
Phenolic Profile and Comparison of the Antioxidant, Anti-Ageing, Anti-Inflammatory, and Protective Activities of Borago officinalis Extracts on Skin Cells
 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
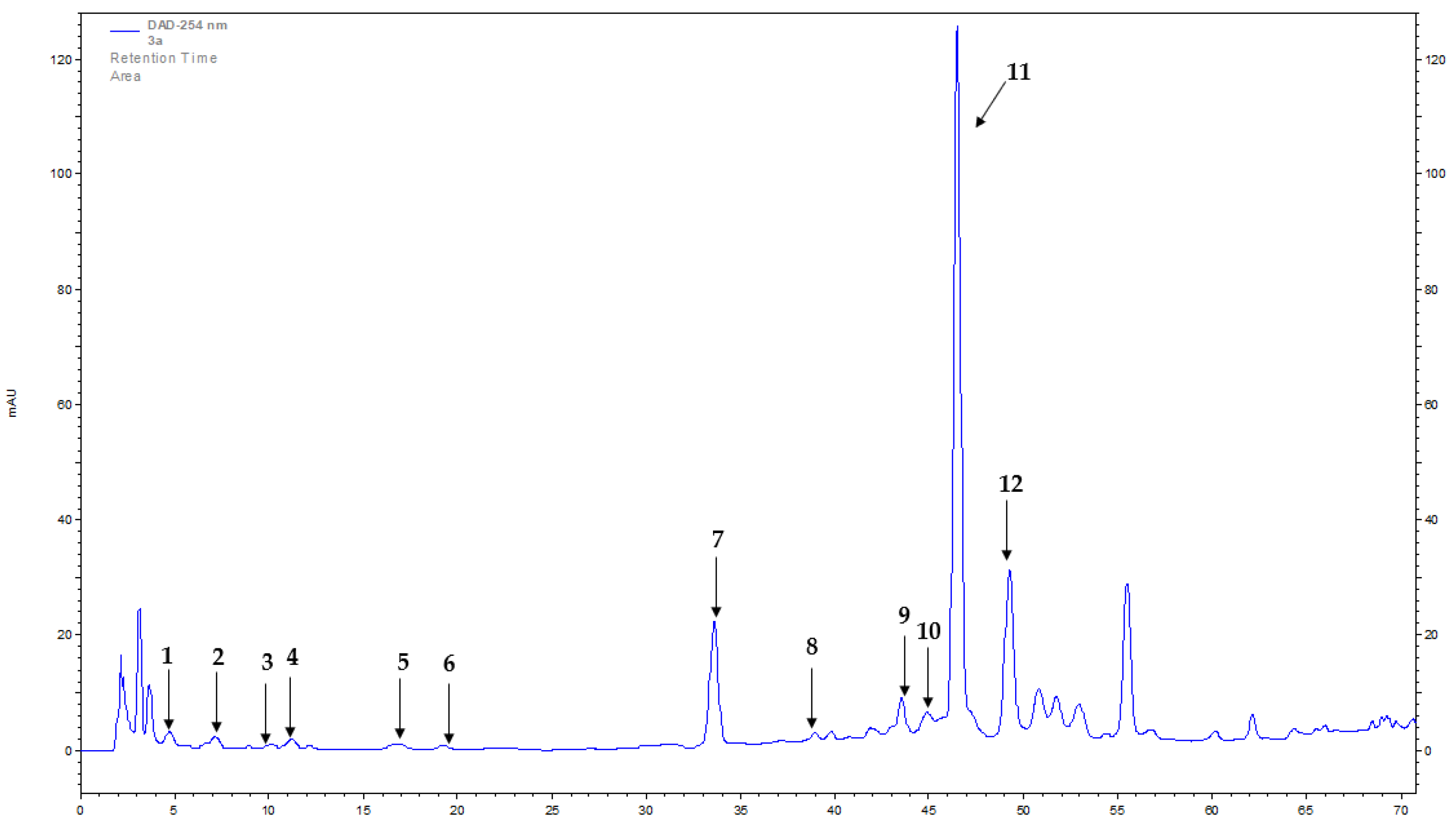
2.1. Phenolic Profiling
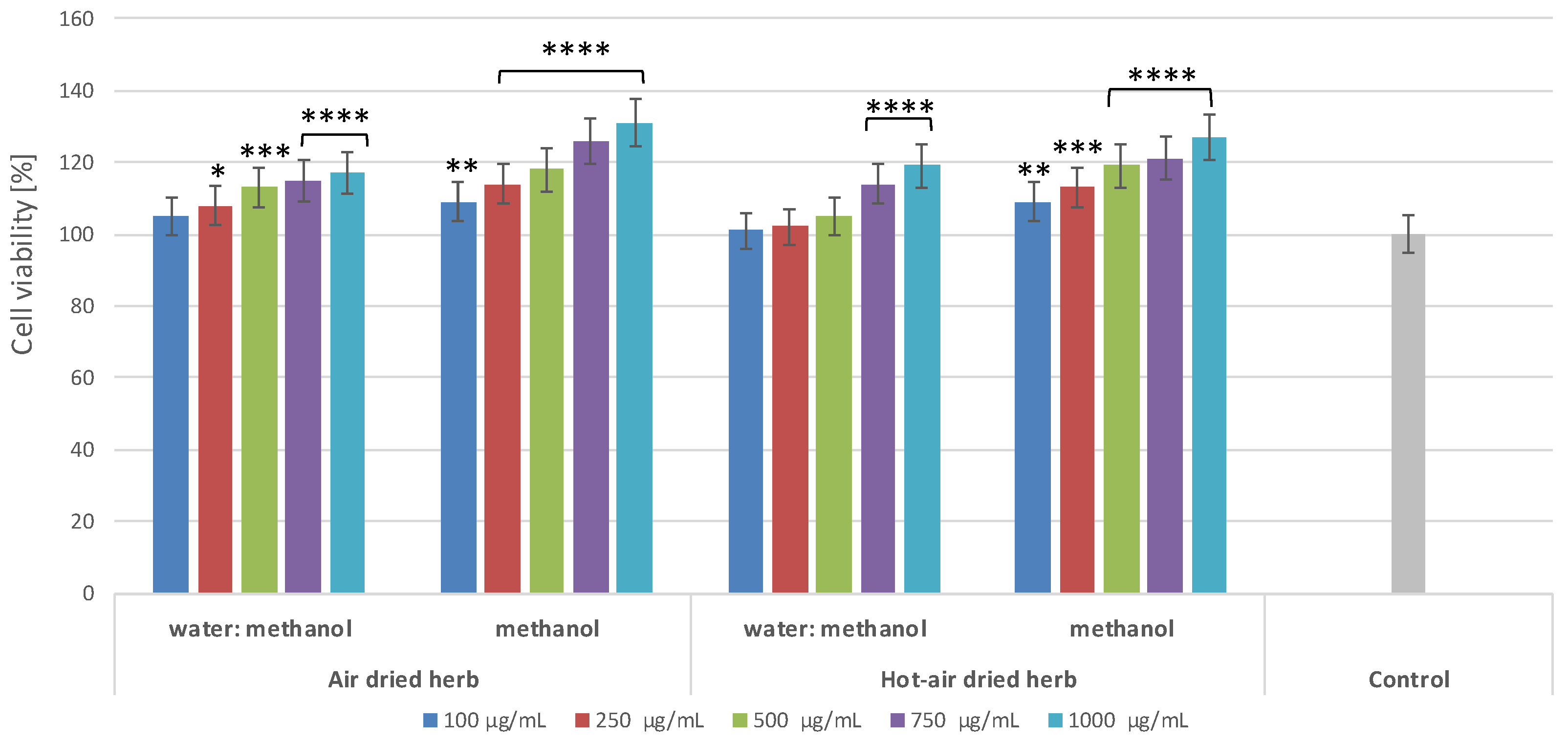
2.2. Effect of B. officinalis Extracts on Skin Cell Viability
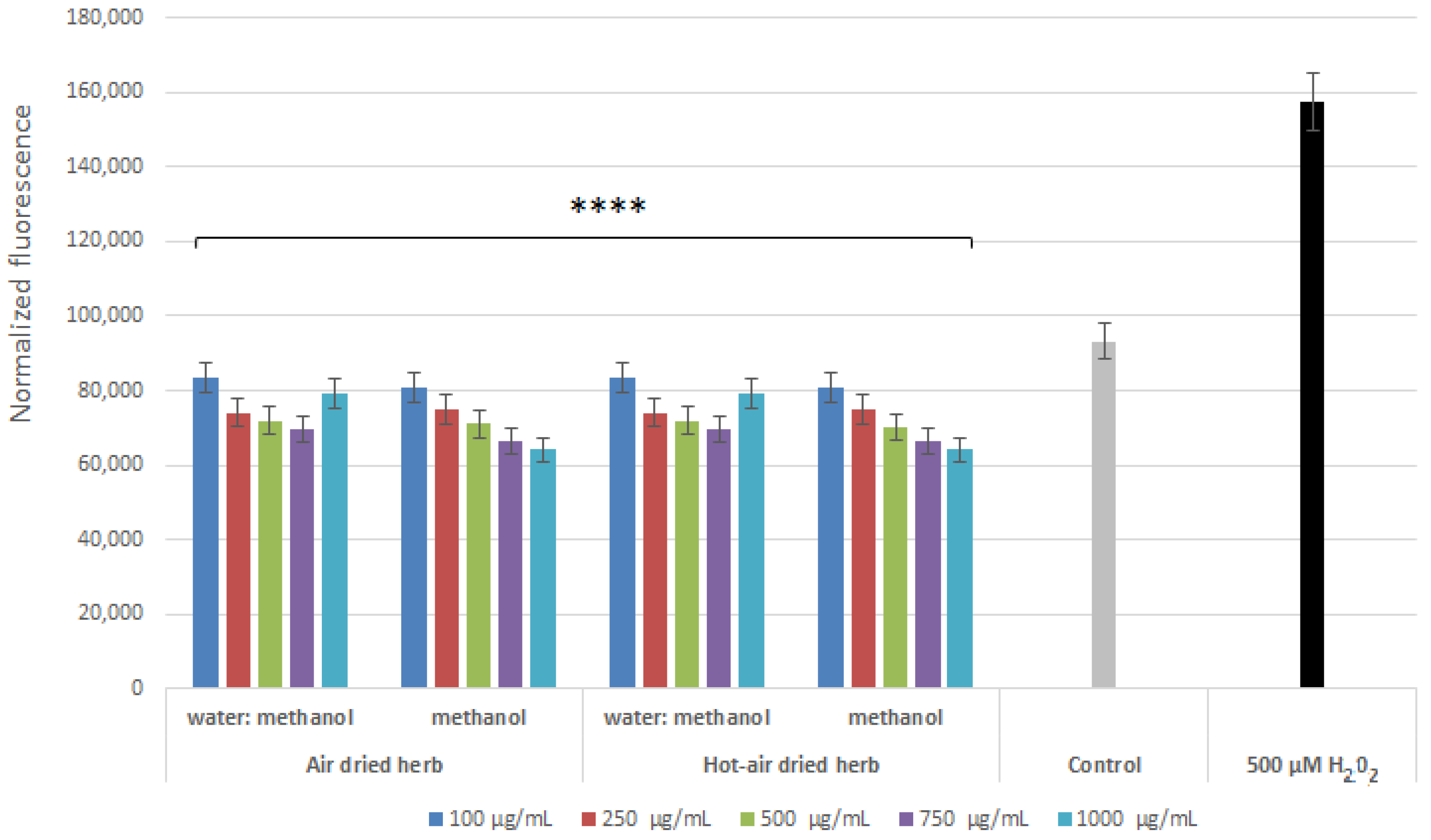
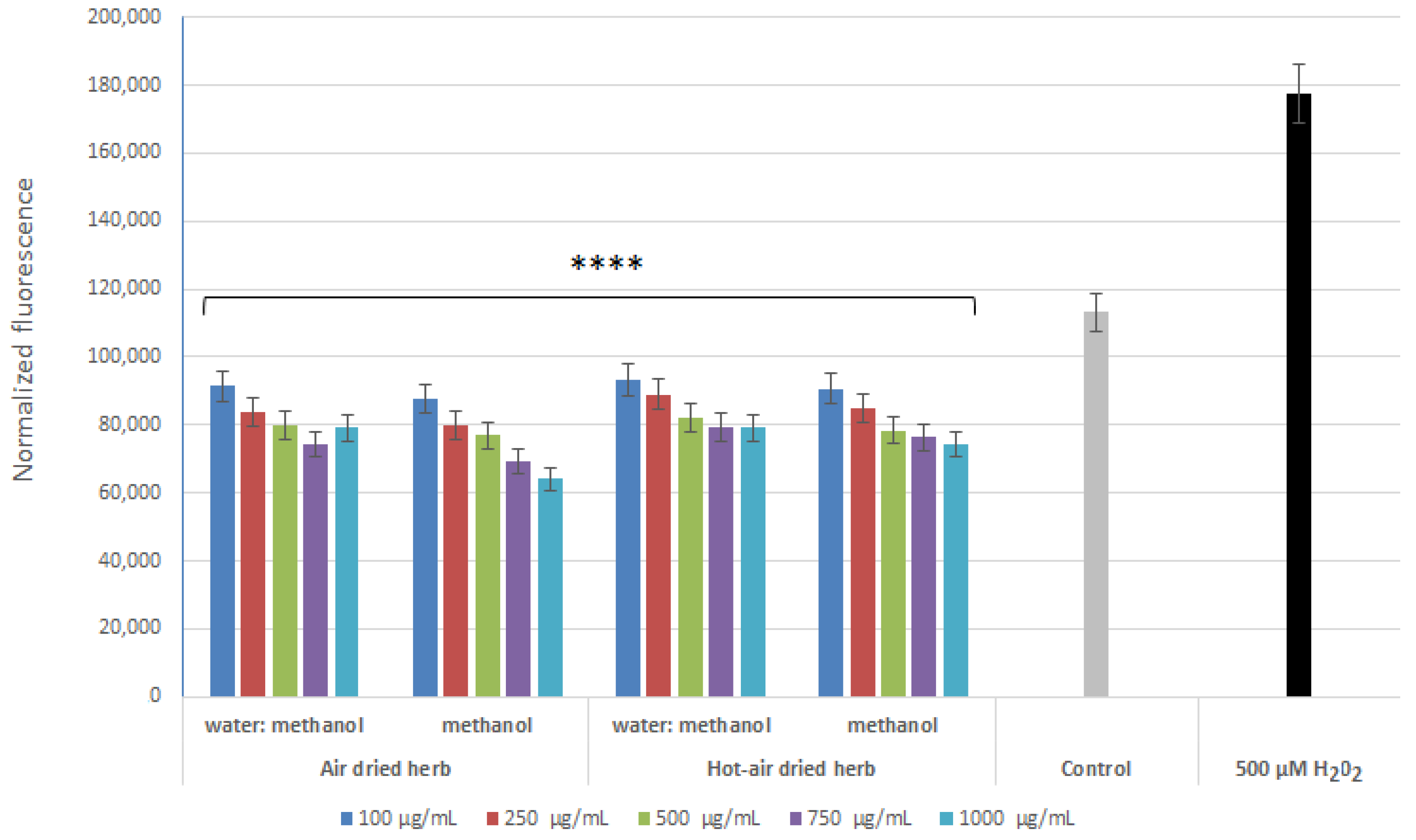
2.3. Intracellular ROS Levels in Skin Cells
2.4. Anti-Inflammatory Properties of B. officinalis
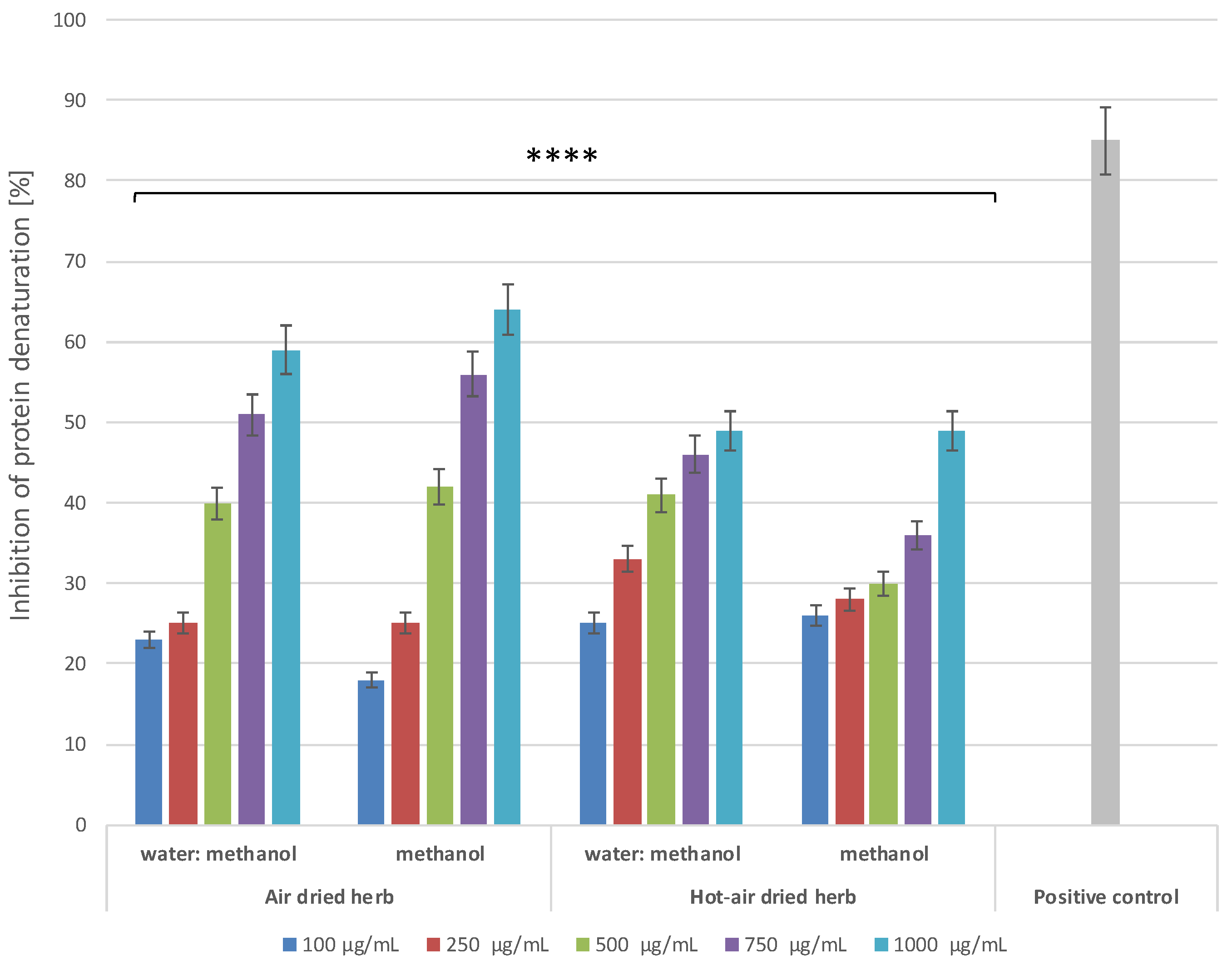
2.4.1. Inhibition of Protein Denaturation
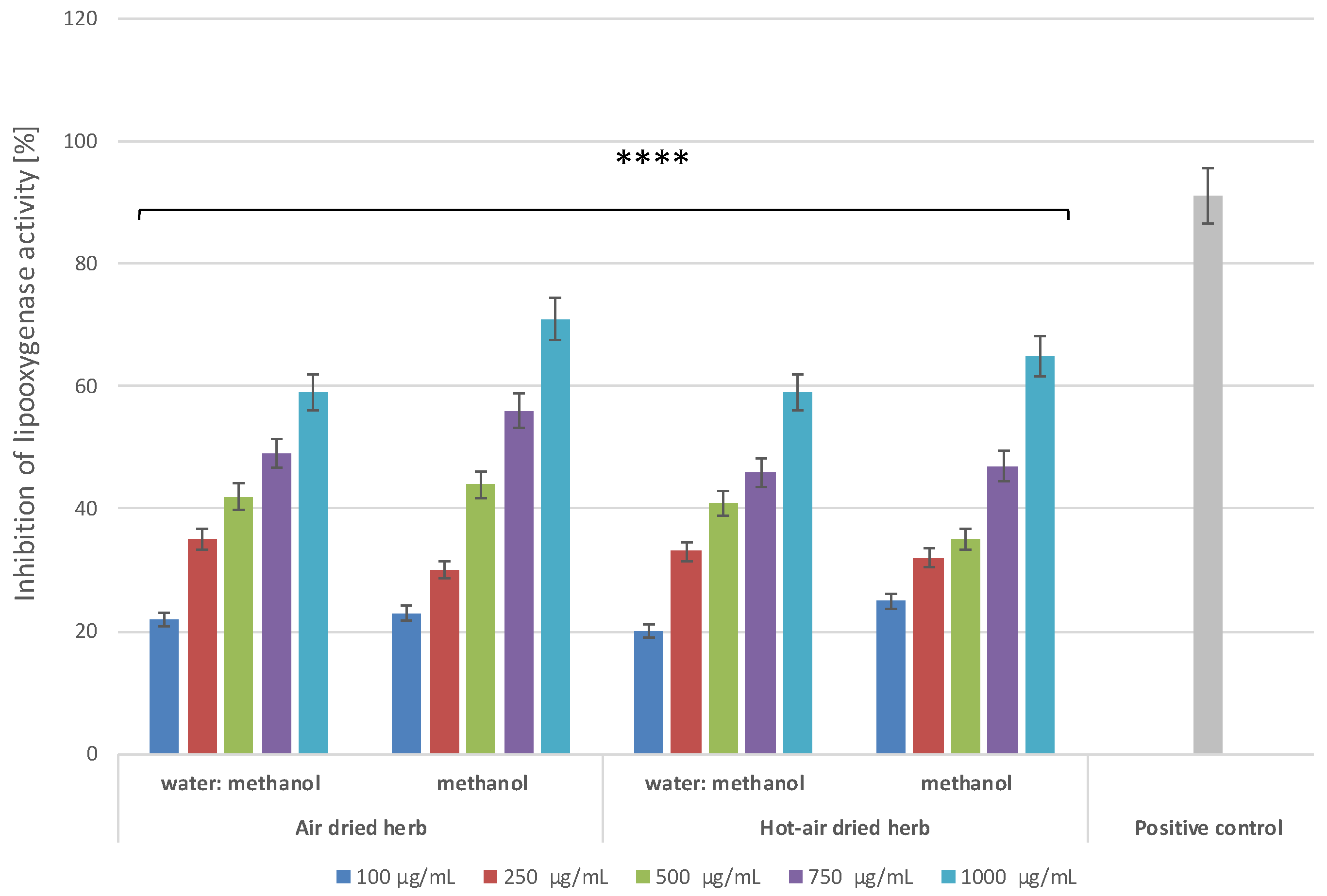
2.4.2. Lipoxygenase Inhibitory Activity
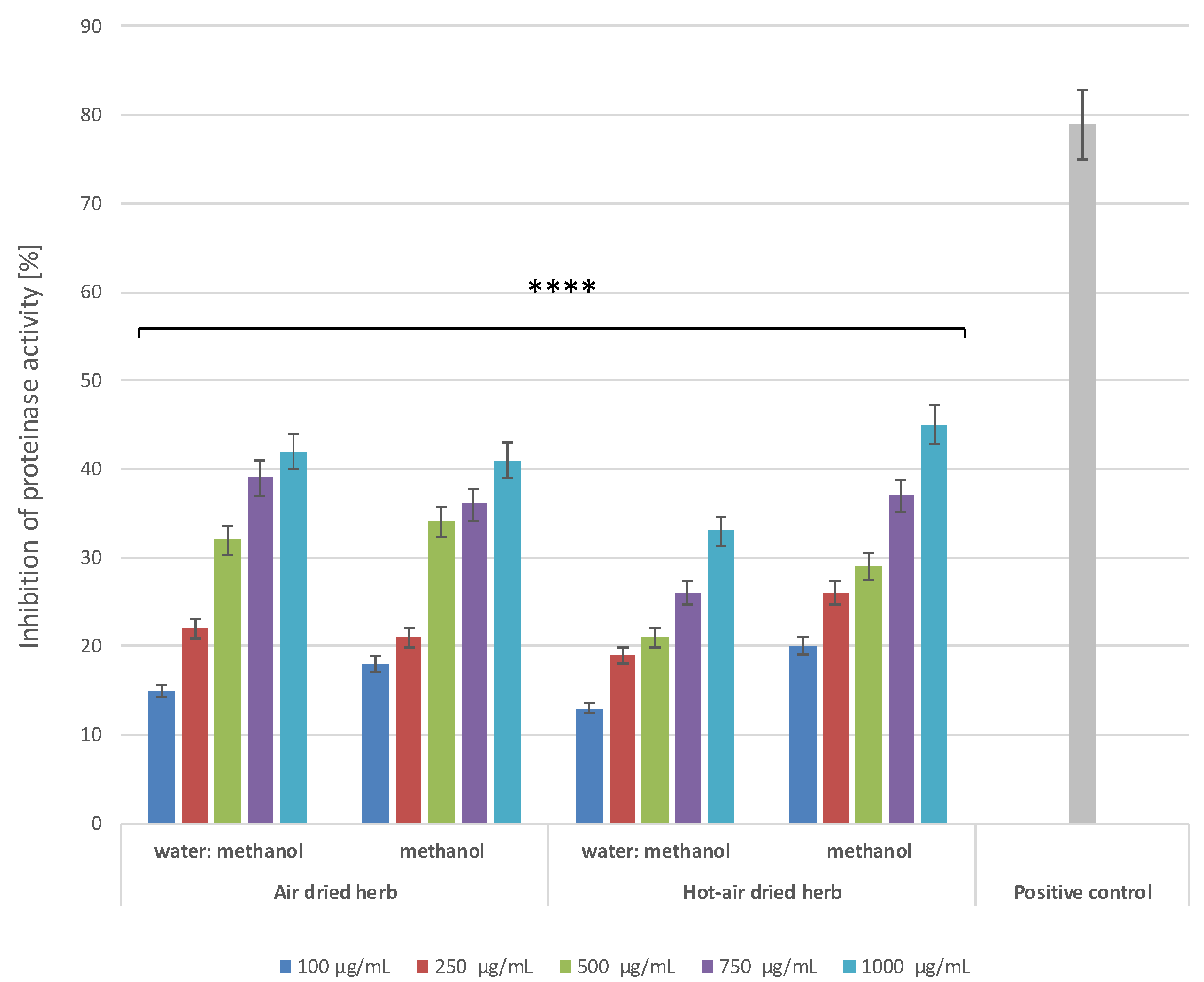
2.4.3. Proteinase Inhibitory Activity
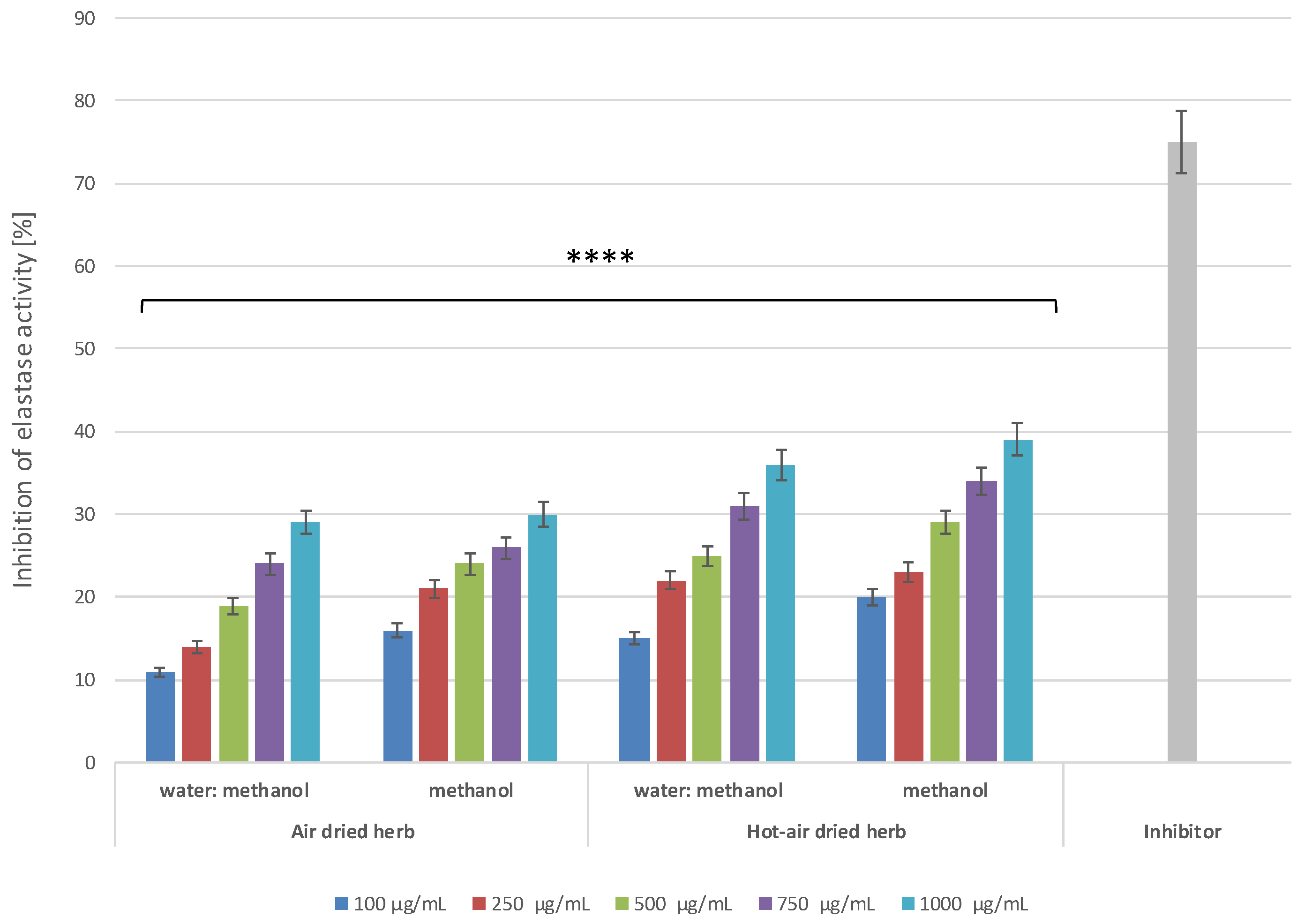
2.5. Anti-Ageing Properties of B. officinalis
3. Materials and Methods
3.1. Plant Material
3.2. Extract Preparation
3.3. Determination of Bioactive Compounds with HPLC-DAD
3.4. Cell Culture
3.5. Assessment of Cytotoxicity—Alamar Blue Assay
3.6. Detection of Intracellular Levels of Reactive Oxygen Species (ROS)
3.7. Anti-Inflammatory Activity
3.7.1. Assessment of Inhibition of Protein Denaturation
- As—the absorbance of the test sample,
- Ac—the absorbance of the control sample.
3.7.2. Assessment of Inhibition of Lipoxygenase Activity
- As—the absorbance of the test sample,
- Ac—the absorbance of the control sample.
3.7.3. Assessment of Inhibition of Proteinase Activity
- A1—the absorbance of the control sample,
- A2—the absorbance of the test sample.
3.8. Anti-Ageing Activity
3.8.1. Determination of Anti-Collagenase Activity
3.8.2. Determination of Anti-Elastase Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Karimi, E.; Oskoueian, E.; Karimi, A.; Noura, R.; Ebrahimi, M. Borago officinalis L. flower: A comprehensive study on bioactive compounds and its health-promoting properties. J. Food. Meas. Charact. 2017, 12, 826–838. [Google Scholar] [CrossRef]
- Pieszak, M.; Mikołajczak, P.Ł.; Manikowska, K. Borage (Borago officinalis L.)—A valuable medicinal plant used in herbal medicine. Herba Pol. 2012, 58, 95–103. [Google Scholar]
- Asadi-Samani, M.; Bahmani, M.; Rafieian-Kopaei, M. The chemical composition, botanical characteristic and biological activities of Borago officinalis: A review. Asian Pac. J. Trop. Med. 2014, 7, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kareem, A.T.; Hamad, M.N. Separation and identification of phenolic acid from Borago officinalis (F:Boraginaceae) cultivated in Iraq. Iraqi J. Pharm. Sci. 2020, 29, 139–151. [Google Scholar] [CrossRef]
- Abu-Qaoud, H.; Shawarb, N.; Hussen, F.; Jaradat, N.; Shtaya, M. Comparison of qualitative, quantitative analysis and antioxidant potential between wild and cultivated Borago officinalis leaves from palestine. Pak. J. Pharm. Sci. 2018, 31, 953–959. [Google Scholar] [PubMed]
- Zemmouri, H.; Ammar, S.; Boumendjel, A.; Messarah, M.; El Feki, A.; Bouaziz, M. Chemical composition and antioxidant activity of Borago officinalis L. leaf extract growing in Algeria. Arab. J. Chem. 2019, 12, 1954–1963. [Google Scholar] [CrossRef]
- Michalak, M. Plant-Derived Antioxidants: Significance in Skin Health and the Ageing Process. Int. J. Mol. Sci. 2022, 23, 585. [Google Scholar] [CrossRef]
- Michalak, M.; Pierzak, M.; Kręcisz, B.; Suliga, E. Bioactive Compounds for Skin Health: A Review. Nutrients 2021, 13, 203. [Google Scholar] [CrossRef]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Cytoprotective Effect of Ascorbic Acid and Rutin against Oxidative Changes in the Proteome of Skin Fibroblasts Cultured in a Three-Dimensional System. Nutrients 2020, 12, 1074. [Google Scholar] [CrossRef] [Green Version]
- Fernando, P.M.D.J.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Hewage, S.R.K.M.; Chae, S.W.; Hyun, J.W. Rosmarinic Acid Attenuates Cell Damage against UVB Radiation-Induced Oxidative Stress via Enhancing Antioxidant Effects in Human HaCaT Cells. Biomol. Ther. 2016, 24, 75–84. [Google Scholar] [CrossRef] [Green Version]
- You, O.H.; Shin, E.A.; Lee, H.; Kim, J.-H.; Sim, D.Y.; Kim, J.H.; Kim, Y.; Khil, J.-H.; Baek, N.-I.; Kim, S.-H. Apoptotic effect of astragalin in melanoma skin cancers via activation of caspases and inhibition of sry-related HMg-box gene 10. Phytother. Res. 2017, 31, 1614–1620. [Google Scholar] [CrossRef] [PubMed]
- Qattan, M.Y.; Khan, M.I.; Alharbi, S.H.; Verma, A.K.; Al-Saeed, F.A.; Abduallah, A.M.; Al Areefy, A.A. Therapeutic Importance of Kaempferol in the Treatment of Cancer through the Modulation of Cell Signalling Pathways. Molecules 2022, 27, 8864. [Google Scholar] [CrossRef] [PubMed]
- Gado, F.; Digiacomo, M.; Salsano, J.E.; Macchia, M.; Manera, C. Phenolic Compounds in Prevention and Treatment of Skin Cancers: A review. Curr. Med. Chem. 2021, 28, 6730–6752. [Google Scholar] [CrossRef]
- Li, W.; Hao, J.; Zhang, L.; Cheng, Z.; Deng, X.; Shu, G. Astragalin reduces hexokinase 2 through increasing miR-125b to inhibit the proliferation of hepatocellular carcinoma cells in vitro and in vivo. J. Agric. Food Chem. 2017, 65, 5961–5972. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, F.; Zha, D.; Wang, X.; Zhang, W.; He, Y.; Huang, Q.; Zhuang, H.; Hua, Z.-C. Astragalin-induced cell death is caspase-dependent and enhances the susceptibility of lung cancer cells to tumor necrosis factor by inhibiting the NF-κB pathway. Oncotarget 2017, 8, 26941–26958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moliner, C.; Cásedas, G.; Barros, L.; Finimundy, T.C.; Gómez-Rincón, C.; López, V. Neuroprotective Profile of Edible Flowers of Borage (Borago officinalis L.) in Two Different Models: Caenorhabditis elegans and Neuro-2a Cells. Antioxidants 2022, 11, 1244. [Google Scholar] [CrossRef]
- Seo, S.A.; Park, B.; Hwang, E.; Park, S.-Y.; Yi, T.-H. Borago officinalis L. attenuates UVB-induced skin photodamage via regulation of AP-1 and Nrf2/ARE pathway in normal human dermal fibroblasts and promotion of collagen synthesis in hairless mice. Exp. Gerontol. 2018, 107, 178–186. [Google Scholar] [CrossRef]
- Wettasinghe, M.; Shahidi, F. Antioxidant and free radical-scavenging properties of ethanolic extracts of defatted borage (Borago offcinalis L.) seeds. Food Chem. 1999, 67, 399–414. [Google Scholar] [CrossRef]
- Mirastschijski, U.; Lupše, B.; Maedler, K.; Sarma, B.; Radtke, A.; Belge, G.; Dorsch, M.; Wedekind, D.; McCawley, L.J.; Boehm, G.; et al. Matrix Metalloproteinase-3 is Key Effector of TNF-α-Induced Collagen Degradation in Skin. Int. J. Mol. Sci. 2019, 20, 5234. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.T. The antiinflammatory effect of an extract of Tripterygium wilfordii hook F on adjuvant-induced paw oedema in rats and inflammatory mediators release. Phytother. Res. 1997, 11, 152–154. [Google Scholar] [CrossRef]
- Gunathilake, K.D.P.P.; Ranaweera, K.K.D.S.; Rupasinghe, H.P.V. In Vitro Anti-Inflammatory Properties of Selected Green Leafy Vegetables. Biomedicines 2018, 19, 107. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Ma, S. Recent development of lipoxygenase inhibitors as anti-inflammatory agents. Med. Chem. Comm. 2017, 29, 212–225. [Google Scholar] [CrossRef]
- Assiry, A.A.; Bhavikatti, S.K.; Althobaiti, F.A.; Mohamed, R.N.; Karobari, M.I. Evaluation of In Vitro Antiprotease Activity of Selected Traditional Medicinal Herbs in Dentistry and Its In Silico PASS Prediction. Biomed. Res. Int. 2022, 2022, 5870443. [Google Scholar] [CrossRef]
- Nowak, A.; Zagórska-Dziok, M.; Perużyńska, M.; Cybulska, K.; Kucharska, E.; Ossowicz-Rupniewska, P.; Piotrowska, K.; Duchnik, W.; Kucharski, Ł.; Sulikowski, T.; et al. Assessment of the Anti-Inflammatory, Antibacterial and Anti-Aging Properties and Possible Use on the Skin of Hydrogels Containing Epilobium angustifolium L. Extracts. Front. Pharmacol. 2022, 13, 896706. [Google Scholar] [CrossRef] [PubMed]
- Asaad, G.F.; Redai, A.Q.; Hakami, A.O.; Ghazwani, F.H.; Nomier, Y.; Alshahrani, S. Potential analgesic and anti-inflammatory effect of Cuminum cyminum and Borago officinalis in rats and mice. Asian J. Pharm. Clin. Res. 2020, 13, 216–218. [Google Scholar]
- Barati, E.; Soleimani Asl, S.; Pourbakhsh, S.A.; Jamshidian, M.; Shahidi, S. Investigating the effect of Borago officnale on hipocampal IL-1 beta protein and gene in the amyloid β-peptide (25-35)-induced of inflammation in rat. Biomed. Pharmacol. J. 2015, 8, 937–943. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as Anti-Inflammatory Agents:Implications in Cancer and Cardiovascular Disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef]
- Ferrándiz, M.L.; Alcaraz, M.J. Anti-inflammatory activity and inhibition of arachidonic acid metabolism by flavonoids. Agents Actions 1991, 32, 283–288. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef]
- Sun, M.; Deng, Y.; Cao, X.; Xiao, L.; Ding, Q.; Luo, F.; Huang, P.; Gao, Y.; Liu, M.; Zhao, H. Effects of Natural Polyphenols on Skin and Hair Health: A Review. Molecules 2022, 27, 7832. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, N.; Yasuda, A.; Natsume, M.; Yoshikawa, T. Rosmarinic acid inhibits epidermal inflammatory responses: Anticarcinogenic effect of Perilla frutescens extract in the murine two-stage skin model. Carcinogenesis 2004, 25, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Lembo, S.; Balato, A.; Di Caprio, R.; Cirillo, T.; Giannini, V.; Gasparri, F.; Monfrecola, G. The modulatory effect of ellagic acid and rosmarinic acid on ultraviolet-B-induced cytokine/chemokine gene expression in skin keratinocyte (HaCaT) cells. Biomed. Res. Int. 2014, 2014, 346793. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, E.; Koh, J.; Kim, Y.S.; Park, D. Effect of rosmarinic acid on atopic dermatitis. J. Dermatol. 2008, 35, 768–771. [Google Scholar] [CrossRef]
- Ha, S.J.; Lee, J.; Park, J.; Kim, Y.H.; Lee, N.H.; Kim, Y.E.; Song, K.-M.; Chang, P.-S.; Jeong, C.-H.; Jung, S.K. Syringic acid prevents skin carcinogenesis via regulation of NoX and EGFR signaling. Biochem. Pharmacol. 2018, 154, 435–445. [Google Scholar] [CrossRef]
- Laronha, H.; Caldeira, J. Structure and function of human matrix metalloproteinases. Cells 2020, 9, 1076. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Khan, M.T.H.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, G. Molecular mechanisms of skin ageing. Mech. Ageing Dev. 2002, 123, 801–810. [Google Scholar] [CrossRef]
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Uyama, H.; Kobayashi, S. Inhibition effects of (+)-catechin-aldehyde polycondensates on proteinases causing proteolytic degradation of extracellular matrix. Biochem. Biophys. Res. Commun. 2004, 320, 256–261. [Google Scholar] [CrossRef]
- Szopa, A.; Starzec, A.; Ekiert, H. The importance of monochromatic lights in the production of phenolic acids and flavonoids in shoot cultures of Aronia melanocarpa, Aronia arbutifolia and Aronia × prunifolia. J. Photochem. Photobiol. B Biol. 2018, 179, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Ekiert, H. In vitro cultures of Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine)—A potential biotechnological rich source of therapeutically important phenolic acids. Appl. Biochem. Biotechnol. 2012, 166, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Wójciak, M.; Zagórska-Dziok, M.; Nizioł-Łukaszewska, Z.; Ziemlewska, A.; Furman-Toczek, D.; Szczepanek, D.; Sowa, I. In Vitro Evaluation of Anti-Inflammatory and Protective Potential of an Extract from Cornus mas L. Fruit against H2O2-Induced Oxidative Stress in Human Skin Keratinocytes and Fibroblasts. Int. J. Mol. Sci. 2022, 23, 13755. [Google Scholar] [CrossRef]
- Nizioł-Łukaszewska, Z.; Furman-Toczek, D.; Zagórska-Dziok, M. Antioxidant activity and cytotoxicity of Jerusalem artichoke tubers and leaves extract on HaCaT and BJ fibroblast cells. Lipids Health Dis. 2018, 17, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarveswaran, R.; Jayasuriya, W.J.A.B.; Suresh, T.S. In vitro assays to investigate the anti-inflammatory activity of herbal extracts: A review. World J. Pharm. Res. 2017, 6, 131–141. [Google Scholar]
- Ziemlewska, A.; Nizioł-Łukaszewska, Z.; Bujak, T.; Zagórska-Dziok, M.; Wójciak, M.; Sowa, I. Effect of fermentation time on the content of bioactive compounds with cosmetic and dermatological properties in Kombucha Yerba Mate extracts. Sci. Rep. 2021, 11, 18792. [Google Scholar] [CrossRef] [PubMed]
- Juvekar, A.; Sakat, S.; Wankhede, S.; Juvekar, M.; Gambhire, M. Evaluation of Antioxidant and Anti-Inflammatory Activity of Methanol Extract of Oxalis Corniculata. Planta Med. 2009, 75, PJ178. [Google Scholar] [CrossRef]
- Nizioł-Łukaszewska, Z.; Ziemlewska, A.; Bujak, T.; Zagórska-Dziok, M.; Zarębska, M.; Hordyjewicz-Baran, Z.; Wasilewski, T. Effect of Fermentation Time on Antioxidant and Anti-Ageing Properties of Green Coffee Kombucha Ferments. Molecules 2020, 25, 5394. [Google Scholar] [CrossRef]
- Zagórska-Dziok, M.; Ziemlewska, A.; Bujak, T.; Nizioł-Łukaszewska, Z.; Hordyjewicz-Baran, Z. Cosmetic and Dermatological Properties of Selected Ayurvedic Plant Extracts. Molecules 2021, 26, 614. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
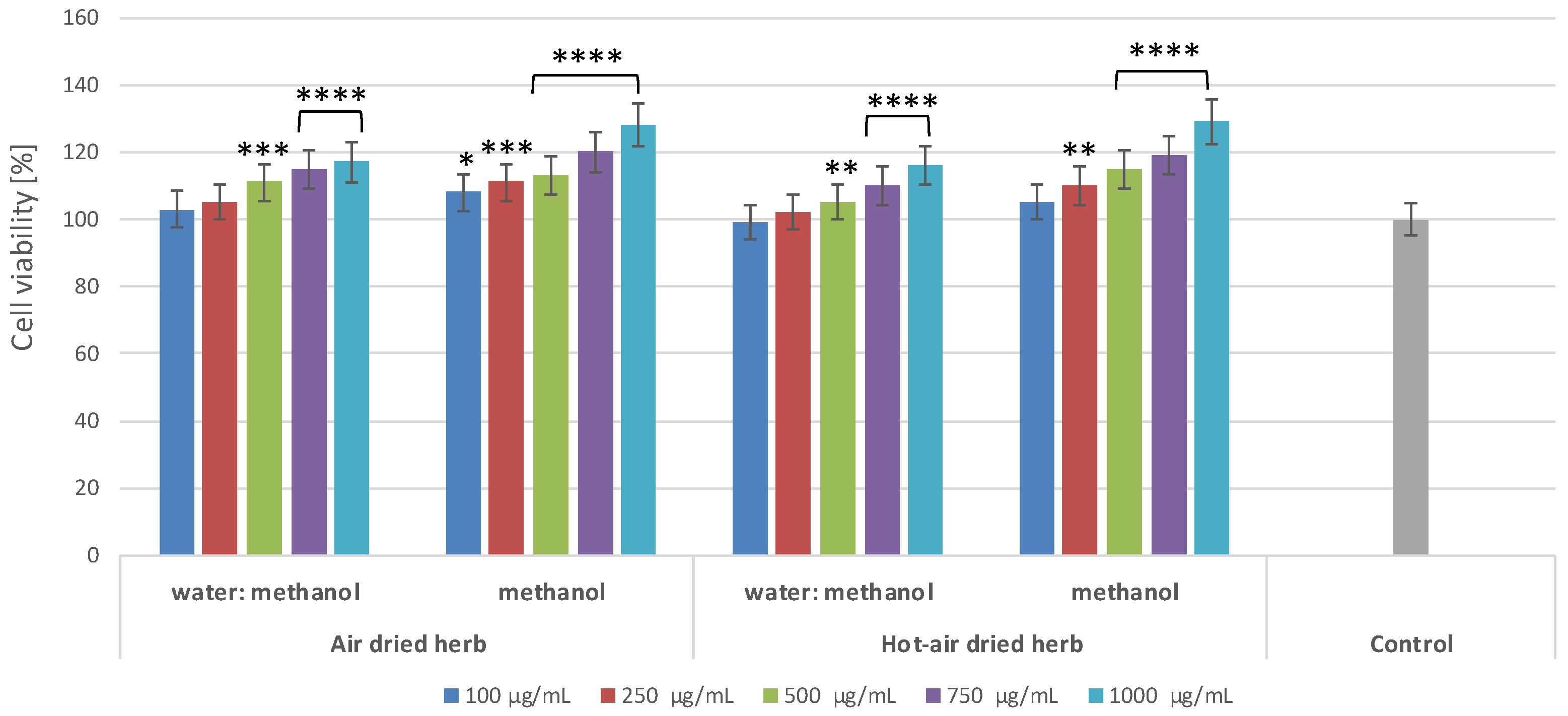{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
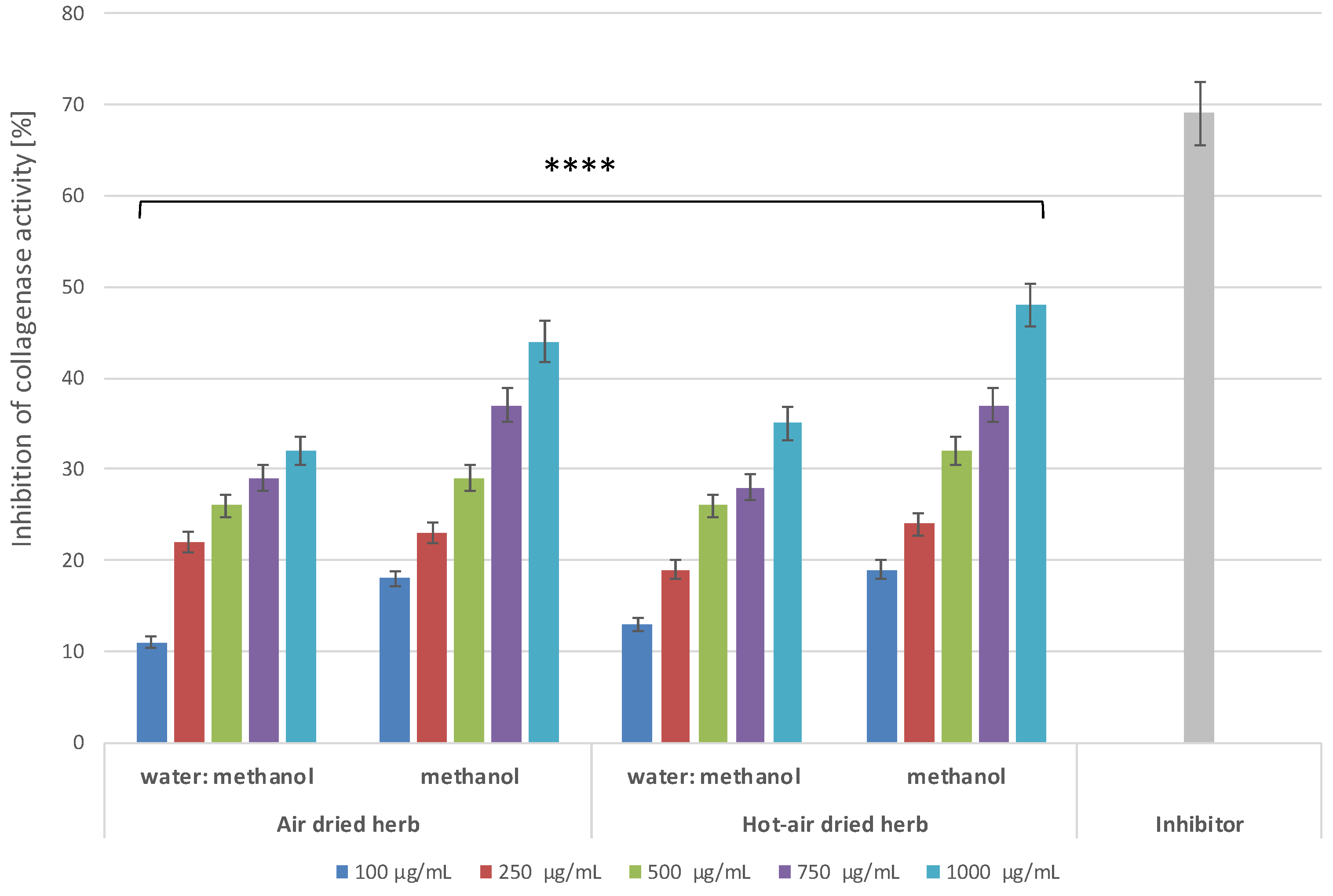{kind=link}
{kind=link}
| Flavonoids | Air-Dried Herb | Hot-Air-Dried Herb | ||
|---|---|---|---|---|
| Methanol Extract | Water:Methanol Extract (70:30) | Methanol Extract | Water:Methanol Extract (70:30) | |
| Astragalin | 147.40 ± 13.77 bcd | 48.96 ± 1.95 ac | 248.60 ± 12.47 abd | 61.54 ± 1.21 ac |
| Kaempferol 4-glucoside | 70.73 ± 11.58 bcd | 39.79 ± 1.38 acd | 141.08 ± 1.43 abd | 26.69 ± 1.36 ac |
| Rutoside | 77.09 ± 11.31 cd | 66.06 ± 2.72 cd | 95.71 ± 1.70 abd | 42.22 ± 0.65 abc |
| Vitexin | 40.84 ± 4.28 bd | 3.28 ± 0.30 ac | 42.84 ± 6.07 bd | 1.41 ± 0.08 ac |
| Total content | 336.06 ± 40.94 bcd | 158.09 ± 6.35 ac | 528.23 ± 21.67 abd | 131.86 ± 3.30 ac |
| Phenolic Acids | Air-Dried Herb | Hot-Air-Dried Herb | ||
|---|---|---|---|---|
| Methanol Extract | Water:Methanol Extract (70:30) | Methanol Extract | Water:Methanol Extract (70:30) | |
| Caffeic acid | 18.56 ± 0.94 bd | 28.66 ± 5.17 ac | 20.42 ± 5.29 bd | 25.13 ± 0.66 ac |
| Chlorogenic acid | 13.71 ± 4.78 bd | 33.90 ± 3.63 abc | 11.13 ± 2.76 bd | 23.63 ± 2.26 abc |
| 3,4-Dihydroxyphenylacetic acid | 101.48 ± 5.48 bcd | 260.17 ± 8.68 ad | 243.61 ± 18.25 ad | 189.88 ± 6.79 abc |
| Ferulic acid | 171.50 ± 16.49 bcd | 9.95 ± 0.63 ac | 205.30 ± 6.64 abd | 7.95 ± 0.41 ac |
| p-Hydroxybenzoic acid | 4.68 ± 0.56 bcd | 6.64 ± 0.40 acd | 5.71 ± 0.59 abd | 5.85 ± 0.17 ab |
| Protocatechuic acid | 8.80 ± 1.23 c | 10.30 ± 0.23 cd | 4.16 ± 0.42 abd | 8.06 ± 0.25 bc |
| Rosmarinic acid | 1173.80 ± 14.03 bcd | 309.22 ± 14.34 acd | 1783.55 ± 54.07 abd | 227.58 ± 9.93 abc |
| Syringic acid | 17.81 ± 1.08 bcd | 10.72 ± 1.06 ad | 7.31 ± 1.19 ad | 0.96 ± 0.07 abc |
| Total content | 1510.34 ± 44.59 bcd | 669.56 ± 34.14 acd | 2281.19 ± 89.21 abd | 489.04 ± 20.54 abc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalak, M.; Zagórska-Dziok, M.; Klimek-Szczykutowicz, M.; Szopa, A. Phenolic Profile and Comparison of the Antioxidant, Anti-Ageing, Anti-Inflammatory, and Protective Activities of Borago officinalis Extracts on Skin Cells. Molecules 2023, 28, 868. https://doi.org/10.3390/molecules28020868
Michalak M, Zagórska-Dziok M, Klimek-Szczykutowicz M, Szopa A. Phenolic Profile and Comparison of the Antioxidant, Anti-Ageing, Anti-Inflammatory, and Protective Activities of Borago officinalis Extracts on Skin Cells. Molecules. 2023; 28(2):868. https://doi.org/10.3390/molecules28020868
Chicago/Turabian StyleMichalak, Monika, Martyna Zagórska-Dziok, Marta Klimek-Szczykutowicz, and Agnieszka Szopa. 2023. "Phenolic Profile and Comparison of the Antioxidant, Anti-Ageing, Anti-Inflammatory, and Protective Activities of Borago officinalis Extracts on Skin Cells" Molecules 28, no. 2: 868. https://doi.org/10.3390/molecules28020868