupled with spe-hplc-dad for the determination of flavonoids in by-products of plant origin: An application study for the valorization of the walnut septum membrane. Molecules 2021, 26, 6418. [Google Scholar] [CrossRef]
- Stefanova, G.; Girova, T.; Gochev, V.; Stoyanova, M.; Petkova, Z.; Stoyanova, A.; Zheljazkov, V.D. Comparative study on the chemical composition of laurel (Laurus nobilis L.) leaves from Greece and Georgia and the antibacterial activity of their essential oil. Heliyon 2020, 6, e05491. [Google Scholar] [CrossRef]
- Tsakni, A.; Chatzilazarou, A.; Tsakali, E.; Tsantes, A.G.; Van Impe, J.; Houhoula, D. Identification of Bioactive Compounds in Plant Extracts of Greek Flora and Their Antimicrobial and Antioxidant Activity. Separations 2023, 10, 373. [Google Scholar] [CrossRef]
- Miceli, N.; Taviano, M.F.; Giuffrida, D.; Trovato, A.; Tzakou, O.; Galati, E.M. Anti-inflammatory activity of extract and fractions from Nepeta sibthorpii Bentham. J. Ethnopharmacol. 2005, 97, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.B.; Patras, A.; Barry-Ryan, C.; Martin-Diana, A.B.; Brunton, N. Application of principal component and hierarchical cluster analysis to classify different spices based on in vitro antioxidant activity and individual polyphenolic antioxidant compounds. J. Funct. Foods 2011, 3, 179–189. [Google Scholar] [CrossRef]
- Agiomyrgianaki, A.; Petrakis, P.V.; Dais, P. Influence of harvest year, cultivar and geographical origin on Greek extra virgin olive oils composition: A study by NMR spectroscopy and biometric analysis. Food Chem. 2012, 135, 2561–2568. [Google Scholar] [CrossRef] [PubMed]
- Blekas, G.; Vassilakis, C.; Harizanis, C.; Tsimidou, M.; Boskou, D.G. Biophenols in table olives. J. Agric. Food Chem. 2002, 50, 3688–3692. [Google Scholar] [CrossRef] [PubMed]
- Christophoridou, S.; Dais, P. Detection and quantification of phenolic compounds in olive oil by high resolution 1H nuclear magnetic resonance spectroscopy. Anal. Chim. Acta 2009, 633, 283–292. [Google Scholar] [CrossRef]
- Kiritsakis, K.; Kontominas, M.G.; Kontogiorgis, C.; Hadjipavlou-Litina, D.; Moustakas, A.; Kiritsakis, A. Composition and antioxidant activity of olive leaf extracts from Greek olive cultivars. JAOCS J. Am. Oil Chem. Soc. 2010, 87, 369–376. [Google Scholar] [CrossRef]
- Kotsiou, K.; Tasioula-Margari, M. Monitoring the phenolic compounds of Greek extra-virgin olive oils during storage. Food Chem. 2016, 200, 255–262. [Google Scholar] [CrossRef]
- Mitsopoulos, G.; Papageorgiou, V.; Komaitis, M.; Hagidimitriou, M. Phenolic Profile of leaves and drupes in major Greek olive varieties. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 162–166. [Google Scholar] [CrossRef]
- Salis, C.; Papadakis, I.E.; Hagidimitriou, M. Identification and Quantification of Phenolic Compounds in Fresh and Processed Table Olives of Cv. ‘kalamata’. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 1–13. [Google Scholar] [CrossRef]
- Tsantili, E. Quality attributes and their relations in fresh black ripe “Kalamon” olives (Olea europaea L.) for table use—Phenolic compounds and total antioxidant capacity. Int. J. Food Sci. Technol. 2014, 49, 657–665. [Google Scholar] [CrossRef]
- Exarchou, V.; Nenadis, N.; Tsimidou, M.; Gerothanassis, I.P.; Troganis, A.; Boskou, D. Antioxidant activities and phenolic composition of extracts from Greek oregano, Greek sage, and summer savory. J. Agric. Food Chem. 2002, 50, 5294–5299. [Google Scholar] [CrossRef] [PubMed]
- Klontza, V.; Graikou, K.; Cheilari, A.; Kasapis, V.; Ganos, C.; Aligiannis, N.; Chinou, I. Phytochemical Study on Seeds of Paeonia clusii subsp. rhodia—Antioxidant and Anti-Tyrosinase Properties. Int. J. Mol. Sci. 2023, 24, 4935. [Google Scholar] [CrossRef] [PubMed]
- Boutsika, A.; Sarrou, E.; Cook, C.M.; Mellidou, I.; Avramidou, E.; Angeli, A.; Martens, S.; Ralli, P.; Letsiou, S.; Selini, A.; et al. Evaluation of parsley (Petroselinum crispum) germplasm diversity from the Greek Gene Bank using morphological, molecular and metabolic markers. Ind. Crops Prod. 2021, 170, 113767. [Google Scholar] [CrossRef]
- Proestos, C.; Chorianopoulos, N.; Nychas, G.J.E.; Komaitis, M. RP-HPLC analysis of the phenolic compounds of plant extracts. Investigation of their antioxidant capacity and antimicrobial activity. J. Agric. Food Chem. 2005, 53, 1190–1195. [Google Scholar] [CrossRef]
- Bampouli, A.; Kyriakopoulou, K.; Papaefstathiou, G.; Louli, V.; Aligiannis, N.; Magoulas, K.; Krokida, M. Evaluation of total antioxidant potential of Pistacia lentiscus var. chia leaves extracts using UHPLC–HRMS. J. Food Eng 2015, 167, 25–31. [Google Scholar] [CrossRef]
- Grigorakis, S.; Halahlah, A.; Makris, D.P. Hydroglycerolic solvent and ultrasonication pretreatment: A green blend for high-efficiency extraction of Salvia fruticosa polyphenols. Sustainability 2020, 12, 4840. [Google Scholar] [CrossRef]
- Cvetkovikj, I.; Stefkov, G.; Acevska, J.; Stanoeva, J.P.; Karapandzova, M.; Stefova, M.; Dimitrovska, A.; Kulevanova, S. Polyphenolic characterization and chromatographic methods for fast assessment of culinary Salvia species from South East Europe. J. Chromatogr. A 2013, 1282, 38–45. [Google Scholar] [CrossRef]
- Choulitoudi, E.; Xristou, M.; Tsimogiannis, D.; Oreopoulou, V. The effect of temperature on the phenolic content and oxidative stability of o/w emulsions enriched with natural extracts from Satureja thymbra. Food Chem. 2021, 349, 129206. [Google Scholar] [CrossRef]
- Proestos, C.; Sereli, D.; Komaitis, M. Determination of phenolic compounds in aromatic plants by RP-HPLC and GC-MS. Food Chem. 2006, 95, 44–52. [Google Scholar] [CrossRef]
- Tavarini, S.; Sgherri, C.; Ranieri, A.M.; Angelini, L.G. Effect of nitrogen fertilization and harvest time on steviol glycosides, flavonoid composition and antioxidant properties in Stevia rebaudiana Bertoni. J. Agric. Food Chem. 2015, 31, 7041–7050. [Google Scholar] [CrossRef]
- Takahashi, A.; Ohnishi, T. The significance of the study about the biological effects of solar ultraviolet radiation using the exposed facility on the international space station. Biol. Sci. Space 2004, 18, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, A.; Caporali, S.; Di Daniele, N.; Rovella, V.; Cardillo, C.; Schinzari, F.; Minieri, M.; Pieri, M.; Candi, E.; Bernardini, S.; et al. Anti-Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. Int. J. Mol. Sci. 2021, 22, 1321. [Google Scholar] [CrossRef]
- Salata, A.; Nurzynska-Wierdak, R.; Kalisz, A.; Kunicki, E.; Ibáñez-Asensio, S.; Moreno-Ramón, H. Effects of organic cropping on phenolic compounds and antioxidant capacity of globe artichoke herbs. Agronomy 2022, 12, 192. [Google Scholar] [CrossRef]
- Ribes-Moya, A.M.; Adalid, A.M.; Raigon, M.D.; Hellin, P.; Fita, A.; Rodríguez-Burruezo, A. Variation in flavonoids in a collection of peppers (Capsicum sp.) under organic and conventional cultivation: Effect of the genotype, ripening stage, and growing system. J. Sci. Food Agric. 2020, 100, 2208–2223. [Google Scholar] [CrossRef] [PubMed]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Abreu, I.N.; Mazzafera, P. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef]
- Hodaei, Μ.; Rahimmalek, Μ.; Arzania, A.; Talebib, Μ. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crops Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Rajabbeigi, E.; Eichholz, I.; Beesk, N.; Ulrichs, C.; Kroh, L.W.; Rohn, S.; Huyskens-Keil, S. Interaction of drought stress and UV-B radiation—Impact on biomass production and flavonoid metabolism in lettuce (Lactuca sativa L.). J. Appl. Bot. Food Qual. 2013, 86, 190–197. [Google Scholar]
- Trivellini, A.; Lucchesini, M.; Maggini, R.; Mosadegh, H.; Villamarin, T.S.S.; Vernieri, P.; Mensuali-Sodi, A.; Pardossi, A. Lamiaceae phenols as multifaceted compounds: Bioactivity, industrial prospects and role of “positive-stress”. Ind. Crops Prod. 2016, 83, 241–254. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Shafeiee, M.; Ehsanzadeh, P. Physiological and biochemical mechanisms of salinity tolerance in several fennel genotypes: Existence of clearly-expressed genotypic variations. Ind. Crops Prod. 2019, 132, 311–318. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Ehsanzadeh, P.; Ghanadian, M. Salinity stress alters ion homeostasis, antioxidant activities and the production of rosmarinic acid, luteolin and apigenin in Dracocephalum kotschyi Boiss. Biologia 2020, 75, 2147–2158. [Google Scholar] [CrossRef]
- Agati, G.; Biricolti, S.; Guidi, L.; Ferrini, F.; Fini, A.; Tattini, M. The biosynthesis of flavonoids is enhanced similarly by UV radiation and root zone salinity in L. vulgare leaves. J. Plant Physiol. 2011, 168, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, S.B.; Aung, B.; Amyot, L.; Lalin, I.; Lachaal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 72. [Google Scholar] [CrossRef]
- El-Shafey, N.M.; AbdElgawad, H. Luteolin, a bioactive flavone compound extracted from Cichorium endivia L. subsp. divaricatum alleviates the harmful effect of salinity on maize. Acta Physiol. Plant. 2012, 34, 2165–2177. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Papageorgiou, V.; Mallouchos, A.; Komaitis, M. Investigation of the antioxidant behavior of air- and freeze-dried aromatic plant materials in relation to their phenolic content and vegetative cycle. J. Agric. Food Chem. 2008, 56, 5743–5752. [Google Scholar] [CrossRef]
- Skoula, M.; Grayer, R.J.; Kite, G.C.; Veitch, N.C. Exudate flavones and flavanones in Origanum species and their interspecific variation. Biochem. Syst. Ecol. 2008, 36, 646–654. [Google Scholar] [CrossRef]
- Sellami, I.H.; Maamouri, E.; Chahed, T.; Wannes, W.A.; Kchouk, M.E.; Marzouk, B. Effect of growth stage on the content and composition of the essential oil and phenolic fraction of sweet marjoran (Origanum majorana L.). Ind. Crops Prod. 2009, 30, 395–402. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Kempuraj, D.; Iliopoulou, B.P. Mast cells, T cells, and inhibition by luteolin: Implications for the pathogenesis and treatment of multiple sclerosis. Immune-Mediat. Dis. Theory Ther. 2007, 601, 423–430. [Google Scholar]
- Kempuraj, D.; Tagen, M.; Iliopoulou, B.P.; Clemons, A.; Vasiadi, M.; Boucher, W.; House, M.; Wolfberg, A.; Theoharides, T.C. Luteolin inhibits myelin basic protein-induced human mast cell activation and mast cell-dependent stimulation of Jurkat T cells. Br. J. Pharmacol. 2008, 155, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. Luteolin as a therapeutic option for multiple sclerosis. J. Neuroinflammation 2009, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Kritas, S.K.; Saggini, A.; Varvara, G.; Murmura, G.; Caraffa, A.; Antinolfi, P.; Toniato, E.; Pantalone, A.; Neri, G.; Frydas, S.; et al. Luteolin inhibits mast cell-mediated allergic inflammation. J. Biol. Regul. Homeost. Agents 2013, 27, 955–959. [Google Scholar]
- Lee, M.N.; Lee, Y.; Wu, D.; Pae, M. Luteolin inhibits NLRP3 inflammasome activation via blocking ASC oligomerization. J. Nutr. Biochem. 2021, 92, 108614. [Google Scholar] [CrossRef]
- Wang, S.; Cao, M.; Xu, S.; Shi, J.; Mao, X.; Yao, X.; Liu, C. Luteolin Alters Macrophage Polarization to Inhibit Inflammation. Inflammation 2020, 43, 95–108. [Google Scholar] [CrossRef]
- Shen, R.; Ma, L.; Zheng, Y. Anti-inflammatory effects of luteolin on acute gouty arthritis rats via TLR/MyD88/NF-kappaB pathway. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2020, 45, 115–122. [Google Scholar]
- Huang, W.C.; Liou, C.J.; Shen, S.C.; Hu, S.; Hsiao, C.Y.; Wu, S.J. Luteolin Attenuates IL-1beta-Induced THP-1 Adhesion to ARPE-19 Cells via Suppression of NF-kappaB and MAPK Pathways. Mediat. Inflamm. 2020, 2020, 9421340. [Google Scholar] [CrossRef]
- Li, B.; Du, P.; Du, Y.; Zhao, D.; Cai, Y.; Yang, Q.; Guo, Z. Luteolin alleviates inflammation and modulates gut microbiota in ulcerative colitis rats. Life Sci. 2021, 269, 119008. [Google Scholar] [CrossRef]
- Kou, J.J.; Shi, J.Z.; He, Y.Y.; Hao, J.J.; Zhang, H.Y.; Luo, D.M.; Song, J.K.; Yan, Y.; Xie, X.M.; Du, G.H.; et al. Luteolin alleviates cognitive impairment in Alzheimer’s disease mouse model via inhibiting endoplasmic reticulum stress-dependent neuroinflammation. Acta Pharmacol. Sin. 2022, 43, 840–849. [Google Scholar] [CrossRef]
- Liu, M.; Cheng, C.; Li, X.; Zhou, S.; Hua, J.; Huang, J.; Li, Y.; Yang, K.; Zhang, P.; Zhang, Y.; et al. Luteolin alleviates ochratoxin A induced oxidative stress by regulating Nrf2 and HIF-1alpha pathways in NRK-52E rat kidney cells. Food Chem. Toxicol. 2020, 141, 111436. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Y.; Zhang, Z.; Chen, M.; Zhang, D.; Tian, C.; Liu, M.; Jiang, G. The Antibacterial Activity and Mechanism of Action of Luteolin Against Trueperella pyogenes. Infect. Drug Resist. 2020, 13, 1697–1711. [Google Scholar] [CrossRef] [PubMed]
- Bangar, S.P.; Kajla, P.; Chaudhary, V.; Sharma, N.; Ozogul, F. Luteolin: A flavone with myriads of bioactivities and food applications. Food Biosci. 2023, 52, 102366. [Google Scholar]
- Bi, F.; Zhang, X.; Liu, J.; Yong, H.; Gao, L.; Liu, J. Development of antioxidant and antimicrobial packaging films based on chitosan, D-α-tocopheryl polyethylene glycol 1000 succinate and silicon dioxide nanoparticles. Food Packag. Shelf Life 2020, 24, 100503. [Google Scholar] [CrossRef]
- Mhalla, D.; Bouaziz, A.; Ennouri, K.; Chawech, R.; Smaoui, S.; Jarraya, R.; Tounsi, S.; Trigui, M. Antimicrobial activity and bioguided fractionation of Rumex tingitanus extracts for meat preservation. Meat Sci. 2017, 125, 22–29. [Google Scholar] [CrossRef]
- Huang, M.; Su, E.; Zheng, F.; Tan, C. Encapsulation of flavonoids in liposomal delivery systems: The case of quercetin, kaempferol and luteolin. Food Funct. 2017, 8, 3198–3208. [Google Scholar] [CrossRef]
- Tan, L.; Liang, C.; Wang, Y.; Jiang, Y.; Zeng, S.; Tan, R. Pharmacodynamic effect of luteolin micelles on alleviating cerebral ischemia reperfusion injury. Pharmaceutics 2018, 10, 248. [Google Scholar] [CrossRef]
- Altamimi, M.A.; Hussain, A.; Alshehri, S.; Imam, S.S.; Alnemer, U.A. Development and evaluations of transdermally delivered luteolin loaded cationic nanoemulsion: In vitro and ex vivo evaluations. Pharmaceutics 2021, 13, 1218. [Google Scholar] [CrossRef]
- Koromili, M.; Kapourani, A.; Barmpalexis, P. Preparation and Evaluation of Amorphous Solid Dispersions for Enhancing Luteolin’s Solubility in Simulated Saliva. Polymers 2023, 15, 169. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Braidy, N.; Gortzi, O.; Sobarzo-Sanchez, E.; Daglia, M.; Skalicka-Woźniak, K.; Nabavi, S.M. Luteolin as an anti-inflammatory and neuroprotective agent: A brief review. Brain Res. Bull. 2015, 119, 1–11. [Google Scholar] [CrossRef]
- Pang, J.; Yang, F.; Zhang, Z.; Yang, W.; Li, Y.; Xu, H. The role of luteolin nanocomposites in rheumatoid arthritis treatment. Mater. Express 2021, 11, 303–309. [Google Scholar]
- Dia, V.P.; Pangloli, P. Epithelial-to-mesenchymal transition in paclitaxel-resistant ovarian cancer cells is downregulated by luteolin. J. Cell. Physiol. 2017, 232, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Y.; Cui, J.H.; Khan, H.; Aschner, M.; Batiha, G.E.S.; Jeandet, P. Luteolin and cancer metastasis suppression: Focus on the role of epithelial to mesenchymal transition. Med. Oncol. 2021, 38, 66. [Google Scholar] [CrossRef] [PubMed]
- Gilani, S.J.; Bin-Jumah, M.; Rizwanullah, M.; Imam, S.S.; Imtiyaz, K.; Alshehri, S.; Rizvi, M.M.A. Chitosan coated luteolin nanostructured lipid carriers: Optimization, In vitro-Ex vivo assessments and cytotoxicity study in breast cancer cells. Coatings 2021, 11, 158. [Google Scholar] [CrossRef]
- Chatzopoulos, G.S.; Karakostas, P.; Kavakloglou, S.; Assimopoulou, A.; Barmpalexis, P.; Tsalikis, L. Clinical Effectiveness of Herbal Oral Care Products in Periodontitis Patients: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 10061. [Google Scholar] [CrossRef]
- Gutiérrez-Venegas, G.; Kawasaki-Cárdenas, P.; Maldonado-Frías, S. Luteolin inhibits lipopolysaccharide actions on human gingival fibroblasts. Eur. J. Pharmacol. 2006, 541, 95–105. [Google Scholar] [CrossRef]
- Balci Yuce, H.; Toker, H.; Yildirim, A.; Tekin, M.B.; Gevrek, F.; Altunbas, N. The effect of luteolin in prevention of periodontal disease in Wistar rats. J. Periodontol. 2019, 90, 1481–1489. [Google Scholar] [CrossRef]
- Quan, H.; Dai, X.; Liu, M.; Wu, C.; Wang, D. Luteolin supports osteogenic differentiation of human periodontal ligament cells. BMC Oral Health 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Chagas, M.D.S.S.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Gonçalves-de-Albuquerque, C.F. Flavonols and Flavones as Potential anti-Inflammatory, Antioxidant, and Antibacterial Compounds. Oxid Med. Cell Longev. 2022, 6, 9966750. [Google Scholar] [CrossRef]
- Kariu, T.; Hamada, N.; Lakshmyya, K. Luteolin inhibits Porphyromonas gingivalis growth and alleviates alveolar bone destruction in experimental murine periodontitis. Biosci. Biotechnol. Biochem. 2023, zbad137. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plant Species | Family | Active Compound(s) | Concentration of Active Compound(s) | Reference |
|---|---|---|---|---|
| Apium graveolens | Apiaceae | luteolin luteolin-7-O-glucoside | 2.31 mg/100 g FW 654 mg/100 g DW | [24] |
| Asphodelus ramosus | Asphodelaceae | luteolin | 0.5 mg/g DW | [25] |
| Crepis incana | Asteraceae | luteolin, luteolin 7-O-β-d-glucopyranoside | 0.13 mg/g EXTR 0.22 mg/g EXTR | [26] |
| Cuminum cyminum | Apiaceae | luteolin | 3.1 mg/100 g DW | [27] |
| Cynara cardunculus | Asteraceae | luteolin 7-O-rutinoside, luteolin 7-O-glucoside, luteolin 7-O-malonylglycoside | 1.31–7.8 mg/g EXTR 1.52–8.5 mg/g EXTR 0.52–11.8 mg/g EXTR | [28] |
| Cynara humilis, Cynara cornigera | Asteraceae | luteolin, luteolin 7-O-β-d-glucoside, luteolin 7-O-β-d-rutinoside, luteolin 7-O-β-gentiobioside | NQ NQ NQ NQ | [29] |
| Juglans regia | Juglandaceae | luteolin | 2.4–3.4 μg/g DW | [30] |
| Laurus nobilis | Lauraceae | luteolin | 393.4 μg/g DW | [31] |
| Mentha spicata | Lamiaceae | luteolin | 19.89 μg/mL EXTR | [32] |
| Nepeta cataria | Lamiaceae | luteolin | 3.6 mg/100 g DW | [27] |
| Nepeta sibthorpii | Lamiaceae | luteolin 7-O-glucoside | 0.387 mg/g DW | [33] |
| Ocimum basilicum | Lamiaceae | luteolin 7-O-glucoside | 127 mg/100 g DW | [34] |
| Olea europaea | Oleaceae | luteolin | NQ | [35] |
| Olea europaea | Oleaceae | luteolin | 1–74 mg/kg FW | [36] |
| Olea europaea | Oleaceae | luteolin | 0.31–1.29 μmol/100 g virgin olive oil | [37] |
| Olea europaea | Oleaceae | luteolin 7-O-glucoside | NQ | [38] |
| Olea europaea | Oleaceae | luteolin | 0.11–1.69 mg/kg virgin olive oil | [39] |
| Olea europaea | Oleaceae | luteolin 4-O-glucoside, luteolin 7-O-glucoside | 0.07–1.60 mg/g FW 0.11–2.03 mg/g FW | [40] |
| Olea europaea | Oleaceae | luteolin | 92.4–118.16 μg/g FW | [41] |
| Olea europaea | Oleaceae | luteolin, luteolin 7-O-glucoside | 0.04 mg/g DW 0.21 mg/g DW | [42] |
| Origanum majorana | Lamiaceae | luteolin 7-O-glucoside | 461 mg/g DW | [34] |
| Origanum vulgare | Lamiaceae | luteolin | <LOQ | [43] |
| Origanum vulgare | Lamiaceae | luteolin | 25.97 μg/mL EXTR | [32] |
| Paeonia clusii | Paeoniaceae | luteolin, luteolin 3′-O-β-d-glucoside, luteolin 3′,4′-di-O-β-d-glucopyranoside | 0.69 mg/g DW 0.33 mg/g DW 0.35 mg/g DW | [44] |
| Petroselinum crispum | Apiaceae | luteolin, luteolin 7-O-glucoside, 3′-methyl-luteolin | 0.13–0.15 mg/100 g DW 0.27–0.45 mg/100 g DW 0.22–0.32 mg/100 g DW | [45] |
| Petroselinum sativum | Apiaceae | luteolin | 2.1 mg/100 g DW | [46] |
| Pistacia lentiscus | Anacardiaceae | luteolin | NQ | [47] |
| Rosmarinus officinalis | Lamiaceae | luteolin | 1.6 mg/100 g DW | [46] |
| Salvia fruticosa | Lamiaceae | luteolin | <LOQ | [43] |
| Salvia fruticosa | Lamiaceae | 6-hydroxy luteolin 7-O-glucoside, luteolin 7-O-glucuronide, luteolin 7-O-rutinoside, 6-methoxyluteolin 7-O-glucoside, 6-methoxyluteolin derivative, 6-methoxyluteolin derivative | <LOQ <LOQ 5.51–6.96 mg/g DW <LOQ <LOQ <LOQ | [48] |
| Salvia fruticosa, Salvia pomifera | Lamiaceae | luteolin 7-O-rutinoside, luteolin diglucuronide | 4.84 mg/g DW 6.18 mg/g DW | [49] |
| Salvia officinalis | Lamiaceae | luteolin | 0.2 mg/100 g DW | [46] |
| Satureja thymbra | Lamiaceae | luteolin 7,4′-di-O-glucuronide, 6-OH luteolin 7,3′-dimethyl ether, 6-OH luteolin 7,3′,4′-trimethyl ether | 15.6 g/kg DE 11.9–13.5 g/kg DE 11.9–13.3 g/kg DE | [50] |
| Spartium junceum | Fabaceae | luteolin | 2.5 mg/100 g DW | [27] |
| Teucrium chamaedrys | Lamiaceae | luteolin | 1.2 mg/100 g DW | [46] |
| Teucrium polium | Lamiaceae | luteolin | 0.48 mg/100 g DW | [51] |
| Thymus vulgaris | Lamiaceae | luteolin | 36 mg/100 g DW | [46] |
| Luteolin-7-O-glucoside (mg g−1 DW) | |||||
|---|---|---|---|---|---|
| N0 | N50 | N150 | N300 | Norg | |
| H1 | 45.67 * | 45.28 | 76.61 | 36.46 | 35.20 |
| H2 | 16.04 | 17.85 | 19.93 | 12.15 | 19.16 |
| H3 | 47.30 | 22.18 | 16.83 | 26.08 | 30.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arampatzis, A.S.; Pampori, A.; Droutsa, E.; Laskari, M.; Karakostas, P.; Tsalikis, L.; Barmpalexis, P.; Dordas, C.; Assimopoulou, A.N. Occurrence of Luteolin in the Greek Flora, Isolation of Luteolin and Its Action for the Treatment of Periodontal Diseases. Molecules 2023, 28, 7720. https://doi.org/10.3390/molecules28237720
Arampatzis AS, Pampori A, Droutsa E, Laskari M, Karakostas P, Tsalikis L, Barmpalexis P, Dordas C, Assimopoulou AN. Occurrence of Luteolin in the Greek Flora, Isolation of Luteolin and Its Action for the Treatment of Periodontal Diseases. Molecules. 2023; 28(23):7720. https://doi.org/10.3390/molecules28237720
Chicago/Turabian StyleArampatzis, Athanasios S., Aspasia Pampori, Eleftheria Droutsa, Maria Laskari, Panagiotis Karakostas, Lazaros Tsalikis, Panagiotis Barmpalexis, Christos Dordas, and Andreana N. Assimopoulou. 2023. "Occurrence of Luteolin in the Greek Flora, Isolation of Luteolin and Its Action for the Treatment of Periodontal Diseases" Molecules 28, no. 23: 7720. https://doi.org/10.3390/molecules28237720