
Chromogenic Assay Is More Efficient in Identifying α-Amylase Inhibitory Properties of Anthocyanin-Rich Samples When Compared to the 3,5-Dinitrosalicylic Acid (DNS) Assay


 , , and
, , and
Abstract
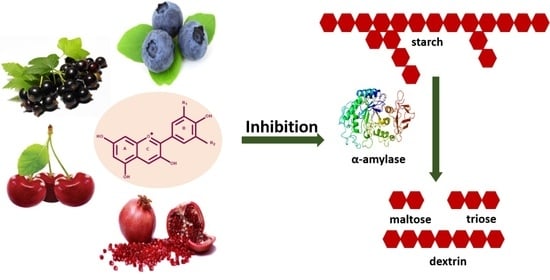
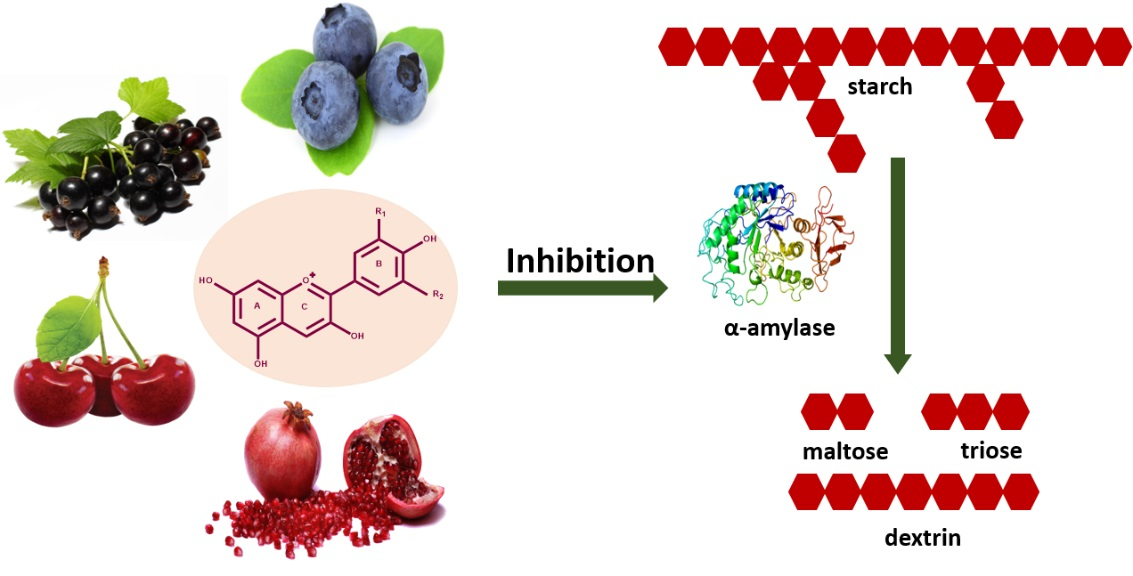
:
1. Introduction
2. Results and Discussion
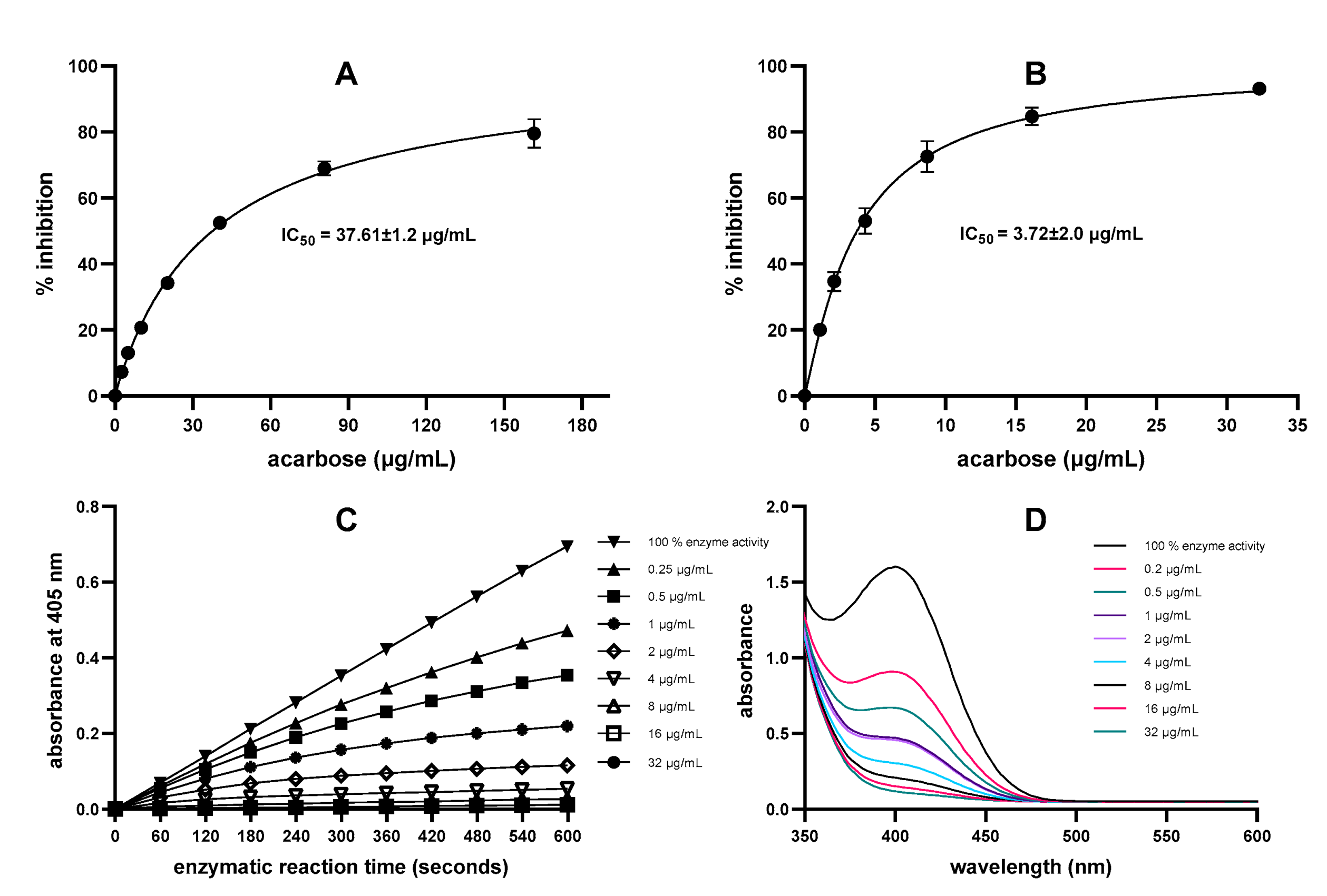
2.1. Comparison of DNS Assay and Direct Chromogenic Assay for α-Amylase Inhibition by Acarbose
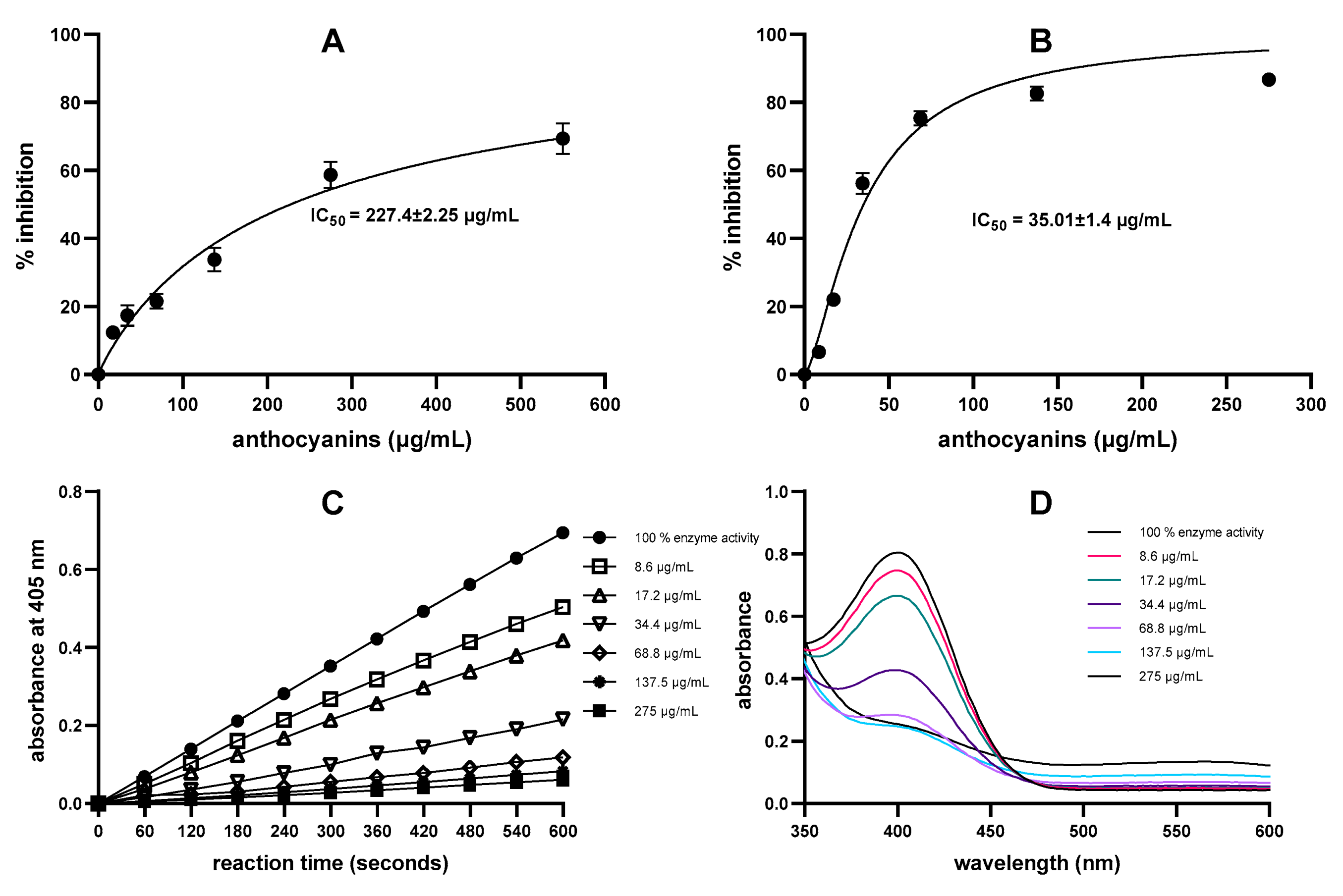
2.2. Comparison of DNS Assay and Direct Chromogenic Assay for α-Amylase Inhibition by Anthocyanin-Containing Samples
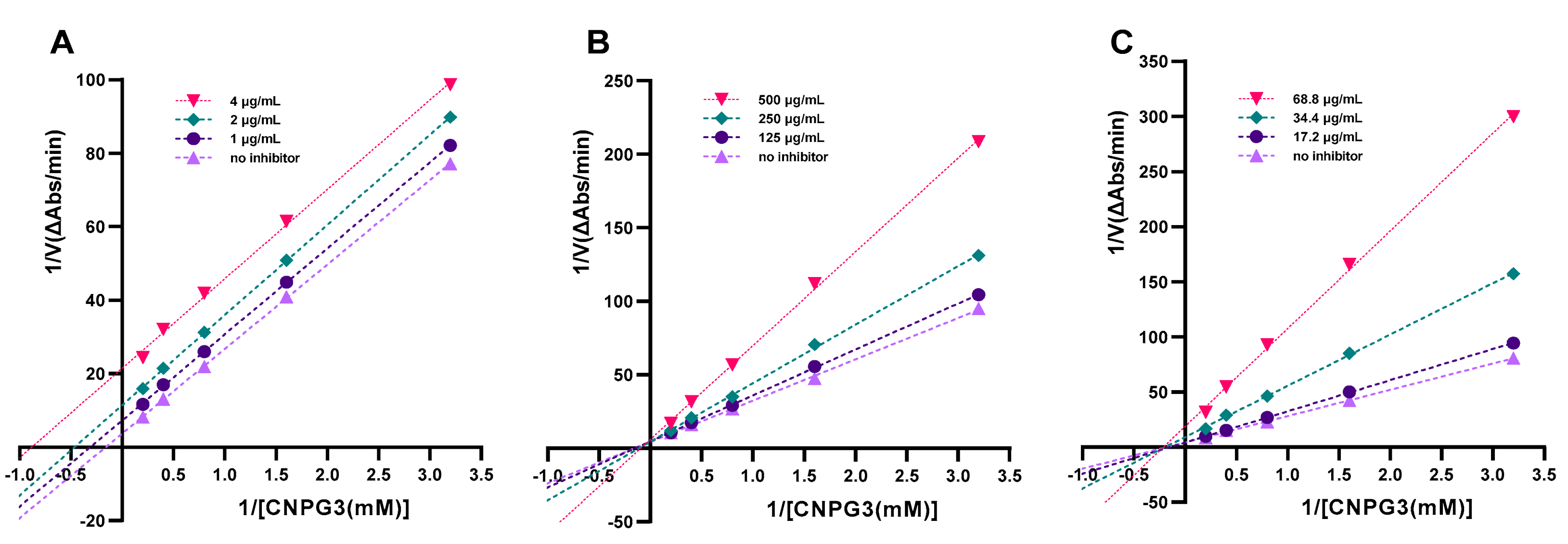
2.3. Mode of Inhibition of Anthocyanins on α-Amylase Activity
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Preparation and Analysis of Samples
3.3. DNS Assay
3.4. Direct Chromogenic Assay
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Konczak, I.; Zhang, W. Anthocyanins—More than nature’s colours. J. Biomed. Biotechnol. 2004, 2004, 239. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Ø.; Jordheim, M. Basic Anthocyanin Chemistry and Dietary Source. In Anthocyanins in Health and Disease; CRC Press: Boca Raton, FL, USA, 2013; pp. 13–90. [Google Scholar]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Putta, S.; Yarla, N.S.; Kumar, K.E.; Lakkappa, D.B.; Kamal, M.A.; Scotti, L.; Scotti, M.T.; Ashraf, G.M.; Rao, B.S.B.; Kumari, S.D.; et al. Preventive and Therapeutic Potentials of Anthocyanins in Diabetes and Associated Complications. Curr. Med. Chem. 2018, 25, 5347–5371. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, L.M.; Cazarin, C.B.B. In vitro and in vivo methods to predict carbohydrate bioaccessibility. Curr. Opin. Food Sci. 2021, 42, 69–75. [Google Scholar] [CrossRef]
- Williamson, G. Possible effects of dietary polyphenols on sugar absorption and digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef]
- Freitas, D.; Le Feunteun, S. Oro-gastro-intestinal digestion of starch in white bread, wheat-based and gluten-free pasta: Unveiling the contribution of human salivary alpha-amylase. Food Chem. 2019, 274, 566–573. [Google Scholar] [CrossRef]
- Sagandira, C.R.; Khasipo, A.Z.; Sagandira, M.B.; Watts, P. An overview of the synthetic routes to essential oral anti-diabetes drugs. Tetrahedron 2021, 96, 132378. [Google Scholar] [CrossRef]
- Ueno, H.; Tsuchimochi, W.; Wang, H.W.; Yamashita, E.; Tsubouchi, C.; Nagamine, K.; Sakoda, H.; Nakazato, M. Effects of Miglitol, Acarbose, and Sitagliptin on Plasma Insulin and Gut Peptides in Type 2 Diabetes Mellitus: A Crossover Study. Diabetes Ther. 2015, 6, 187–196. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Charoenlertkul, P.; Yibchok-Anun, S. alpha-Glucosidase inhibitory activity of cyanidin-3-galactoside and synergistic effect with acarbose. J. Enzym. Inhib. Med. Chem. 2009, 24, 65–69. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Ngamrojanavanich, N.; Kalampakorn, K.; Tiravanit, W.; Roengsumran, S.; Yibchok-Anun, S. Inhibitory activity of cyanidin-3-rutinoside on alpha-glucosidase. J. Enzym. Inhib. Med. Chem. 2004, 19, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Pinto, M.; Kwon, Y.-I.; Apostolidis, E.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Evaluation of Red Currants (Ribes rubrum L.), Black Currants (Ribes nigrum L.), Red and Green Gooseberries (Ribes Uva-Crispa) for Potential Management of Type 2 Diabetes and Hypertension Usingin Vitromodels. J. Food Biochem. 2010, 34, 639–660. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Hogan, S.; Chung, H.; Welbaum, G.E.; Zhou, K. Inhibitory effect of raspberries on starch digestive enzyme and their antioxidant properties and phenolic composition. Food Chem. 2010, 119, 592–599. [Google Scholar] [CrossRef]
- Akkarachiyasit, S.; Yibchok-Anun, S.; Wacharasindhu, S.; Adisakwattana, S. In vitro inhibitory effects of cyandin-3-rutinoside on pancreatic alpha-amylase and its combined effect with acarbose. Molecules 2011, 16, 2075–2083. [Google Scholar] [CrossRef]
- Visvanathan, R.; Qader, M.; Jayathilake, C.; Jayawardana, B.C.; Liyanage, R.; Sivakanesan, R. Critical review on conventional spectroscopic alpha-amylase activity detection methods: Merits, demerits, and future prospects. J. Sci. Food Agric. 2020, 100, 2836–2847. [Google Scholar] [CrossRef]
- Khadayat, K.; Marasini, B.P.; Gautam, H.; Ghaju, S.; Parajuli, N. Evaluation of the alpha-amylase inhibitory activity of Nepalese medicinal plants used in the treatment of diabetes mellitus. Clin. Phytoscience 2020, 6, 34. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of alpha-glucosidase and alpha-amylase by flavonoids. J. Nutr. Sci. 2006, 52, 149–153. [Google Scholar] [CrossRef]
- Visvanathan, R.; Houghton, M.J.; Williamson, G. Maltoheptaoside hydrolysis with chromatographic detection and starch hydrolysis with reducing sugar analysis: Comparison of assays allows assessment of the roles of direct alpha-amylase inhibition and starch complexation. Food Chem. 2021, 343, 128423. [Google Scholar] [CrossRef]
- Nyambe-Silavwe, H.; Villa-Rodriguez, J.A.; Ifie, I.; Holmes, M.; Aydin, E.; Jensen, J.M.; Williamson, G. Inhibition of human α-amylase by dietary polyphenols. J. Funct. Foods 2015, 19, 723–732. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Damager, I.; Numao, S.; Chen, H.; Brayer, G.D.; Withers, S.G. Synthesis and characterisation of novel chromogenic substrates for human pancreatic alpha-amylase. Carbohydr. Res. 2004, 339, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.; Ostberg-Potthoff, J.J.; Bakuradze, T.; Winterhalter, P.; Richling, E. Carbohydrate Hydrolase-Inhibitory Activity of Juice-Based Phenolic Extracts in Correlation to Their Anthocyanin/Copigment Profile. Molecules 2020, 25, 5224. [Google Scholar] [CrossRef]
- Robyt, J.F.; French, D. The Action Pattern of Porcine Pancreatic α-Amylase in Relationship to the Substrate Binding Site of the Enzyme. J. Biol. Chem. 1970, 245, 3917–3927. [Google Scholar] [CrossRef]
- Rauscher, E.; Neumann, U.; Schaich, E.; von Bülow, S.; Wahlefeld, A.W. Optimized conditions for determining activity concentration of alpha-amylase in serum, with 1,4-alpha-D-4-nitrophenylmaltoheptaoside as substrate. Clin. Chem. 1985, 31, 14–19. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of alpha-glucosidase, alpha-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-Anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal alpha-glucosidase and pancreatic alpha-amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Mo, J.; Ni, J.; Xu, Y.; Su, H.; Xie, J.; Chen, W. Structure-based design of human pancreatic amylase inhibitors from the natural anthocyanin database for type 2 diabetes. Food Funct. 2020, 11, 2910–2923. [Google Scholar] [CrossRef]
- Farooque, S.; Rose, P.M.; Benohoud, M.; Blackburn, R.S.; Rayner, C.M. Enhancing the Potential Exploitation of Food Waste: Extraction, Purification, and Characterization of Renewable Specialty Chemicals from Blackcurrants (Ribes nigrum L.). J. Agric. Food Chem. 2018, 66, 12265–12273. [Google Scholar] [CrossRef]
- Wilson, T.; Singh, A.P.; Vorsa, N.; Goettl, C.D.; Kittleson, K.M.; Roe, C.M.; Kastello, G.M.; Ragsdale, F.R. Human glycemic response and phenolic content of unsweetened cranberry juice. J. Med. Food 2008, 11, 46–54. [Google Scholar] [CrossRef]
- Torronen, R.; Kolehmainen, M.; Sarkkinen, E.; Mykkanen, H.; Niskanen, L. Postprandial glucose, insulin, and free fatty acid responses to sucrose consumed with blackcurrants and lingonberries in healthy women. Am. J. Clin. Nutr. 2012, 96, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Castro-Acosta, M.L.; Smith, L.; Miller, R.J.; McCarthy, D.I.; Farrimond, J.A.; Hall, W.L. Drinks containing anthocyanin-rich blackcurrant extract decrease postprandial blood glucose, insulin and incretin concentrations. J. Nutr. Biochem. 2016, 38, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Adisakwattana, S.; Ruengsamran, T.; Kampa, P.; Sompong, W. In vitro inhibitory effects of plant-based foods and their combinations on intestinal α-glucosidase and pancreatic α-amylase. BMC Complement. Altern. Med. 2012, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Barik, S.K.; Russell, W.R.; Moar, K.M.; Cruickshank, M.; Scobbie, L.; Duncan, G.; Hoggard, N. The anthocyanins in black currants regulate postprandial hyperglycaemia primarily by inhibiting alpha-glucosidase while other phenolics modulate salivary alpha-amylase, glucose uptake and sugar transporters. J. Nutr. Biochem. 2020, 78, 108325. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Zhang, Y.; Zhou, W. In vitro and in silico studies of the inhibition activity of anthocyanins against porcine pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- Bunea, A.; Rugina, D.; Sconta, Z.; Pop, R.M.; Pintea, A.; Socaciu, C.; Tabaran, F.; Grootaert, C.; Struijs, K.; VanCamp, J. Anthocyanin determination in blueberry extracts from various cultivars and their antiproliferative and apoptotic properties in B16-F10 metastatic murine melanoma cells. Phytochemistry 2013, 95, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Guillén, F.; Martínez-Romero, D.; Castillo, S.; Valero, D. Chemical Constituents and Antioxidant Activity of Sweet Cherry at Different Ripening Stages. J. Agric. Food Chem. 2005, 53, 2741–2745. [Google Scholar] [CrossRef]
- Mousavinejad, G.; Emam-Djomeh, Z.; Rezaei, K.; Khodaparast, M.H.H. Identification and quantification of phenolic compounds and their effects on antioxidant activity in pomegranate juices of eight Iranian cultivars. Food Chem. 2009, 115, 1274–1278. [Google Scholar] [CrossRef]
- Ifie, I.; Marshall, L.J.; Ho, P.; Williamson, G. Hibiscus sabdariffa (Roselle) Extracts and Wine: Phytochemical Profile, Physicochemical Properties, and Carbohydrase Inhibition. J. Agric. Food Chem. 2016, 64, 4921–4931. [Google Scholar] [CrossRef]
- Blando, F.; Calabriso, N.; Berland, H.; Maiorano, G.; Gerardi, C.; Carluccio, M.A.; Andersen, O.M. Radical Scavenging and Anti-Inflammatory Activities of Representative Anthocyanin Groupings from Pigment-Rich Fruits and Vegetables. Int. J. Food Sci. Nutr. 2018, 19, 169. [Google Scholar] [CrossRef]
- Bellesia, A.; Verzelloni, E.; Tagliazucchi, D. Pomegranate ellagitannins inhibit alpha-glucosidase activity in vitro and reduce starch digestibility under simulated gastro-intestinal conditions. Int. J. Food Sci. Nutr. 2015, 66, 85–92. [Google Scholar] [CrossRef]
- McDougall, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit alpha-amylase and alpha-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef] [PubMed]
- Grussu, D.; Stewart, D.; McDougall, G.J. Berry polyphenols inhibit α-amylase in vitro: Identifying active components in rowanberry and raspberry. J. Agric. Food Chem. 2011, 59, 2324–2331. [Google Scholar] [CrossRef]
- Gusakov, A.V.; Kondratyeva, E.G.; Sinitsyn, A.P. Comparison of two methods for assaying reducing sugars in the determination of carbohydrase activities. Int. J. Anal. Chem. 2011, 2011, 283658. [Google Scholar] [CrossRef] [PubMed]
- Nyambe-Silavwe, H.; Williamson, G. Chlorogenic and phenolic acids are only very weak inhibitors of human salivary α-amylase and rat intestinal maltase activities. Food Res. Int. (Ott. Ont.) 2018, 113, 452–455. [Google Scholar] [CrossRef]
- Promyos, N.; Temviriyanukul, P.; Suttisansanee, U. Investigation of Anthocyanidins and Anthocyanins for Targeting alpha-Glucosidase in Diabetes Mellitus. Prev. Nutr. Food Sci. 2020, 25, 263–271. [Google Scholar] [CrossRef]
- Matsui, T.; Ueda, T.; Oki, T.; Sugita, K.; Terahara, N.; Matsumoto, K. alpha-Glucosidase inhibitory action of natural acylated anthocyanins. 1. Survey of natural pigments with potent inhibitory activity. J. Agric. Food Chem. 2001, 49, 1948–1951. [Google Scholar] [CrossRef] [PubMed]
- Kaeswurm, J.A.H.; Könighofer, L.; Hogg, M.; Scharinger, A.; Buchweitz, M. Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine α-Amylase by Anthocyanin-3-Glycosides. Foods 2020, 9, 367. [Google Scholar] [CrossRef]
- Dona, A.C.; Pages, G.; Gilbert, R.G.; Kuchel, P.W. Digestion of starch: In vivo and in vitro kinetic models used to characterise oligosaccharide or glucose release. Carbohydr. Polym. 2010, 80, 599–617. [Google Scholar] [CrossRef]
- Homoki, J.R.; Nemes, A.; Fazekas, E.; Gyemant, G.; Balogh, P.; Gal, F.; Al-Asri, J.; Mortier, J.; Wolber, G.; Babinszky, L.; et al. Anthocyanin composition, antioxidant efficiency, and alpha-amylase inhibitor activity of different Hungarian sour cherry varieties (Prunus cerasus L.). Food Chem. 2016, 194, 222–229. [Google Scholar] [CrossRef]
- Ji, Y.; Liu, D.; Jin, Y.; Zhao, J.; Zhao, J.; Li, H.; Li, L.; Zhang, H.; Wang, H. In vitro and in vivo inhibitory effect of anthocyanin-rich bilberry extract on α-glucosidase and α-amylase. LWT 2021, 145, 111484. [Google Scholar] [CrossRef]
- Karim, Z.; Holmes, M.; Orfila, C. Inhibitory effect of chlorogenic acid on digestion of potato starch. Food Chem. 2017, 217, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, S.L.; Ellis, P.R.; Butterworth, P.J. An investigation of the action of porcine pancreatic α-amylase on native and gelatinised starches. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2001, 1525, 29–36. [Google Scholar] [CrossRef]
- Fernando, G.S.N.; Wood, K.; Papaioannou, E.H.; Marshall, L.J.; Sergeeva, N.N.; Boesch, C. Application of an Ultrasound-Assisted Extraction Method to Recover Betalains and Polyphenols from Red Beetroot Waste. ACS Sustain. Chem. Eng. 2021, 9, 8736–8747. [Google Scholar] [CrossRef]
- Sindi, H.A.; Marshall, L.J.; Morgan, M.R.A. Comparative chemical and biochemical analysis of extracts of Hibiscus sabdariffa. Food Chem. 2014, 164, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, S.; Marshall, J.L.; Boesch, C. Hibiscus sabdariffa inhibits α-glucosidase activity in vitro and lowers postprandial blood glucose response in humans. Hum. Nutr. Metab. 2022, 30, 200164. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts (Pure/Crude) | DNS Assay (IC50/% Inhibition) | Direct Chromogenic Assay (IC50/% Inhibition) | Predominant Anthocyanins |
|---|---|---|---|
| Blackcurrant (purified) * | 227.4 ± 2.25 µg/mL | 35.01 ± 1.4 µg/mL | Dp3rut, Cy3rut, Dp3glu, Cy3glu [29] |
| Blueberry | NO (645 μg/mL) | 80.44 ± 2.0 µg/mL | Dp3glu, Mv3glu, Dp3glc, Mv3glc [36] |
| Cherry | NO (at 536 μg/mL) | 30% at 268 μg/mL | Cy3rut, Cy3glu [37] |
| Pomegranate | NO (at 31 μg/mL) | 11.33 ± 2.3 μg/mL | Dp3,5 diglu, Cy3,5 diglu, Dp3glu [38] |
| Hibiscus | NO (at 218 μg/mL) | NO (at 218 μg/mL) | Dp3sam, Cy3sam [39] |
| Mahaleb cherry (purified) * | ND | 5% (at 34 μg/mL) | Cy3glu [40] |
| Mahaleb cherry (crude) | ND | 21% (at 39 μg/mL) | Cy3glu [40] |
| Black carrot (purfied) * | ND | NO (at 57.7 μg/mL) | Cy3glc [40] |
| Black carrot (crude) | ND | NO (at 48 μg/mL) | Cy3glc [40] |
| Cyanidin | 7% | 141 ± 1.6 μg/mL (491 μM) | |
| Cyanidin-3-O-galactopyranoside | ND | 414 ± 2.6 μg/mL (845 μM) |
| Type of Concentrate | Polyphenols (mg/mL) | Anthocyanins (mg/mL) | Total Sugars (mg/mL) |
|---|---|---|---|
| Blueberry | 18.96 ± 0.37 b | 11 ± 0.08 a | 576.6 ± 1.7 b |
| Cherry | 9.83 ± 0.36 c | 3.01 ± 0.02 b | 502.3 ± 0.85 c |
| Hibiscus | 7.66 ± 0.15 c | 4.4 ± 0.03 b | 17.42 ± 0.74 d |
| Pomegranate | 23.32 ± 1.55 a | 1.85 ± 0.01 c | 636.5 ± 8.82 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulfiqar, S.; Blando, F.; Orfila, C.; Marshall, L.J.; Boesch, C. Chromogenic Assay Is More Efficient in Identifying α-Amylase Inhibitory Properties of Anthocyanin-Rich Samples When Compared to the 3,5-Dinitrosalicylic Acid (DNS) Assay. Molecules 2023, 28, 6399. https://doi.org/10.3390/molecules28176399
Zulfiqar S, Blando F, Orfila C, Marshall LJ, Boesch C. Chromogenic Assay Is More Efficient in Identifying α-Amylase Inhibitory Properties of Anthocyanin-Rich Samples When Compared to the 3,5-Dinitrosalicylic Acid (DNS) Assay. Molecules. 2023; 28(17):6399. https://doi.org/10.3390/molecules28176399
Chicago/Turabian StyleZulfiqar, Sadia, Federica Blando, Caroline Orfila, Lisa J. Marshall, and Christine Boesch. 2023. "Chromogenic Assay Is More Efficient in Identifying α-Amylase Inhibitory Properties of Anthocyanin-Rich Samples When Compared to the 3,5-Dinitrosalicylic Acid (DNS) Assay" Molecules 28, no. 17: 6399. https://doi.org/10.3390/molecules28176399