
Analysis of Intestinal Metabolites in SR−B1 Knockout Mice via Ultra−Performance Liquid Chromatography Quadrupole Time−of−Flight Mass Spectrometry
 , ,
, ,
Abstract
:1. Introduction
2. Results
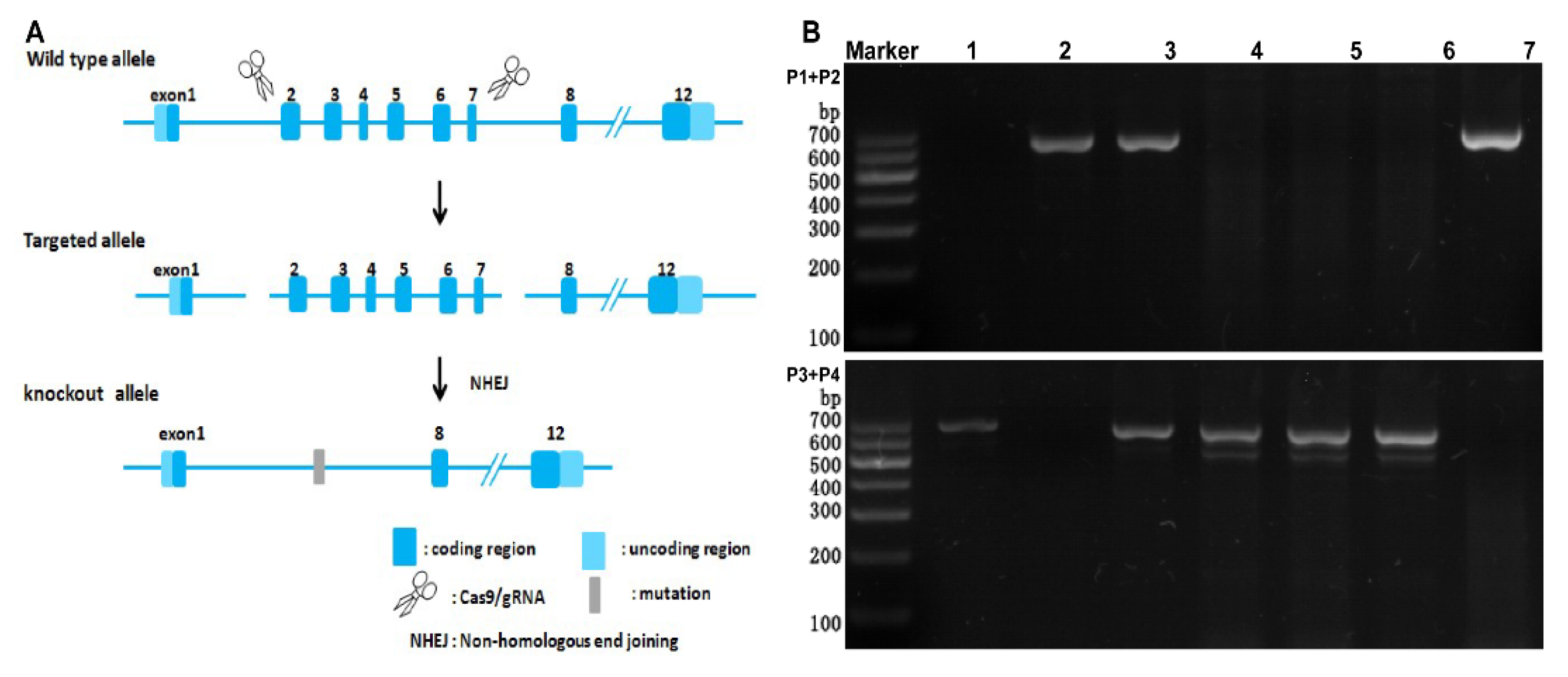
2.1. Detection of the Expression of SR−B1 mRNA by RT−PCR
2.2. Scavenger Receptor Class B Type 1 Deficiency Influences Intestinal Metabolomics
2.3. Scavenger Receptor Class B Type 1 Deficiency Influences the Amino Acid Contents in the Intestine
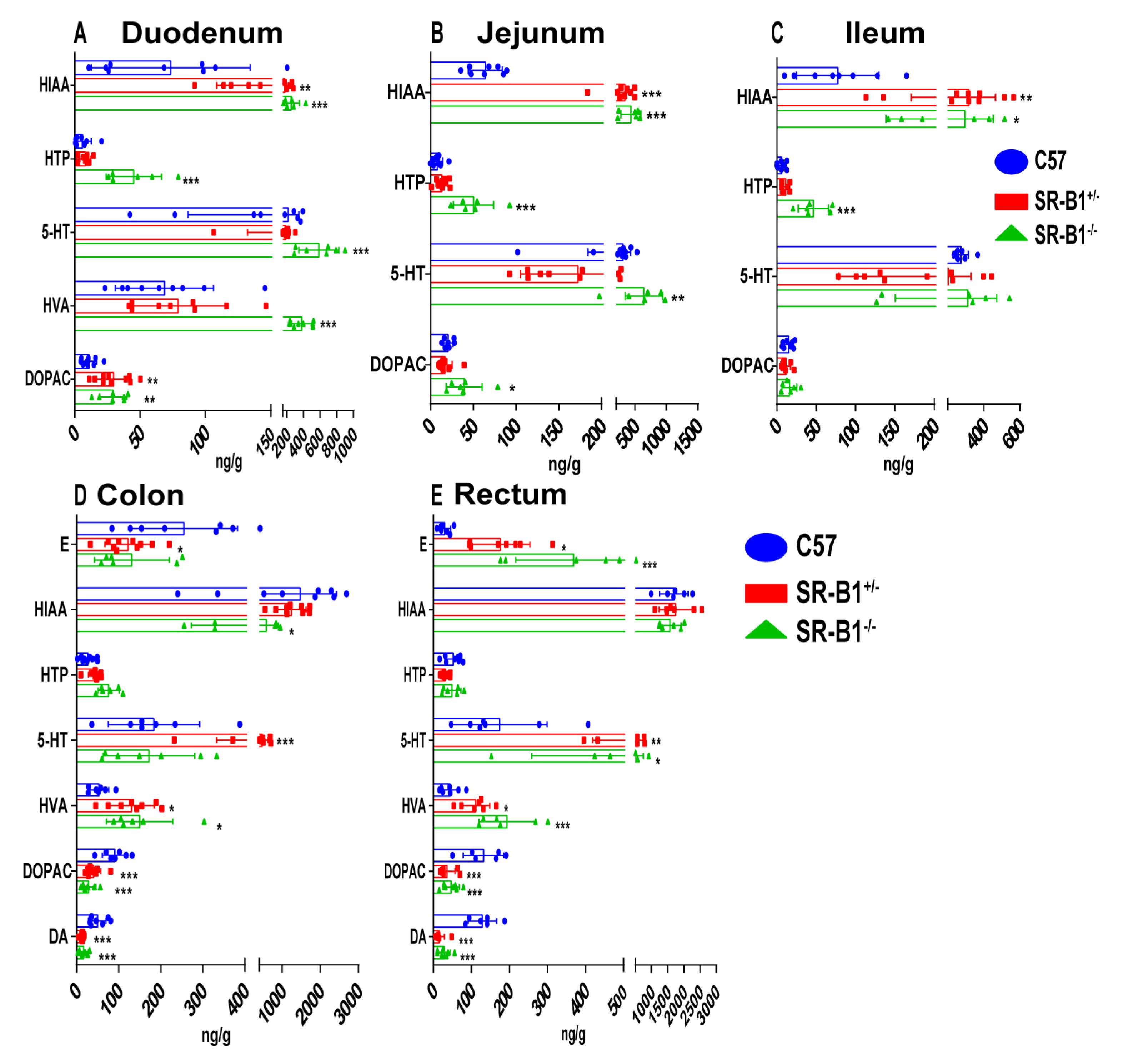
2.4. Scavenger Receptor Class B Type 1 Deficiency Influences the Neurotransmitter Contents in the Intestine
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Detection of SR−B1 mRNA Expression by RT–PCR
4.3. Metabolomics Analysis of the Intestines
4.4. Determination of Amino Acid Contents in the Intestines
4.5. Determination of Neurotransmitter Contents in the Intestines
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Huang, L.; Chambliss, K.L.; Gao, X.; Yuhanna, I.S.; Behling−Kelly, E.; Bergaya, S.; Ahmed, M.; Michaely, P.; Luby−Phelps, K.; Darehshouri, A.; et al. SR−B1 drives endothelial cell LDL transcytosis via DOCK4 to promote atherosclerosis. Nature 2019, 569, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yan, Z.; Hu, J.; Shen, W.J.; Azhar, S.; Kraemer, F.B. Scavenger receptor class B, type 1 facilitates cellular fatty acid uptake. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158554. [Google Scholar] [CrossRef] [PubMed]
- Baranova, I.N.; Vishnyakova, T.G.; Bocharov, A.V.; Leelahavanichkul, A.; Kurlander, R.; Chen, Z.; Souza, A.C.; Yuen, P.S.; Star, R.A.; Csako, G.; et al. Class B scavenger receptor types I and II and CD36 mediate bacterial recognition and proinflammatory signaling induced by Escherichia coli, lipopolysaccharide, and cytosolic chaperonin 60. J. Immunol. 2012, 188, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Landschulz, K.T.; Pathak, R.K.; Rigotti, A.; Krieger, M.; Hobbs, H.H. Regulation of scavenger receptor, class B, type I, a high density lipoprotein receptor, in liver and steroidogenic tissues of the rat. J. Clin. Investig. 1996, 98, 95–984. [Google Scholar] [CrossRef]
- Acton, S.; Rigotti, A.; Landschulz, K.T.; Xu, S.; Hobbs, H.H.; Krieger, M. Identification of scavenger receptor SR−BI as a high density lipoprotein receptor. Science 1996, 271, 518–520. [Google Scholar] [CrossRef]
- Cai, S.F.; Kirby, R.J.; Howles, P.N.; Hui, D.Y. Differentiation−dependent expression and localization of the class B type I scavenger receptor in intestine. J. Lipid Res. 2001, 42, 902–909. [Google Scholar] [CrossRef]
- Calvo, D.; Gómez−Coronado, D.; Lasunción, M.A.; Vega, M.A. CLA−1 is an 85−kD plasma membrane glycoprotein that acts as a high−affinity receptor for both native (HDL, LDL, and VLDL) and modified (OxLDL and AcLDL) lipoproteins. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 2341–2349. [Google Scholar] [CrossRef]
- Quiroz, A.; Molina, P.; Santander, N.; Gallardo, D.; Rigotti, A.; Busso, D. Ovarian cholesterol efflux: ATP−binding cassette transporters and follicular fluid HDL regulate cholesterol content in mouse oocytes. Biol. Reprod. 2020, 102, 348–361. [Google Scholar] [CrossRef]
- Van Eck, M.; Twisk, J.; Hoekstra, M.; Van Rij, B.T.; Van der Lans, C.A.; Bos, I.S.; Kruijt, J.K.; Kuipers, F.; Van Berkel, T.J. Differential effects of scavenger receptor BI deficiency on lipid metabolism in cells of the arterial wall and in the liver. J. Biol. Chem. 2003, 278, 23699–23705. [Google Scholar] [CrossRef]
- Rigotti, A.; Trigatti, B.L.; Penman, M.; Rayburn, H.; Herz, J.; Krieger, M. A targeted mutation in the murine gene encoding the high density lipoprotein (HDL) receptor scavenger receptor class B type I reveals its key role in HDL metabolism. Proc. Natl. Acad. Sci. USA 1997, 94, 12610–12615. [Google Scholar] [CrossRef]
- Seetharam, D.; Mineo, C.; Gormley, A.K.; Gibson, L.L.; Vongpatanasin, W.; Chambliss, K.L.; Hahner, L.D.; Cummings, M.L.; Kitchens, R.L.; Marcel, Y.L.; et al. High−density lipoprotein promotes endothelial cell migration and reendothelialization via scavenger receptor−B type I. Circ. Res. 2006, 98, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Li, X.A.; Titlow, W.B.; Jackson, B.A.; Giltiay, N.; Nikolova−Karakashian, M.; Uittenbogaard, A.; Smart, E.J. High density lipoprotein binding to scavenger receptor, Class B, type I activates endothelial nitric−oxide synthase in a ceramide−dependent manner. J. Biol. Chem. 2002, 277, 11058–11063. [Google Scholar] [CrossRef]
- Terasaka, N.; Wang, N.; Yvan−Charvet, L.; Tall, A.R. High−density lipoprotein protects macrophages from oxidized low−density lipoprotein−induced apoptosis by promoting efflux of 7−ketocholesterol via ABCG1. Proc. Natl. Acad. Sci. USA 2007, 104, 15093–15098. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Guo, L.; Wang, D.; Gao, H.; Hou, G.; Zheng, Z.; Ai, J.; Foreman, O.; Daugherty, A.; Li, X.A. Deficiency of scavenger receptor BI leads to impaired lymphocyte homeostasis and autoimmune disorders in mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2543–2551. [Google Scholar] [CrossRef]
- Grewal, T.; de Diego, I.; Kirchhoff, M.F.; Tebar, F.; Heeren, J.; Rinninger, F.; Enrich, C. High density lipoprotein−induced signaling of the MAPK pathway involves scavenger receptor type BI−mediated activation of Ras. J. Biol. Chem. 2003, 278, 16478–16481. [Google Scholar] [CrossRef] [PubMed]
- Baranova, I.N.; Kurlander, R.; Bocharov, A.V.; Vishnyakova, T.G.; Chen, Z.; Remaley, A.T.; Csako, G.; Patterson, A.P. Eggerman TL. Role of human CD36 in bacterial recognition, phagocytosis, and pathogen−induced JNK−mediated signaling. J. Immunol. 2008, 181, 7147–7156. [Google Scholar] [CrossRef]
- Hersoug, L.G.; Møller, P.; Loft, S. Gut microbiota−derived lipopolysaccharide uptake and trafficking to adipose tissue: Implications for inflammation and obesity. Obes. Rev. 2016, 17, 297–312. [Google Scholar] [CrossRef]
- Makide, K.; Uwamizu, A.; Shinjo, Y.; Ishiguro, J.; Okutani, M.; Inoue, A.; Aoki, J. Novel lysophosphoplipid receptors: Their structure and function. J. Lipid Res. 2014, 55, 1986–1995. [Google Scholar] [CrossRef]
- Wang, S.; Tang, K.; Lu, Y.; Tian, Z.; Huang, Z.; Wang, M.; Zhao, J.; Xie, J. Revealing the role of glycerophospholipid metabolism in asthma through plasma lipidomics. Clin. Chim. Acta 2021, 513, 34–42. [Google Scholar] [CrossRef]
- Kennelly, J.P.; Carlin, S.; Ju, T.; van der Veen, J.N.; Nelson, R.C.; Buteau, J.; Thiesen, A.; Richard, C.; Willing, B.P.; Jacobs, R.L. Intestinal Phospholipid Disequilibrium Initiates an ER Stress Response That Drives Goblet Cell Necroptosis and Spontaneous Colitis in Mice. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 999–1021. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, L.; Tan, H.; Zhang, Y.; Fang, Q.; Yang, J.; Cui, S.W.; Nie, S. Impact of pectin with various esterification degrees on the profiles of gut microbiota and serum metabolites. Appl. Microbiol. Biotechnol. 2022, 106, 3707–3720. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, X.; Qiu, J.; Zhou, A.; Xu, P.; Liu, Y.; Wu, H. A UHPLC−Q−TOF−MS−based serum and urine metabolomics approach reveals the mechanism of Gualou−Xiebai herb pair intervention against atherosclerosis process in ApoE−/− mice. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2022, 1215, 123567. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef] [PubMed]
- Petkevicius, K.; Virtue, S.; Bidault, G.; Jenkins, B.; Çubuk, C.; Morgantini, C.; Aouadi, M.; Dopazo, J.; Serlie, M.J.; Koulman, A.; et al. Accelerated phosphatidylcholine turnover in macrophages promotes adipose tissue inflammation in obesity. eLife 2019, 8, e47990. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, R.; Zhang, T.; Liu, F.; Zhang, W.; Wang, G.; Gu, G.; Han, Q.; Xu, D.; Yao, C.; et al. Identification of Lysophosphatidylcholines and Sphingolipids as Potential Biomarkers for Acute Aortic Dissection via Serum Metabolomics. Eur. J. Vasc. Endovasc. Surg. 2019, 57, 434–441. [Google Scholar] [CrossRef]
- Gonçalves, I.; Edsfeldt, A.; Ko, N.Y.; Grufman, H.; Berg, K.; Björkbacka, H.; Nitulescu, M.; Persson, A.; Nilsson, M.; Prehn, C.; et al. Evidence supporting a key role of Lp−PLA2−generated lysophosphatidylcholine in human atherosclerotic plaque inflammation. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1505–1512. [Google Scholar] [CrossRef]
- Alewijnse, A.E.; Peters, S.L. Sphingolipid signalling in the cardiovascular system: Good, bad or both? Eur. J. Pharmacol. 2008, 585, 292–302. [Google Scholar] [CrossRef]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar]
- Augé, N.; Maupas−Schwalm, F.; Elbaz, M.; Thiers, J.C.; Waysbort, A.; Itohara, S.; Krell, H.W.; Salvayre, R.; Nègre−Salvayre, A. Role for matrix metalloproteinase−2 in oxidized low−density lipoprotein−induced activation of the sphingomyelin/ceramide pathway and smooth muscle cell proliferation. Circulation 2004, 110, 571–578. [Google Scholar] [CrossRef]
- Brown, E.M.; Ke, X.; Hitchcock, D.; Jeanfavre, S.; Avila−Pacheco, J.; Nakata, T.; Arthur, T.D.; Fornelos, N.; Heim, C.; Franzosa, E.A.; et al. Bacteroides−Derived Sphingolipids Are Critical for Maintaining Intestinal Homeostasis and Symbiosis. Cell Host Microbe 2019, 25, 668–680.e7. [Google Scholar] [CrossRef]
- Yang, Z.H.; Nill, K.; Takechi−Haraya, Y.; Playford, M.P.; Nguyen, D.; Yu, Z.X.; Pryor, M.; Tang, J.; Rojulpote, K.V.; Mehta, N.N.; et al. Differential Effect of Dietary Supplementation with a Soybean Oil Enriched in Oleic Acid versus Linoleic Acid on Plasma Lipids and Atherosclerosis in LDLR−Deficient Mice. Int. J. Mol. Sci. 2022, 23, 8385. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Nagamine, R.; Tanaka, Y.; Tsai, W.T.; Jiang, Z.; Takeyama, A.; Imaizumi, K.; Sato, M. The effects of dietary linoleic acid on reducing serum cholesterol and atherosclerosis development are nullified by a high−cholesterol diet in male and female apoE−deficient mice. Br. J. Nutr. 2022, 1, 1–8. [Google Scholar] [CrossRef]
- Zhou, J.; Yao, N.; Wang, S.; An, D.; Cao, K.; Wei, J.; Li, N.; Zhao, D.; Wang, L.; Chen, X.; et al. Fructus Gardeniae−induced gastrointestinal injury was associated with the inflammatory response mediated by the disturbance of vitamin B6, phenylalanine, arachidonic acid, taurine and hypotaurine metabolism. J. Ethnopharmacol. 2019, 235, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, J.; Tan, K.S.; Deng, L.; Liu, F.; Tan, W. Isosteviol sodium ameliorates dextran sodium sulfate−induced chronic colitis through the regulation of metabolic profiling, macrophage polarization, and NF−κB pathway. Oxid. Med. Cell Longev. 2022, 2022, 4636618. [Google Scholar] [CrossRef] [PubMed]
- Chojnacki, C.; Popławski, T.; Konrad, P.; Fila, M.; Błasiak, J.; Chojnacki, J. Antimicrobial treatment improves tryptophan metabolism and mood of patients with small intestinal bacterial overgrowth. Nutr. Metab. 2022, 19, 66. [Google Scholar] [CrossRef]
- Sathyasaikumar, K.V.; Notarangelo, F.M.; Kelly, D.L.; Rowland, L.M.; Hare, S.M.; Chen, S.; Mo, C.; Buchanan, R.W.; Schwarcz, R. Tryptophan Challenge in Healthy Controls and People with Schizophrenia: Acute Effects on Plasma Levels of Kynurenine, Kynurenic Acid and 5−Hydroxyindoleacetic Acid. Pharmaceuticals 2022, 15, 1003. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Huang, C.Y.; Chen, C.M.; Liu, H.L. Focused Ultrasound−Induced Blood−Brain Barrier Opening Enhanced α−Synuclein Expression in Mice for Modeling Parkinson’s Disease. Pharmaceutics 2022, 14, 444. [Google Scholar] [CrossRef]
- Hosseini, R.; Fakhraei, N.; Malekisarvar, H.; Mansourpour, D.; Nili, F.; Farahani, M.; Dehpour, A.R. Effect of sumatriptan on acetic acid−induced experimental colitis in rats: A possible role for the 5−HT1B/1D receptors. Naunyn−Schmiedeberg’s Arch. Pharmacol. 2022, 395, 563–577. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut−Brain Axis. J. Cell. Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef]
- Gracia−Rubio, I.; Martín, C.; Civeira, F.; Cenarro, A. SR−B1, a Key Receptor Involved in the Progression of Cardiovascular Disease: A Perspective from Mice and Human Genetic Studies. Biomedicines 2021, 9, 612. [Google Scholar] [CrossRef]
- Hoekstra, M.; Sorci−Thomas, M. Rediscovering scavenger receptor type BI: Surprising new roles for the HDL receptor. Curr. Opin. Lipidol. 2017, 28, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.J.; Azhar, S.; Kraemer, F.B. SR−B1: A unique multifunctional receptor for cholesterol influx and efflux. Annu. Rev. Physiol. 2018, 80, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qu, C.; Li, T.; Cui, W.; Wang, X.; Du, J. Phagocytosis mediated by scavenger receptor class BI promotes macrophage transition during skeletal muscle regeneration. J. Biol. Chem. 2019, 294, 15672–15685. [Google Scholar] [CrossRef] [PubMed]
- Muehler, A.; Slizgi, J.R.; Kohlhof, H.; Groeppel, M.; Peelen, E.; Vitt, D. Clinical relevance of intestinal barrier dysfunction in common gastrointestinal diseases. World J. Gastrointest. Pathophysiol. 2020, 11, 114–130. [Google Scholar] [CrossRef]
- Chen, Y.; Cui, W.; Li, X.; Yang, H. Interaction between commensal bacteria, immune response and the intestinal barrier in inflammatory bowel disease. Front. Immunol. 2021, 12, 761981. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | A (%) | B (%) |
|---|---|---|
| 0.0 | 21.0 | 79.0 |
| 18.0 | 21.0 | 79.0 |
| 19.0 | 28.0 | 72.0 |
| 40.0 | 28.0 | 72.0 |
| 41.0 | 32.0 | 68.0 |
| 44.0 | 32.0 | 68.0 |
| 46.0 | 36.0 | 64.0 |
| 47.0 | 46.5 | 53.5 |
| 52.0 | 46.5 | 53.5 |
| 53.0 | 49.5 | 50.5 |
| 58.0 | 49.5 | 50.5 |
| 59.0 | 59.0 | 41.0 |
| 64.0 | 59.0 | 41.0 |
| 64.5 | 70.0 | 30.0 |
| 70.0 | 70.0 | 30.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Wang, L.; Chen, J.; Song, H.; Xing, W.; Wang, Z.; Song, X.; Yang, H.; Zhao, W. Analysis of Intestinal Metabolites in SR−B1 Knockout Mice via Ultra−Performance Liquid Chromatography Quadrupole Time−of−Flight Mass Spectrometry. Molecules 2023, 28, 610. https://doi.org/10.3390/molecules28020610
Chen Q, Wang L, Chen J, Song H, Xing W, Wang Z, Song X, Yang H, Zhao W. Analysis of Intestinal Metabolites in SR−B1 Knockout Mice via Ultra−Performance Liquid Chromatography Quadrupole Time−of−Flight Mass Spectrometry. Molecules. 2023; 28(2):610. https://doi.org/10.3390/molecules28020610
Chicago/Turabian StyleChen, Qijun, Lixue Wang, Jinlong Chen, Hui Song, Wen Xing, Ziqian Wang, Xueying Song, Hua Yang, and Wenhua Zhao. 2023. "Analysis of Intestinal Metabolites in SR−B1 Knockout Mice via Ultra−Performance Liquid Chromatography Quadrupole Time−of−Flight Mass Spectrometry" Molecules 28, no. 2: 610. https://doi.org/10.3390/molecules28020610