
Hemoglobin Derivatives in Beef Irradiated with Accelerated Electrons
 , , , and
, , , and
Abstract
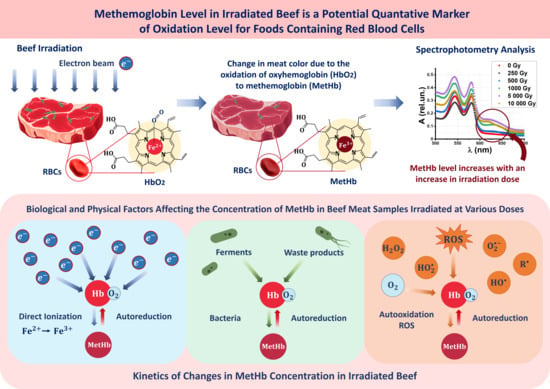
:
1. Introduction
2. Results
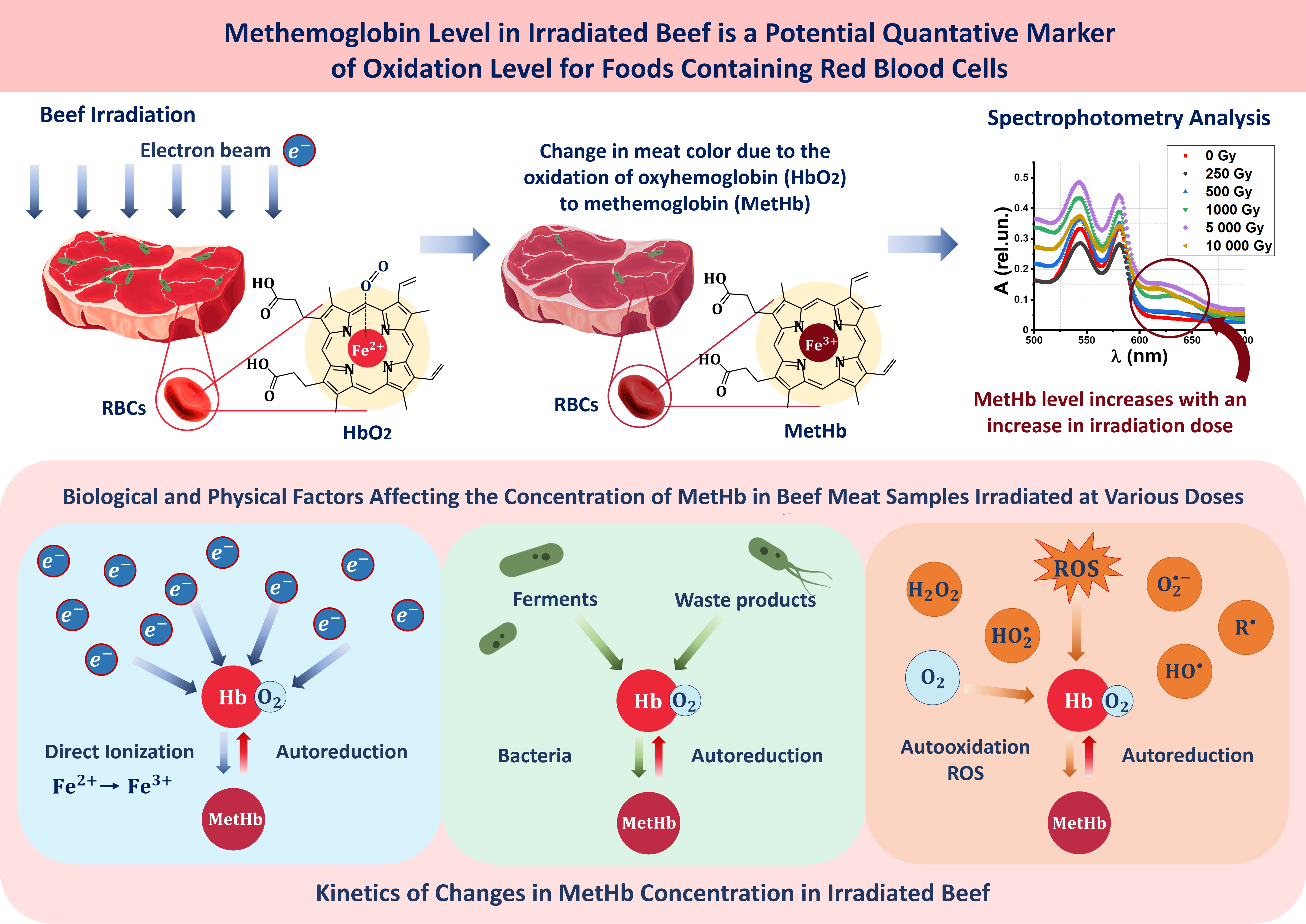
2.1. Research Stages
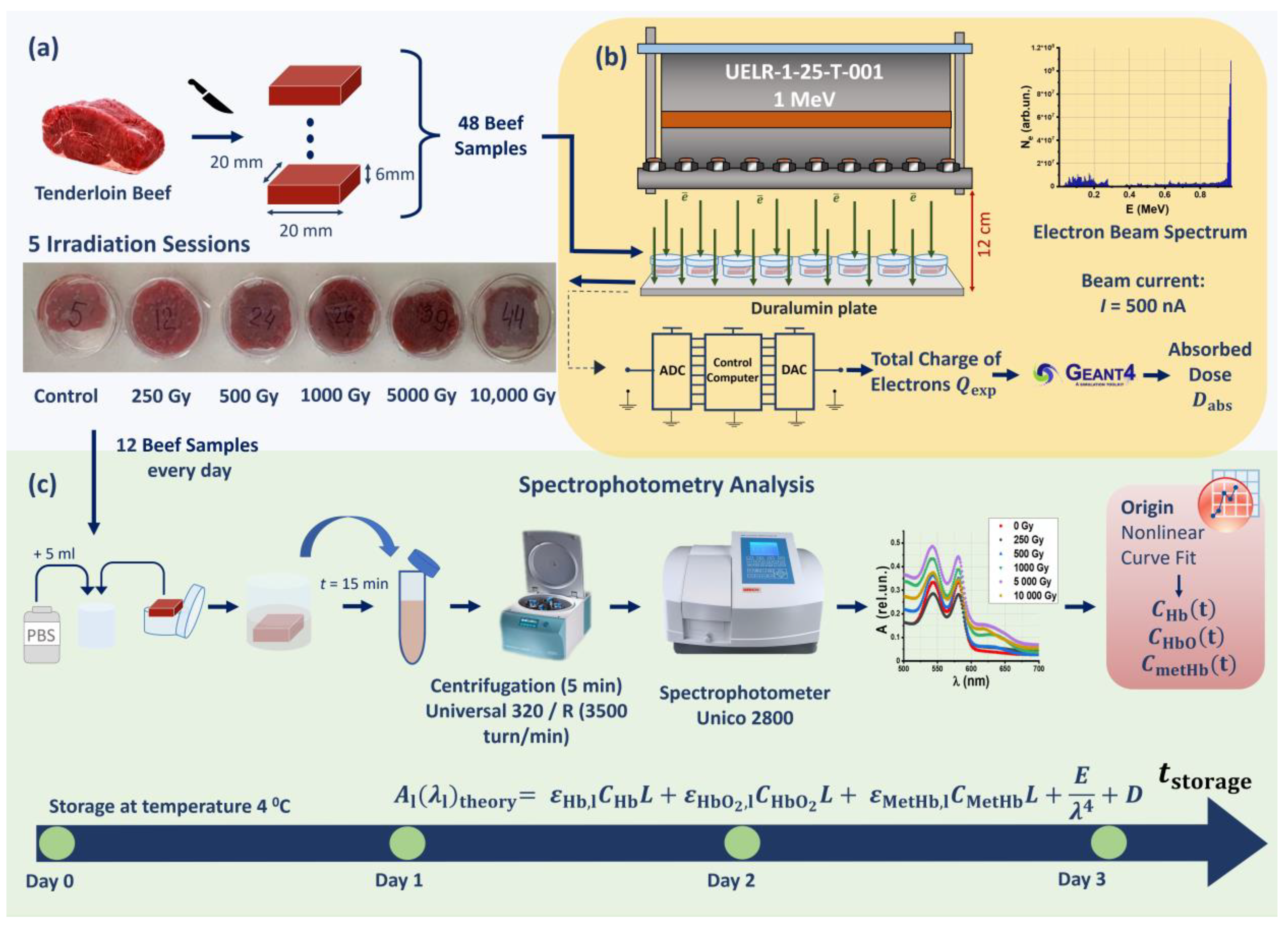
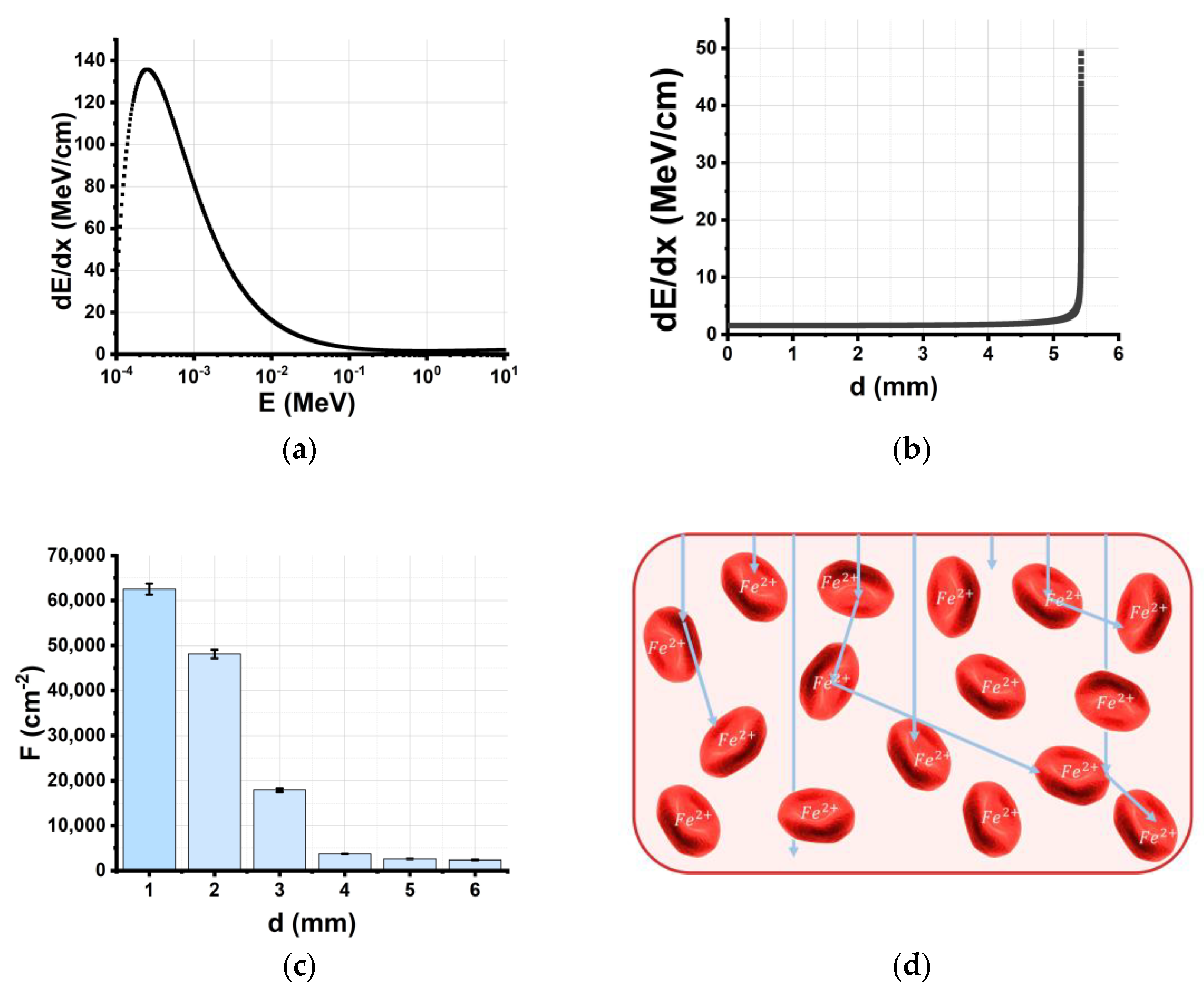
2.2. Nonuniformity of Depth–Dose Distribution across Beef Samples
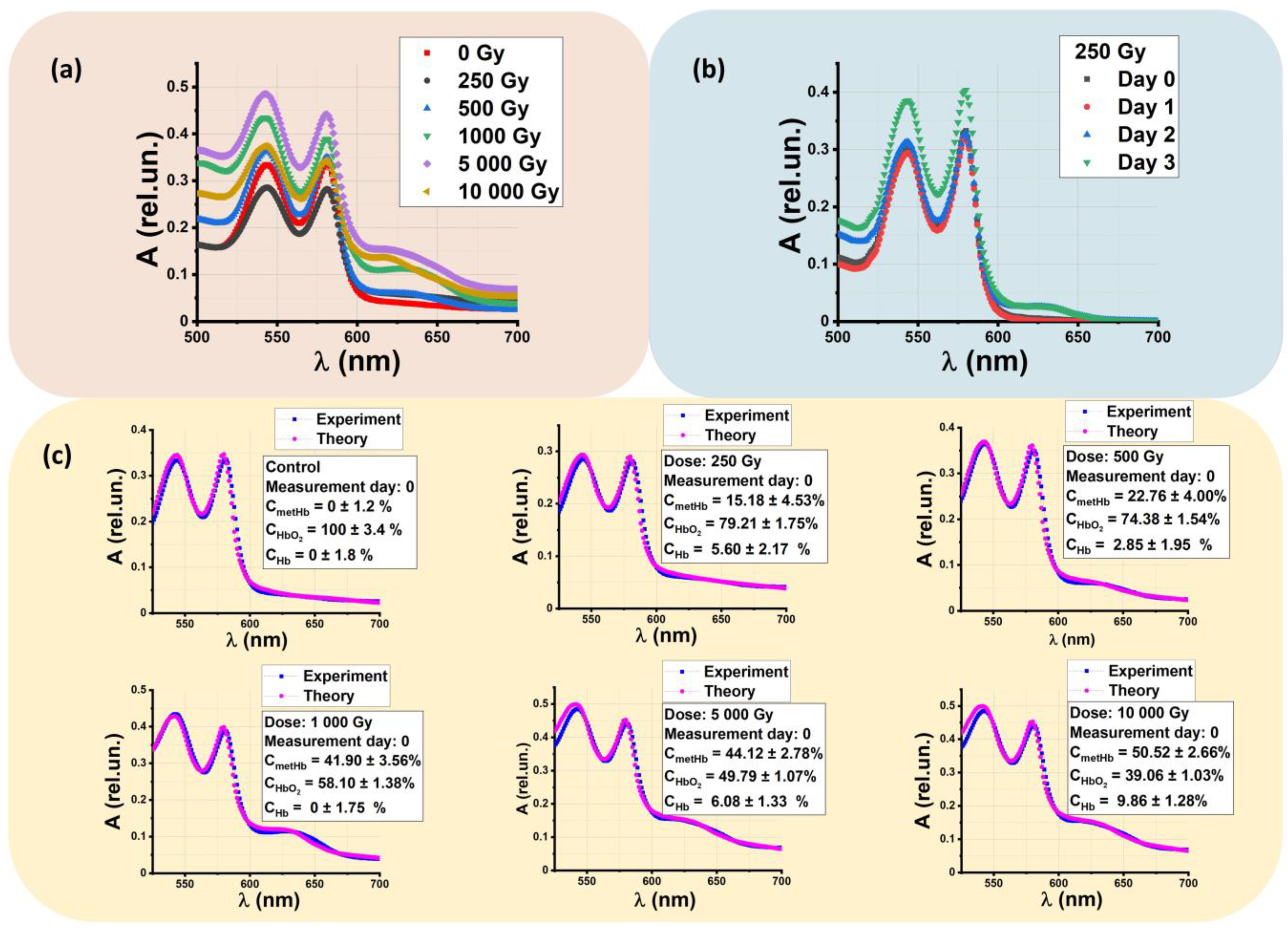
2.3. Concentrations of Hemoglobin Derivatives in Beef Samples after Exposure to Accelerated Electrons
2.3.1. Theoretical Absorption Optical Spectra and Estimation of Hemoglobin Derivative Concentrations in Beef Suspensions
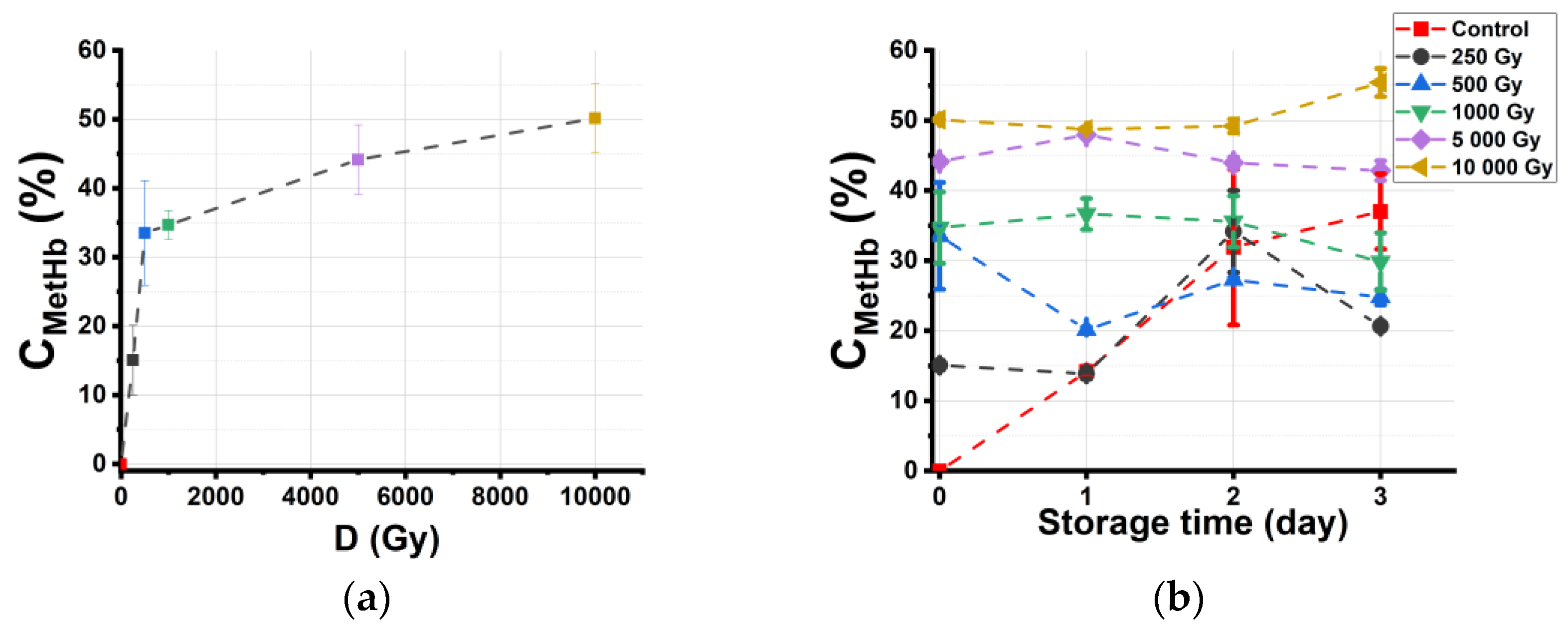
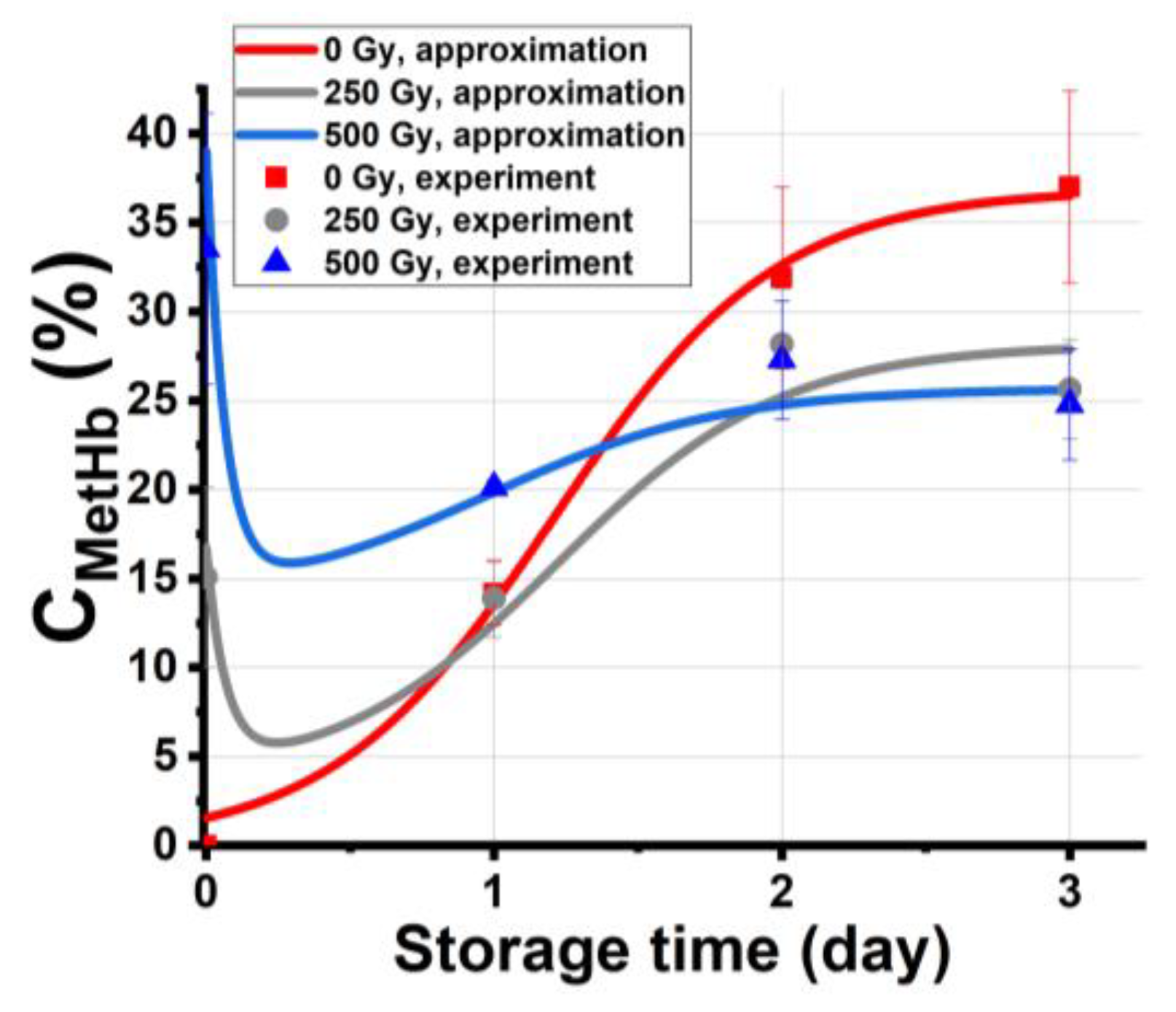
2.3.2. Dependencies of the Hemoglobin Derivative Concentration in Beef on the Irradiation Dose and Storage Time
2.3.3. Effective Dose Range for Beef Irradiation
3. Discussion
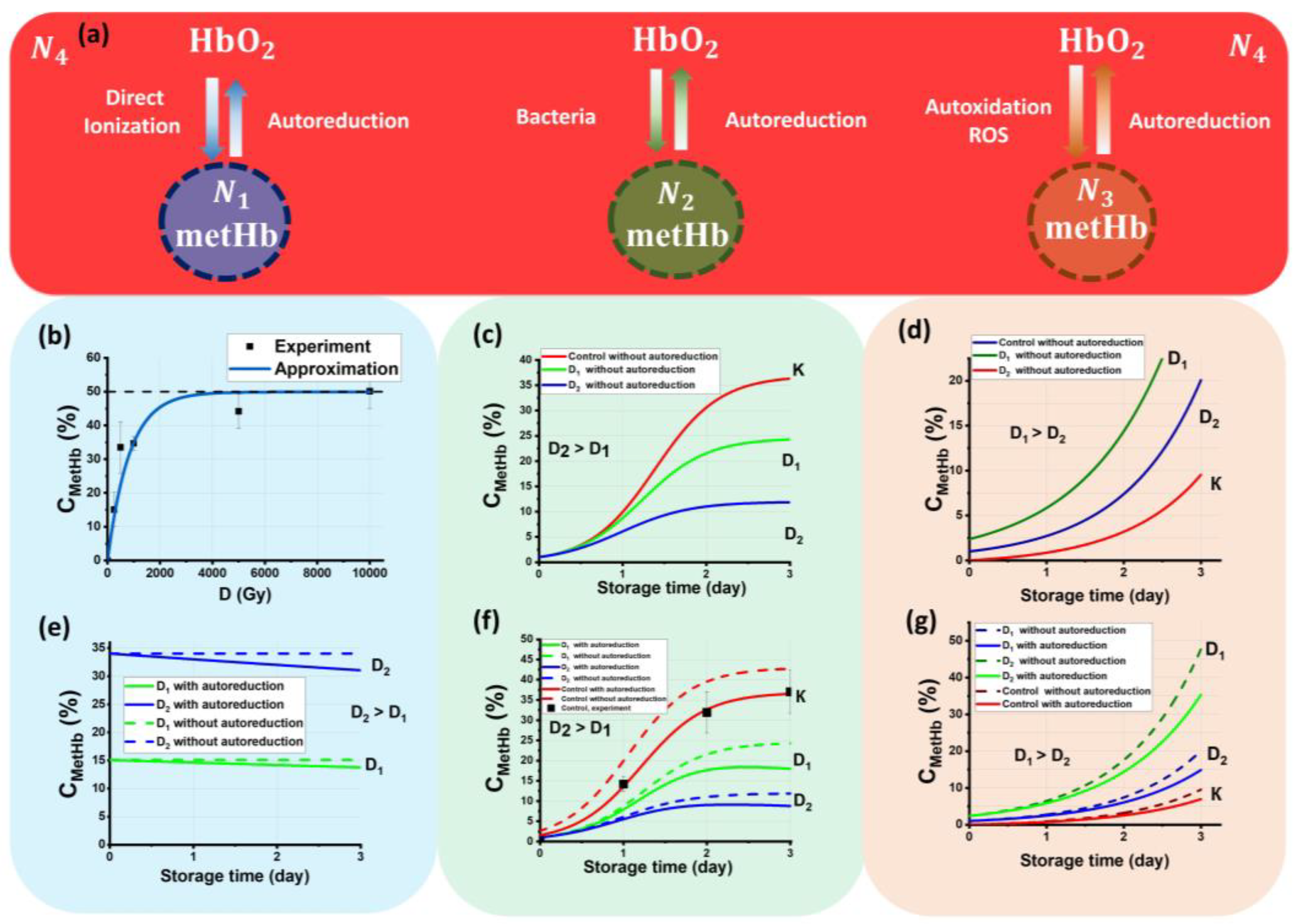
3.1. Reasons for Methemoglobin Concentration Change in Beef Samples during Storage after Electron Beam Irradiation
3.1.1. Direct Ionization of Fe2+ Caused by Accelerated Electrons
3.1.2. Bacterial Activity
3.1.3. Reactive Oxygen Species
3.1.4. Autoreduction of Methemoglobin to Oxyhemoglobin
3.2. Kinetics of Methemoglobin Concentration in Beef Samples after Irradiation
- Ensemble N1 is metHb molecules that appear because of the transformation of ions Fe2+ into Fe3+ as a result of the direct action of accelerated electrons during irradiation;
- Ensemble N2 is metHb molecules that are formed as a result of bacterial activity during the entire period of storage;
- Ensemble N3 is metHb molecules that are formed because of the autoxidation of HbO2 in the presence of ROS present both in nonirradiated and irradiated samples;
- Ensemble N4 is a pool of HbO2 molecules that can transform into metHb molecules because of oxidation and back into HbO2 molecules because of autoreduction during the observation time.
- Each ensemble occurs because of specific factors, such as exposure to the direct action of accelerated electrons, bacterial activity, and autoxidation.
- At the beginning of the observation, t = 0 hemoglobin molecules N in the beef samples are represented only as HbO2 molecules, i.e., N(t = 0) = N4(t = 0) = 100%. As further dynamic transformation of hemoglobin derivatives occurs, the sum of all molecules in four ensembles is always equal to 100%.
- The model assumes that mutual transformations of HbO2 into metHb and back occur during the observation time, and the process of HbO2 transition into deoxyhemoglobin Hb and back is not taken into account. Hb concentration is assumed to be 0% during the whole observation time.
3.2.1. The Kinetics of Ensemble N1
- Nonuniform depth distribution of electron energy losses in the beef sample;
- A decline in the electron beam fluence as electrons penetrate deeper layers of the beef samples;
- Nonuniform depth–dose distribution in the beef samples;
- Discrete character of the interaction of electrons with the matter.
3.2.2. The Kinetics of Ensemble N2
3.2.3. Kinetics of Ensemble N3
3.2.4. Autoreduction of Methemoglobin to Oxyhemoglobin
4. Materials and Methods
4.1. Object of Study
4.2. Electron Beam Irradiation
4.3. Computer Simulation to Determine the Dose Absorbed by Beef Samples
4.4. Spectrophotometry of Hemoglobin Derivatives
4.5. Hemoglobin Derivative Concentration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Global Industry Analysts. Food Irradiation Trends: Global Strategic Business Report; Global Industry Analysts: San Jose, CA, USA, 2023; 196p. [Google Scholar]
- ISO 14470:2011; Food Irradiation. Requirements for the Development, Validation and Routine Control of the Process of Irradiation Using Ionizing Radiation for the Treatment of Food. ISO: Geneva, Switzerland, 2011.
- Pillai, S.D. Introduction to Electron-Beam Food Irradiation Chem. Eng. Progr. 2016, 112, 36–44. [Google Scholar]
- Chulikova, N.; Malyuga, A.; Borshchegovskaya, P.; Zubritskaya, Y.; Ipatova, V.; Chernyaev, A.; Yurov, D.; Zolotov, S.; Nikitchenko, A.; Bliznyuk, U.; et al. Electron Beam Irradiation to Control Rhizoctonia solani in Potato. Agriculture 2023, 13, 1221. [Google Scholar] [CrossRef]
- Singh, R.; Singh, A. Applications of Food Irradiation Technology. Def. Life Sci. J. 2020, 5, 54–62. [Google Scholar] [CrossRef]
- CODEX STAN 106-1983; Rev.1-2003, Codex Alimentarius Commission. General Standard for Irradiated Foods. FAO/WHO: Rome, Italy, 2003.
- Brewer, S. Irradiation effects on meat color—A review. Meat Sci. 2004, 68, 1–17. [Google Scholar] [CrossRef]
- Suman, S.P.; Joseph, P. Myoglobin Chemistry and Meat Color. Annu. Rev. Food Sci. Technol. 2013, 4, 79–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.; Min, B.; Park, K.; Lee, S.; Ahn, D. Effects of Ascorbic Acid and Antioxidants on the Lipid Oxidation and Volatiles of Irradiated Ground Beef. J. Food Sci. 2003, 68, 1680–1685. [Google Scholar] [CrossRef]
- Indiarto, R.; Irawan, A.N.; Subroto, E. Meat Irradiation: A Comprehensive Review of Its Impact on Food Quality and Safety. Foods 2023, 12, 1845. [Google Scholar] [CrossRef]
- Ahn, D.U.; Lee, E.J. Mechanisms and Prevention of Off-Odor Production and Color Changes in Irradiated Meat. Irradiat. Food Packag. ACS Symp. Ser. 2004, 875, 43–76. [Google Scholar] [CrossRef]
- Bliznyuk, U.; Borshchegovskaya, P.; Bolotnik, T.; Chernyaev, A.; Ipatova, V.; Nikitchenko, A.; Shinkarev, O.; Yurov, D.; Khmelevskiy, O.; Rodin, I. Research into Gas Chromatography–Mass Spectrometry (GC-MS) for Ensuring the Effect of 1 MeV-Accelerated Electrons on Volatile Organic Compounds in Turkey Meat. Separations 2022, 9, 227. [Google Scholar] [CrossRef]
- Dave, D.; Ghaly, A.E. Meat Spoilage Mechanisms and Preservation Techniques: A Critical Review. Am. J. Agric. Biol. Sci. 2011, 6, 486–510. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Ahn, D.U. Lipid oxidation and its implications to meat quality and human health. Food Sci. Biotechnol. 2019, 28, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Smit, G.; Smit, B.A.; Engels, W.J. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Palleroni, N.J.; Doudoroff, M. The Aerobic Pseudomonads: A Taxonomic Study. J. Gen. Microbiol. 1996, 43, 159–271. [Google Scholar] [CrossRef] [Green Version]
- Talon, R.; Chastagnac, C.; Vergnais, L.; Montel, M.; Berdagué, J. Production of esters by Staphylococci. Int. J. Food Microbiol. 1998, 45, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F. Proteolysis and Lipolysis in Flavour Development of Dry-cured Meat Products. Meat Sci. 1998, 49, S101–S110. [Google Scholar] [CrossRef]
- Ha, J.-U.; Kim, Y.-M.; Lee, D.-S. Multilayered antimicrobial polyethylene films applied to the packaging of ground beef. Packag. Technol. Sci. 2001, 14, 55–62. [Google Scholar] [CrossRef]
- Gram, L.; Ravn, L.; Rasch, M.; Bruhn, J.B.; Christensen, A.B.; Givskov, M. Food spoilage—Interactions between food spoilage bacteria. Int. J. Food Microbiol. 2002, 78, 79–97. [Google Scholar] [CrossRef]
- Franke, C.; Hilgarth, M.; Vogel, R.F.; Petermeier, H.; Langowski, H.-C. Characterization of the dynamics of volatile organic compounds released by lactic acid bacteria on modified atmosphere packed beef by PTR-MS. Food Packag. Shelf Life 2019, 22, 100400. [Google Scholar] [CrossRef]
- Yushina, Y.K.; Bataeva, D.S.; Zaiko, E.V.; Machova, A.A.; Velebit, B. Bacterial populations and volatile organic compounds associated with meat spoilage. IOP Conf. Ser. Earth Environ. Sci. 2019, 333, 012114. [Google Scholar] [CrossRef] [Green Version]
- Tauro, P.; Kapoor, K.K.; Yadav, K.S. An Introduction to Microbiology, 1st ed.; New Age International Publisher: New Delhi, India, 1986; p. 364. [Google Scholar]
- Kozlova, E.; Chernysh, A.; Sergunova, V.; Gudkova, O.; Manchenko, E.; Kozlov, A. Atomic force microscopy study of red blood cell membrane nanostructure during oxidation-reduction processes. J. Mol. Recognit. 2018, 31, e2724. [Google Scholar] [CrossRef]
- Kozlova, E.; Sherstyukova, E.; Sergunova, V.; Grechko, A.; Kuzovlev, A.; Lyapunova, S.; Inozemtsev, V.; Kozlov, A.; Chernysh, A. Atomic Force Microscopy and High-Resolution Spectrophotometry for Study of Anoxemia and Normoxemia in Model Experiment In Vitro. Int. J. Mol. Sci. 2023, 24, 11043. [Google Scholar] [CrossRef]
- Almac, E.; Bezemer, R.; Hilarius-Stokman, P.M.; Goedhart, P.; de Korte, D.; Verhoeven, A.J.; Ince, C. Red blood cell storage increases hypoxia-induced nitric oxide bioavailability and methemoglobin formation in vitro and in vivo. Transfusion 2014, 54, 3178–3185. [Google Scholar] [CrossRef]
- Studenikin, F.R.; Bliznyuk, U.A.; Chernyaev, A.P.; Krusanov, G.A.; Nikitchenko, A.D.; Zolotov, S.A.; Ipatova, V.S. Electron beam modification for improving dose uniformity in irradiated objects. Eur. Phys. J. Spec. Top. 2023, 1–5. [Google Scholar] [CrossRef]
- Bliznyuk, U.; Avdyukhina, V.; Borshchegovskaya, P.; Bolotnik, T.; Ipatova, V.; Nikitina, Z.; Nikitchenko, A.; Rodin, I.; Studenikin, F.; Chernyaev, A.; et al. Effect of electron and X-ray irradiation on microbiological and chemical parameters of chilled turkey. Sci. Rep. 2022, 12, 750. [Google Scholar] [CrossRef]
- Voeikov, V. Reactive oxygen species, water, photons and life. Riv. Biol. Biol. Forum 2010, 103, 321–342. [Google Scholar]
- Kudryashov, Y.B. Radiation Biophysics (Ionizing Radiations); Nova Science Publishers: Hauppauge, NY, USA, 2013; 362p. [Google Scholar]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Yang, P.; Hong, L.; Cai, X.; Zeng, Z.; Wu, X.; Zeng, H.; Shao, L. Oxidative Stress in Radiation-Induced Cardiotoxicity. Oxidative Med. Cell. Longev. 2020, 2020, 3579143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalafalla, G.M.; Nasr, N.F.; Gaafar, A.M.; Abo-Zaid, R.M. Effect of gamma irradiation on microbial load, physicochemical characteristicsand shelf-life of raw minced beef meat Middle East. J. Appl. Sci. 2018, 8, 625–634. [Google Scholar]
- Bhoir, S.A.; Jhaveri, M.; Chawla, S.P. Evaluation and predictive modeling of the effect of chitosan and gamma irradiation on quality of stored chilled chicken meat. J. Food Process. Eng. 2019, 42, e13254. [Google Scholar] [CrossRef]
- Schopf, S.; Gotzmann, G.; Dietze, M.; Gerschke, S.; Kenner, L.; König, U. Investigations Into the Suitability of Bacterial Suspensions as Biological Indicators for Low-Energy Electron Irradiation. Front. Immunol. 2022, 13, 814767. [Google Scholar] [CrossRef]
- Atolia, E.; Cesar, S.; Arjes, H.A.; Rajendram, M.; Shi, H.; Knapp, B.D.; Khare, S.; Aranda-Díaz, A.; Lenski, R.E.; Huang, K.C. Environmental and Physiological Factors Affecting High-Throughput Measurements of Bacterial Growth. mBio 2020, 11, e01378-20. [Google Scholar] [CrossRef] [PubMed]
- Tilak, K.S.; Veeraiah, K.; Raju, J.M.P. Effects of ammonia, nitrite and nitrate on hemoglobin content and oxygen consumption of freshwater fish, Cyprinus carpio (Linnaeus). J. Environ. Biol. 2007, 28, 45–47. [Google Scholar]
- Faustman, C.; Cassens, R. The Biochemical Basis for Discoloration in Fresh Meat: A Review. J. Muscle Foods 1990, 1, 217–243. [Google Scholar] [CrossRef]
- Kuhn, V.; Diederich, L.; Keller, T.C.S.; Kramer, C.M.; Lückstädt, W.; Panknin, C.; Suvorava, T.; Isakson, B.E.; Kelm, M.; Cortese-Krott, M.M. Red Blood Cell Function and Dysfunction: Redox Regulation, Nitric Oxide Metabolism, Anemia. Antioxid. Redox Signal. 2017, 26, 718–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlova, E.; Sherstyukova, E.; Sergunova, V.; Kozlov, A.; Gudkova, O.; Inozemtsev, V.; Chernysh, A. The Toxic Influence of Excess Free Iron on Red Blood Cells in the Biophysical Experiment: An In Vitro Study. J. Toxicol. 2022, 2022, 7113958. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, E.; Sergunova, V.; Sherstyukova, E.; Grechko, A.; Lyapunova, S.; Inozemtsev, V.; Kozlov, A.; Gudkova, O.; Chernysh, A. Mechanochemical Synergism of Reactive Oxygen Species Influences on RBC Membrane. Int. J. Mol. Sci. 2023, 24, 5952. [Google Scholar] [CrossRef]
- Macho-González, A.; Garcimartín, A.; López-Oliva, M.E.; Bastida, S.; Benedí, J.; Ros, G.; Nieto, G.; Sánchez-Muniz, F.J. Can Meat and Meat-Products Induce Oxidative Stress? Antioxidants 2020, 9, 638. [Google Scholar] [CrossRef]
- Wang, D.; Piknova, B.; Solomon, S.B.; Cortes-Puch, I.; Kern, S.J.; Sun, J.; Kanias, T.; Gladwin, M.T.; Helms, C.; Kim-Shapiro, D.B.; et al. In vivo reduction of cell-free methemoglobin to oxyhemoglobin results in vasoconstriction in canines. Transfusion 2013, 53, 3149–3163. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, A.; Nakayama, Y.; Kitayama, T.; Tomita, M. Simulation study of methemoglobin reduction in erythrocytes. FEBS J. 2007, 274, 1449–1458. [Google Scholar] [CrossRef]
- Hillen, T. Applications and Limitations of the Verhulst Model for Populations. Math Biol. 2003, 6, 19–20. [Google Scholar]
- Brown, J.; Yates, E.; Bielecki, M.; Olczak, T.; Smalley, J. Potential role for Streptococcus gordonii-derived hydrogen peroxide in heme acquisition by Porphyromonas gingivalis. Mol. Oral Microbiol. 2018, 33, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Rifkind, J.M.; Nagababu, E.; Ramasamy, S.; Ravi, L.B. Hemoglobin redox reactions and oxidative stress. Redox Rep. 2003, 8, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, W.; Buursma, A. Spectrophotometry of Hemoglobin: Absorption Spectra of Bovine Oxyhemoglobin, Deoxyhemoglobin, Carboxyhemoglobin, and Methemoglobin. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1997, 118, 743–749. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irradiation Session | Dexp (Gy) | texp (s) | Qexp (nC) | D (Gy) |
|---|---|---|---|---|
| 1, 2 | 248 ± 6 | 55 ± 1 | 16,310 ± 285 | 250 |
| 3, 4 | 502 ± 10 | 100 ± 1 | 32,700 ± 423 | 500 |
| 5, 6 | 1018 ± 20 | 186 ± 1 | 65,058 ± 1112 | 1000 |
| 7, 8 | 5023 ± 50 | 988 ± 1 | 325,165 ± 5870 | 5000 |
| 9, 10 | 10,037 ± 90 | 1864 ± 1 | 650,108 ± 8860 | 10,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bliznyuk, U.; Borshchegovskaya, P.; Chernyaev, A.; Ipatova, V.; Kozlov, A.; Khmelevskiy, O.; Mezhetova, I.; Nikitchenko, A.; Rodin, I.; Kozlova, E. Hemoglobin Derivatives in Beef Irradiated with Accelerated Electrons. Molecules 2023, 28, 5773. https://doi.org/10.3390/molecules28155773
Bliznyuk U, Borshchegovskaya P, Chernyaev A, Ipatova V, Kozlov A, Khmelevskiy O, Mezhetova I, Nikitchenko A, Rodin I, Kozlova E. Hemoglobin Derivatives in Beef Irradiated with Accelerated Electrons. Molecules. 2023; 28(15):5773. https://doi.org/10.3390/molecules28155773
Chicago/Turabian StyleBliznyuk, Ulyana, Polina Borshchegovskaya, Alexander Chernyaev, Victoria Ipatova, Aleksandr Kozlov, Oleg Khmelevskiy, Irina Mezhetova, Alexander Nikitchenko, Igor Rodin, and Elena Kozlova. 2023. "Hemoglobin Derivatives in Beef Irradiated with Accelerated Electrons" Molecules 28, no. 15: 5773. https://doi.org/10.3390/molecules28155773