
Quercitrin Is a Novel Inhibitor of Salmonella enterica Serovar Typhimurium Type III Secretion System
 and
and 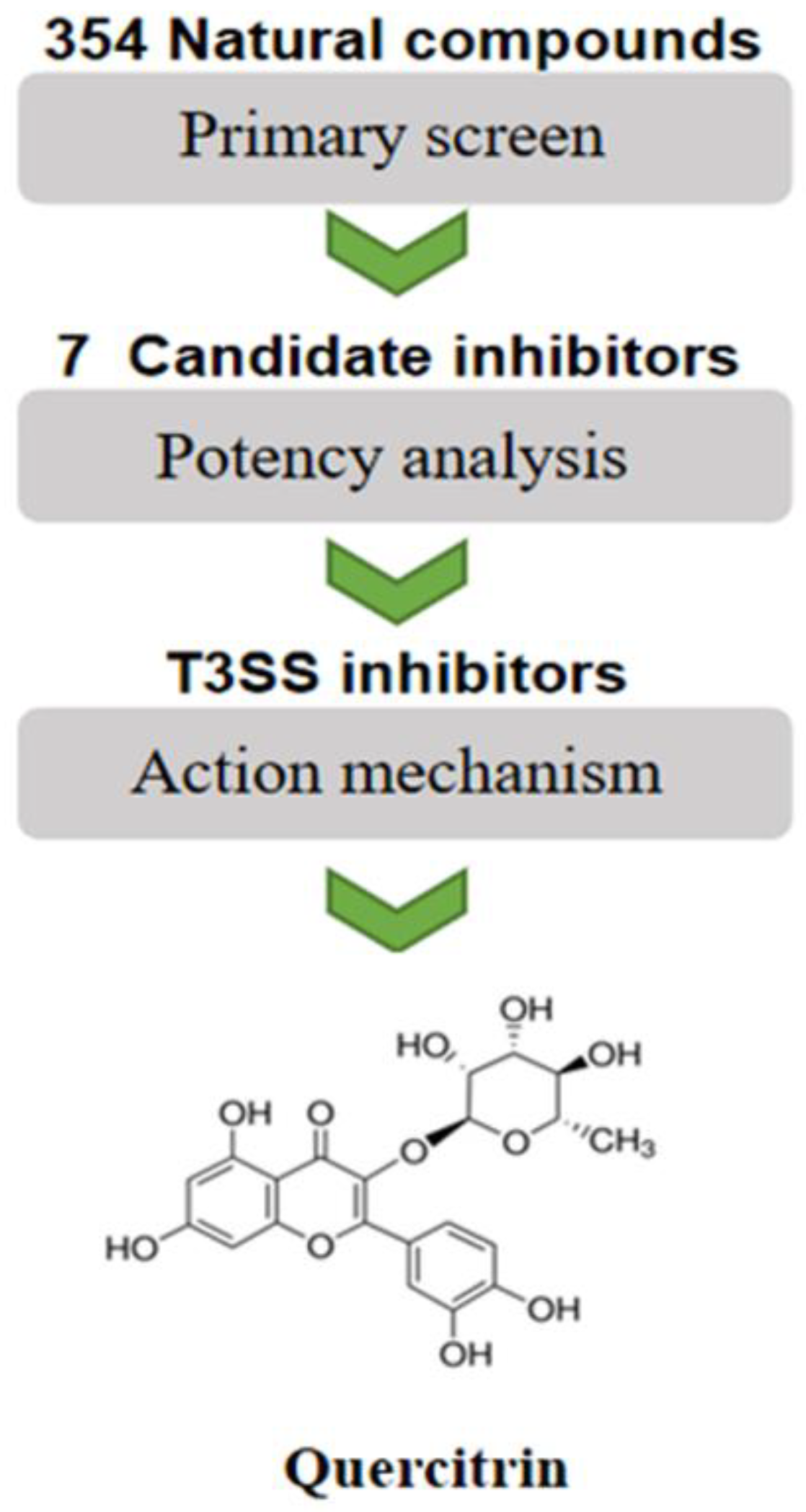{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Screening Natural Compound Inhibitors of S. Typhimurium T3SS
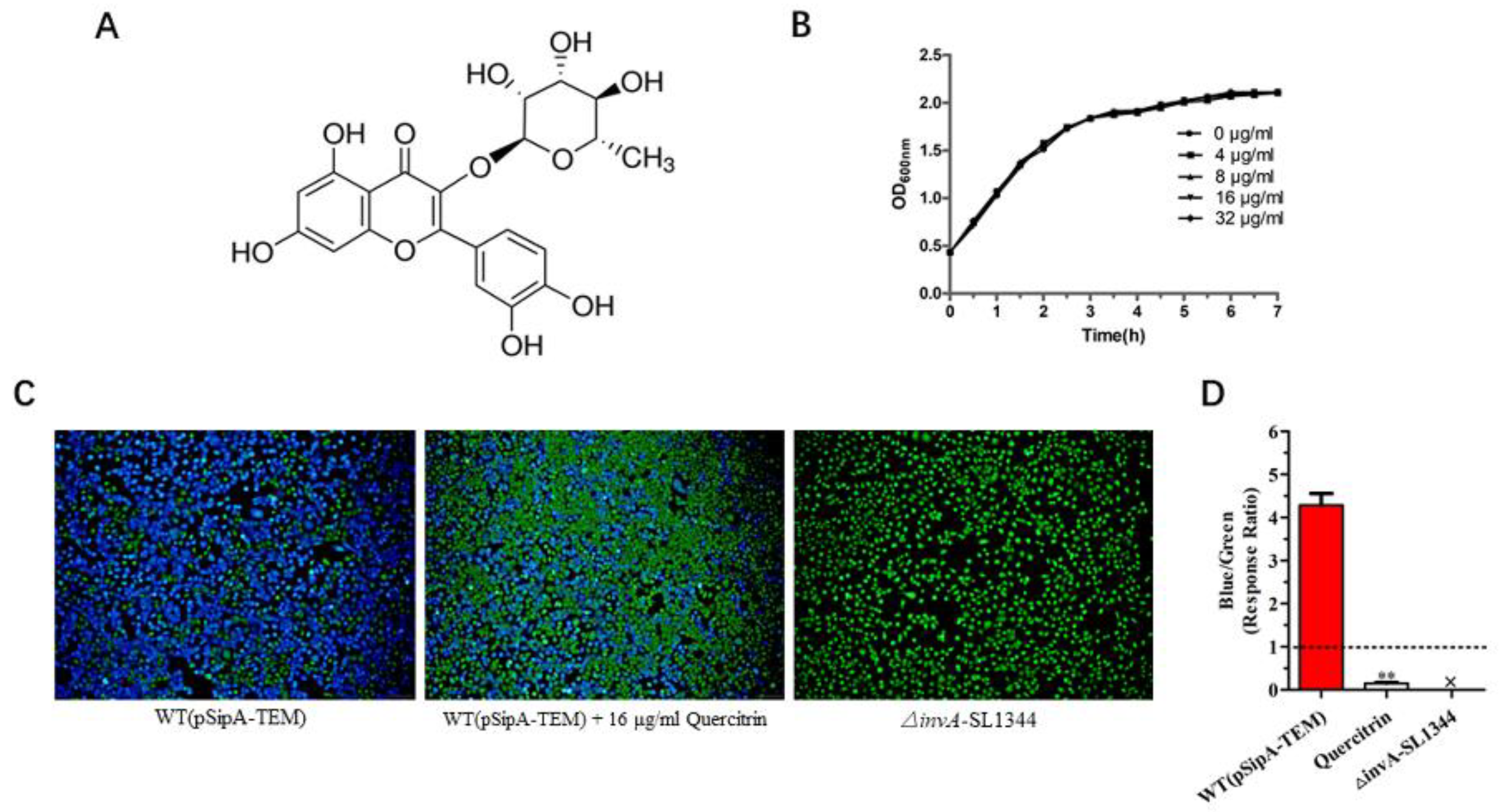
2.2. Quercitrin Blocks the Translocation of T3SS Effector SipA
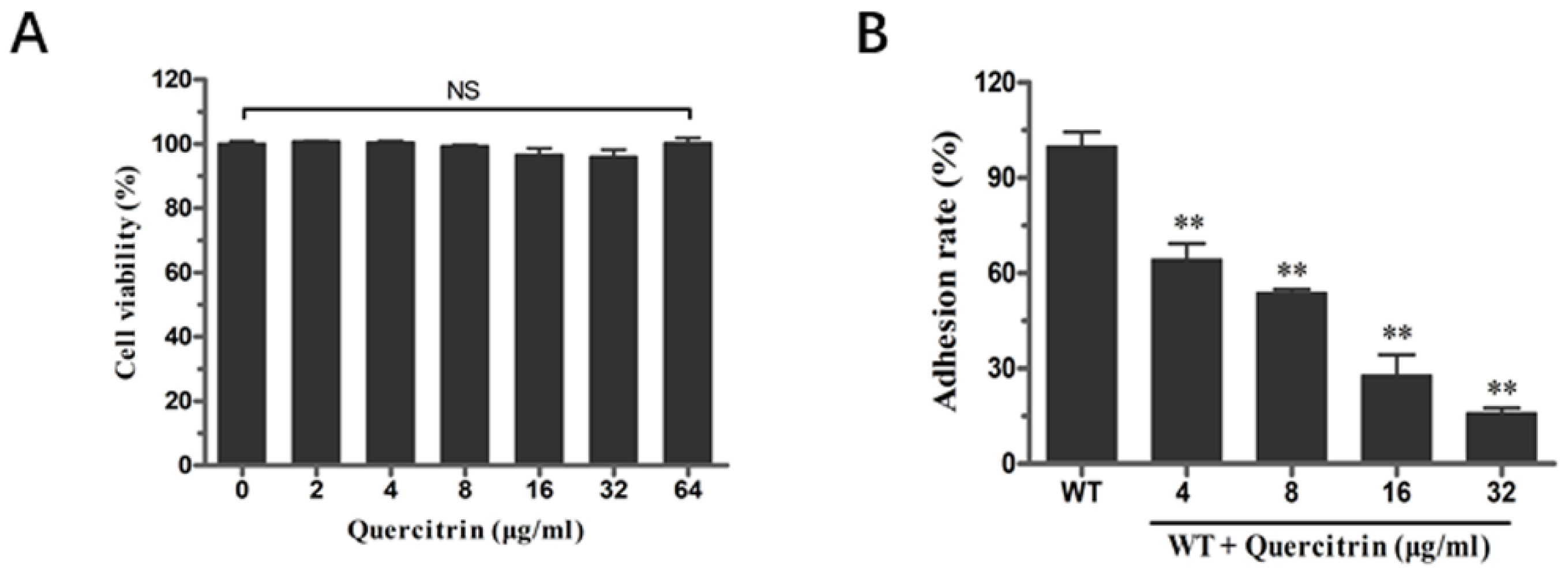
2.3. Quercitrin Inhibits S. Typhimurium Adhesion in a Dose-Dependent Manner
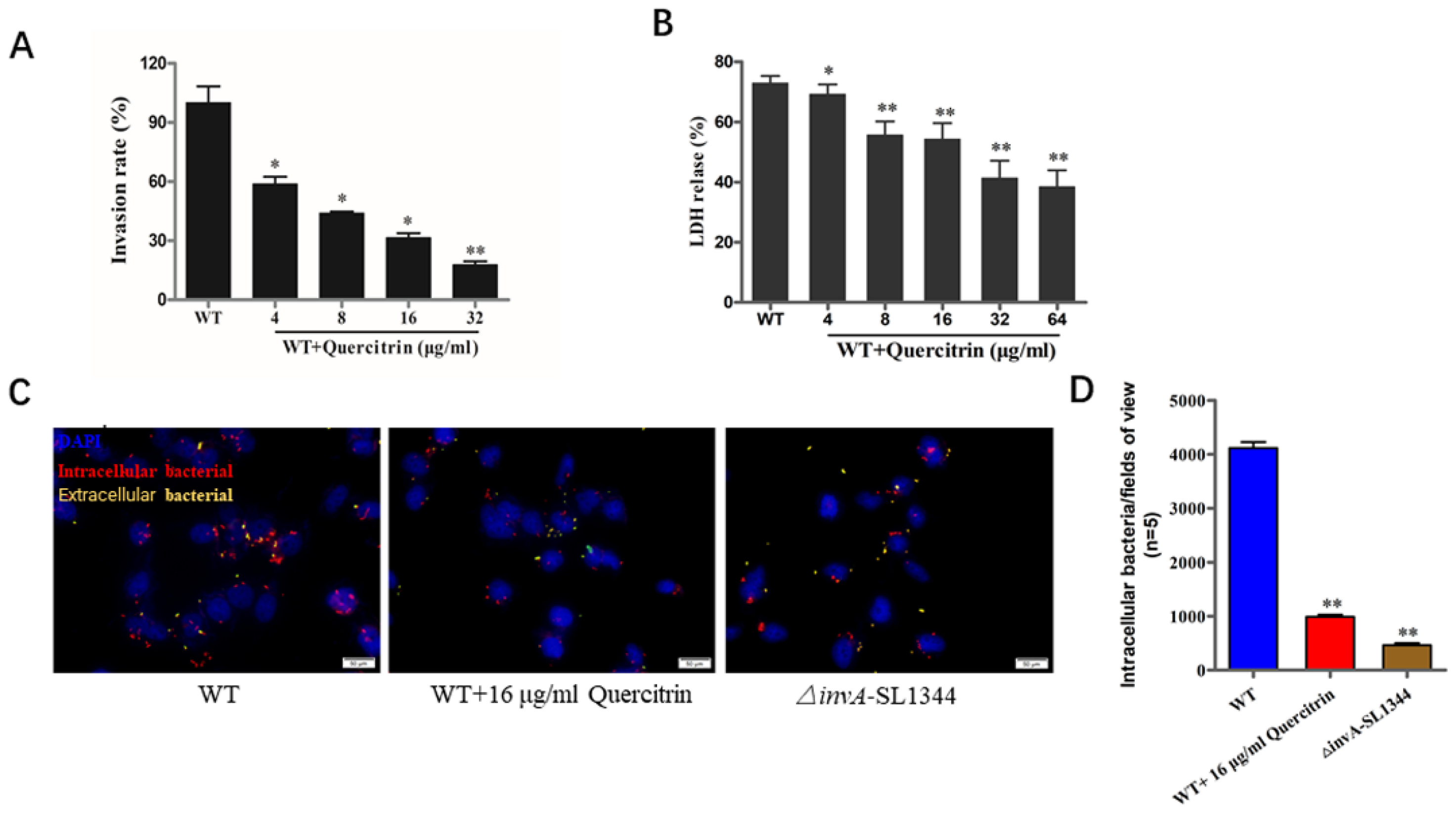
2.4. Quercitrin Inhibits S. Typhimurium Invasion in a Dose-Dependent Manner
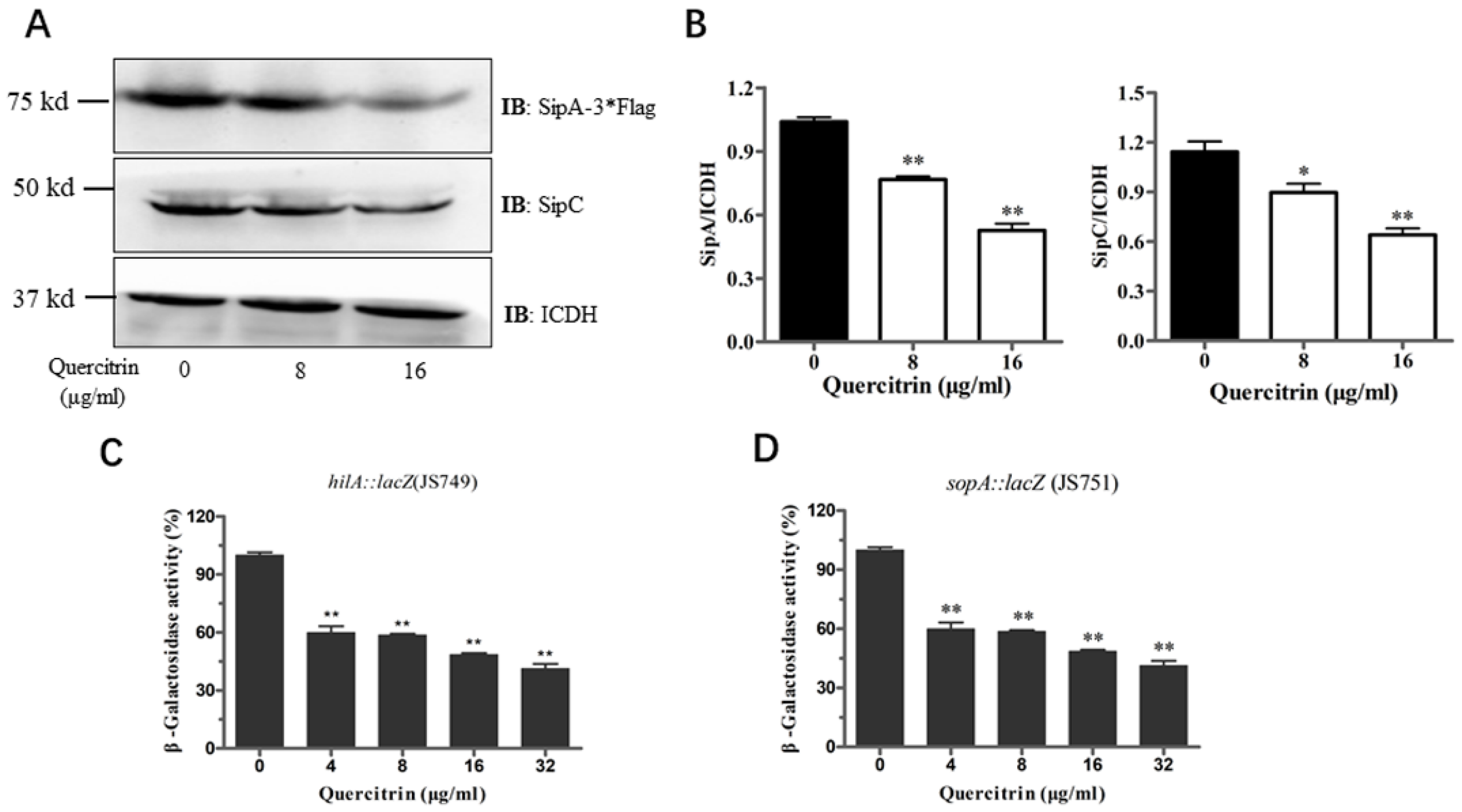
2.5. Quercitrin Inhibits Expression of Related Genes and Effectors in T3SS
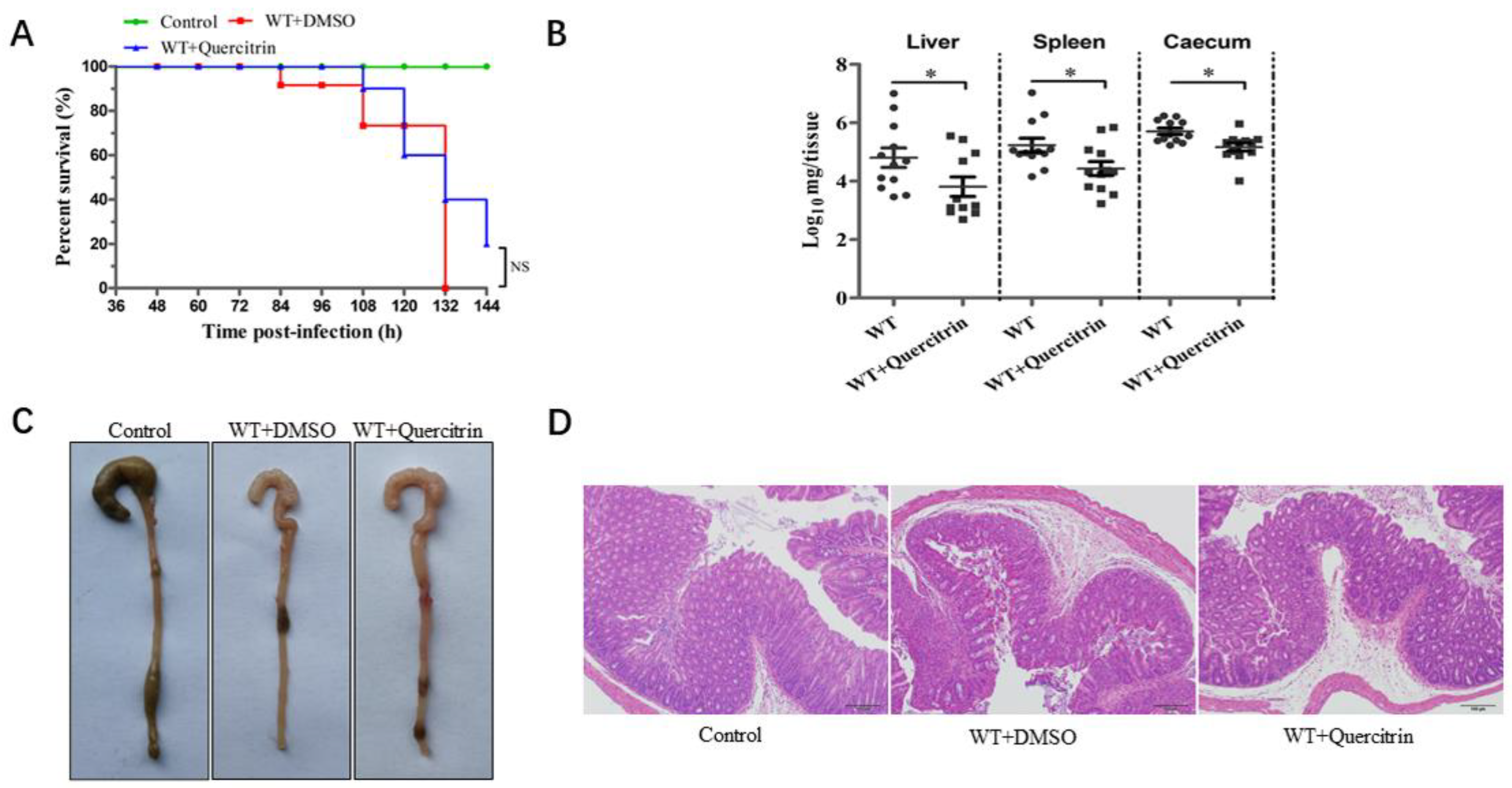
2.6. Quercitrin Improves Cecum Morphology and Reduces the Organ Colonization of Mice after S. Typhimurium Challenge
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Growth Conditions, and Natural Compounds
4.2. High-Throughput Screening for T3SS Inhibitors Using SipA-β-lactamase Fusion Reporter System
4.3. Determination of Bacterial Viability following Quercitrin Exposure
4.4. Cytotoxicity of Quercitrin Indicated by Lactate Dehydrogenase Release
4.5. Adherence Assay
4.6. Gentamicin Protection Assay and Immunofluorescence Staining
4.7. Antibodies and Western Blot
4.8. β-Galactosidase Activity Assay
4.9. Animal Experiment
4.10. Statistics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Knodler, L.A.; Elfenbein, J.R. Salmonella enterica. Trends. Microbiol. 2019, 27, 964–965. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Mylona, E.; Frankel, G. Typhoidal Salmonella: Distinctive virulence factors and pathogenesis. Cell. Microbiol. 2018, 20, e12939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deiwick, J.; Salcedo, S.P.; Boucrot, E.; Gilliland, S.M.; Henry, T.; Petermann, N.; Waterman, S.R.; Gorvel, J.-P.; Holden, D.W.; Méresse, S. The translocated Salmonella effector proteins SseF and SseG interact and are required to establish an intracellular replication niche. Infect. Immun. 2006, 74, 6965–6972. [Google Scholar] [CrossRef] [Green Version]
- Threlfall, E.J. Antimicrobial drug resistance in Salmonella: Problems and perspectives in food- and water-borne infections. FEMS Microbiol. Rev. 2002, 26, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Galán, J.E.; Curtiss, R. Cloning and molecular characterization of genes whose products allow Salmonella typhimurium to penetrate tissue culture cells. Proc. Natl. Acad. Sci. USA 1989, 86, 6383–6387. [Google Scholar] [CrossRef]
- Wallis, T.S.; Galyov, E.E. Molecular basis of Salmonella-induced enteritis. Mol. Microbiol. 2000, 36, 997–1005. [Google Scholar] [CrossRef]
- Lostroh, C.P.; Lee, C.A. The Salmonella pathogenicity island-1 type III secretion system. Microbes Infect. 2001, 3, 1281–1291. [Google Scholar] [CrossRef]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Mooseker, M.S.; Galán, J.E. An invasion-associated Salmonella protein modulates the actin-bundling activity of plastin. Proc. Natl. Acad. Sci. USA 1999, 96, 10176–10181. [Google Scholar] [CrossRef]
- Hersh, D.; Monack, D.M.; Smith, M.R.; Ghori, N.; Falkow, S.; Zychlinsky, A. The Salmonella invasin SipB induces macrophage apoptosis by binding to caspase-1. Proc. Natl. Acad. Sci. USA 1999, 96, 2396–2401. [Google Scholar] [CrossRef]
- Kaniga, K.; Tucker, S.; Trollinger, D.; Galán, J.E. Homologs of the Shigella IpaB and IpaC invasins are required for Salmonella typhimurium entry into cultured epithelial cells. J. Bacteriol. 1995, 177, 3965–3971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, Y.; Qiu, J.; Luo, Z.-Q.; Deng, X. The Herbal Compound Thymol Protects Mice from Lethal Infection by Typhimurium. Front. Microbiol. 2018, 9, 1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Diard, M.; Stecher, B.; Hardt, W.-D. The streptomycin mouse model for Salmonella diarrhea: Functional analysis of the microbiota, the pathogen’s virulence factors, and the host’s mucosal immune response. Immunol. Rev. 2012, 245, 56–83. [Google Scholar] [CrossRef] [PubMed]
- Tournier, I.; Bernuau, D.; Poliard, A.; Schoevaert, D.; Feldmann, G. Detection of albumin mRNAs in rat liver by in situ hybridization: Usefulness of paraffin embedding and comparison of various fixation procedures. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1987, 35, 453–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinaud, L.; Sansonetti, P.J.; Phalipon, A. Host Cell Targeting by Enteropathogenic Bacteria T3SS Effectors. Trends. Microbiol. 2018, 26, 266–283. [Google Scholar] [CrossRef]
- Negrea, A.; Bjur, E.; Ygberg, S.E.; Elofsson, M.; Wolf-Watz, H.; Rhen, M. Salicylidene acylhydrazides that affect type III protein secretion in Salmonella enterica serovar typhimurium. Antimicrob. Agents. Chemother. 2007, 51, 2867–2876. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Li, X.; Li, J.; Yang, X.; Zhou, Y.; Lu, C.; Shen, Y. Licoflavonol is an inhibitor of the type three secretion system of Salmonella enterica serovar Typhimurium. Biochem. Biophys. Res. Commun. 2016, 477, 998–1004. [Google Scholar] [CrossRef]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug. Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Hussain, S.; Ouyang, P.; Zhu, Y.; Khalique, A.; He, C.; Liang, X.; Shu, G.; Yin, L. Type 3 secretion system 1 of Salmonella typhimurium and its inhibitors: A novel strategy to combat salmonellosis. Environ. Sci. Pollut. Res. Int. 2021, 28, 34154–34166. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, A.; Manzanaro-Moreno, N.; Colom, C.; Rønold, H.J.; Lyngstadaas, S.P.; Monjo, M.; Ramis, J.M. Quercitrin Nanocoated Implant Surfaces Reduce Osteoclast Activity In Vitro and In Vivo. Int. J. Mol. Sci. 2018, 19, 3319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worrall, L.J.; Vuckovic, M.; Strynadka, N.C.J. Crystal structure of the C-terminal domain of the Salmonella type III secretion system export apparatus protein InvA. Protein. Sci. Publ. Protein. Soc. 2010, 19, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.-Q.; Isberg, R.R. Multiple substrates of the Legionella pneumophila Dot/Icm system identified by interbacterial protein transfer. Proc. Natl. Acad. Sci. USA 2004, 101, 841–846. [Google Scholar] [CrossRef]
- Smedowski, A.; Pietrucha-Dutczak, M.; Maniar, R.; Ajeleti, M.; Matuszek, I.; Lewin-Kowalik, J. FluoroGold-Labeled Organotypic Retinal Explant Culture for Neurotoxicity Screening StudieS. Oxidative. Med. Cell. Longev. 2018, 2018, 2487473. [Google Scholar] [CrossRef] [PubMed]
- Tsou, L.K.; Lara-Tejero, M.; RoseFigura, J.; Zhang, Z.J.; Wang, Y.-C.; Yount, J.S.; Lefebre, M.; Dossa, P.D.; Kato, J.; Guan, F.; et al. Antibacterial Flavonoids from Medicinal Plants Covalently Inactivate Type III Protein Secretion Substrates. J. Am. Chem. Soc. 2016, 138, 2209–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, Y.; Luo, J.; Jie, J.; Deng, X.; Song, L. The Herbal Compound Thymol Targets Multiple Salmonella Typhimurium Virulence Factors for Lon Protease Degradation. Front. Front. Pharmacol. 2021, 12, 674955. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Wang, L.; Xu, J.; Liu, S.; Song, Z.; Chen, T.; Deng, X.; Wang, J.; Lv, Q. Quercitrin Is a Novel Inhibitor of Salmonella enterica Serovar Typhimurium Type III Secretion System. Molecules 2023, 28, 5455. https://doi.org/10.3390/molecules28145455
Li Q, Wang L, Xu J, Liu S, Song Z, Chen T, Deng X, Wang J, Lv Q. Quercitrin Is a Novel Inhibitor of Salmonella enterica Serovar Typhimurium Type III Secretion System. Molecules. 2023; 28(14):5455. https://doi.org/10.3390/molecules28145455
Chicago/Turabian StyleLi, Qingjie, Lianping Wang, Jingwen Xu, Shuang Liu, Zeyu Song, Tingting Chen, Xuming Deng, Jianfeng Wang, and Qianghua Lv. 2023. "Quercitrin Is a Novel Inhibitor of Salmonella enterica Serovar Typhimurium Type III Secretion System" Molecules 28, no. 14: 5455. https://doi.org/10.3390/molecules28145455