
Identification of Anti-Inflammatory Compounds from Peucedanum praeruptorum Roots by Using Nitric Oxide-Producing Rat Hepatocytes Stimulated by Interleukin 1β
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
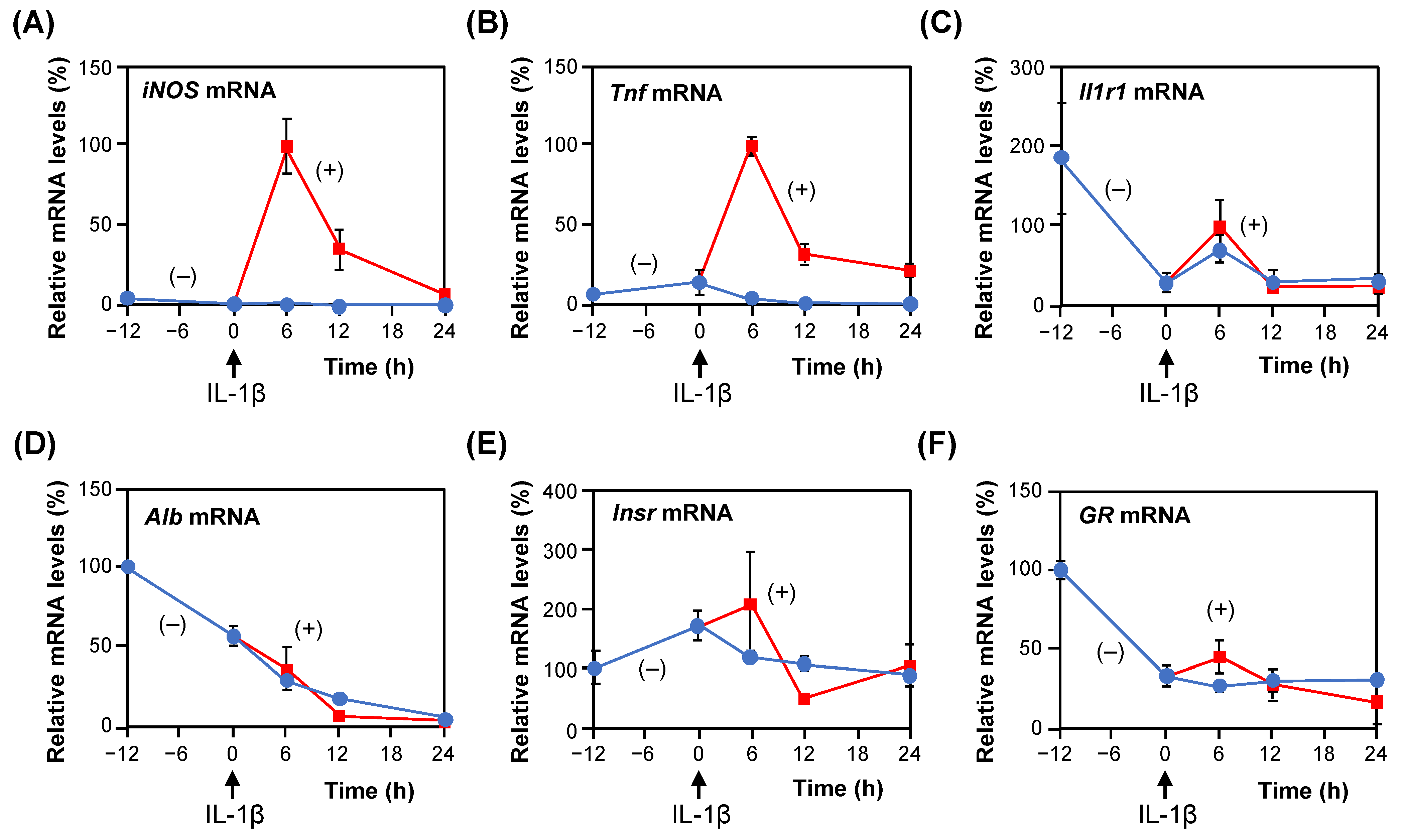
2.1. Gene Expression in the Presence of IL-1β in Rat Hepatocytes
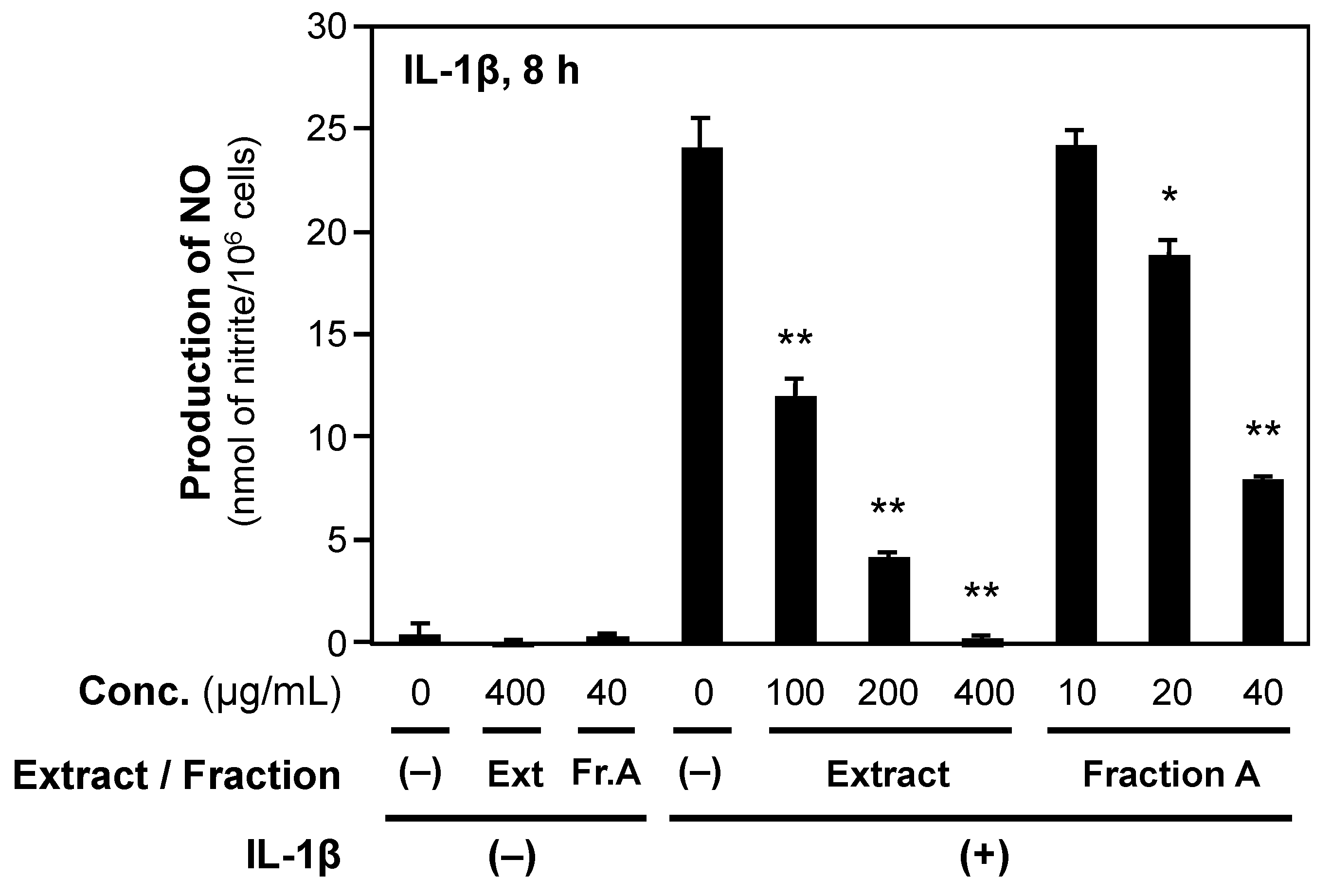
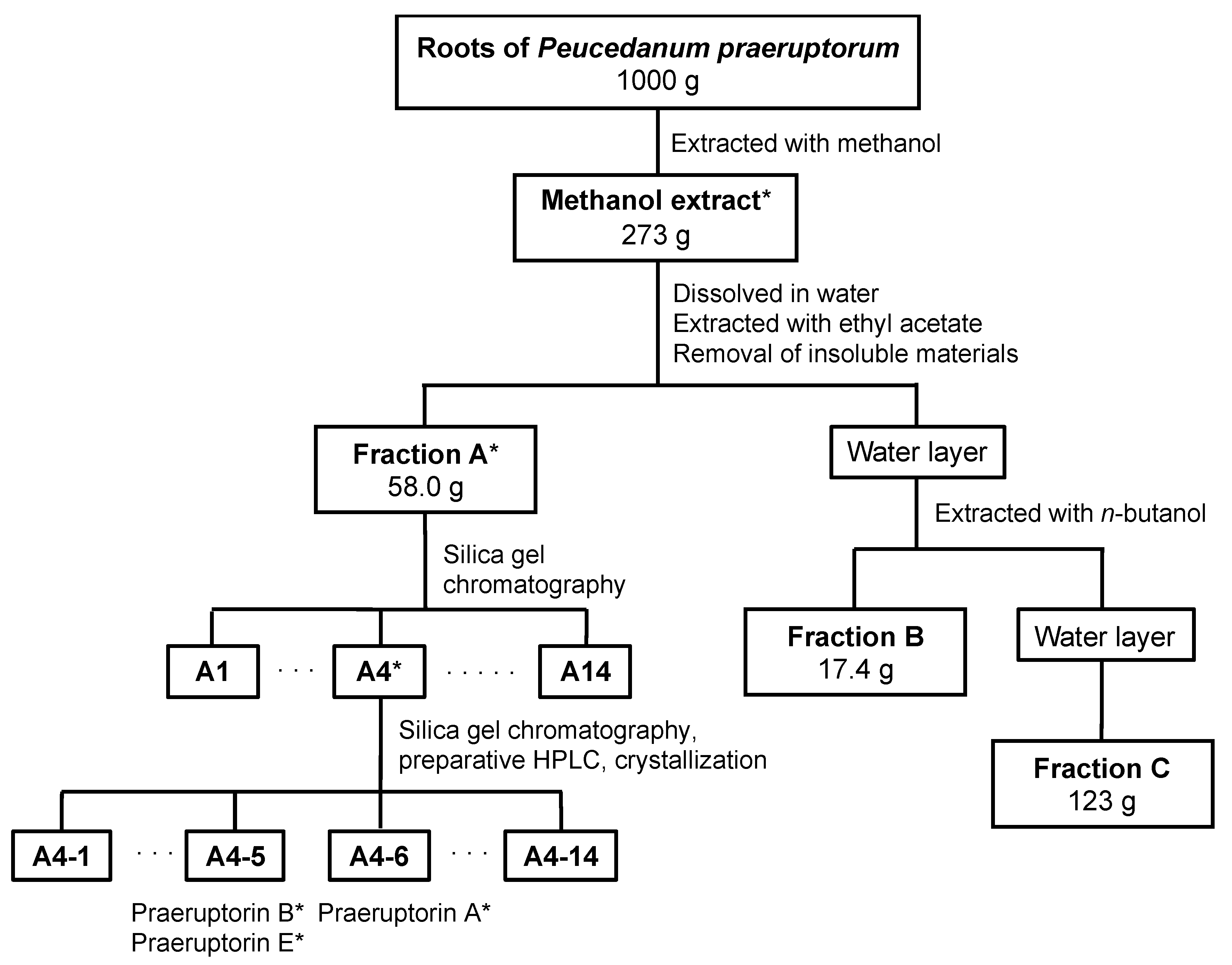
2.2. Crude Fractionation of P. praeruptorum Root Extract
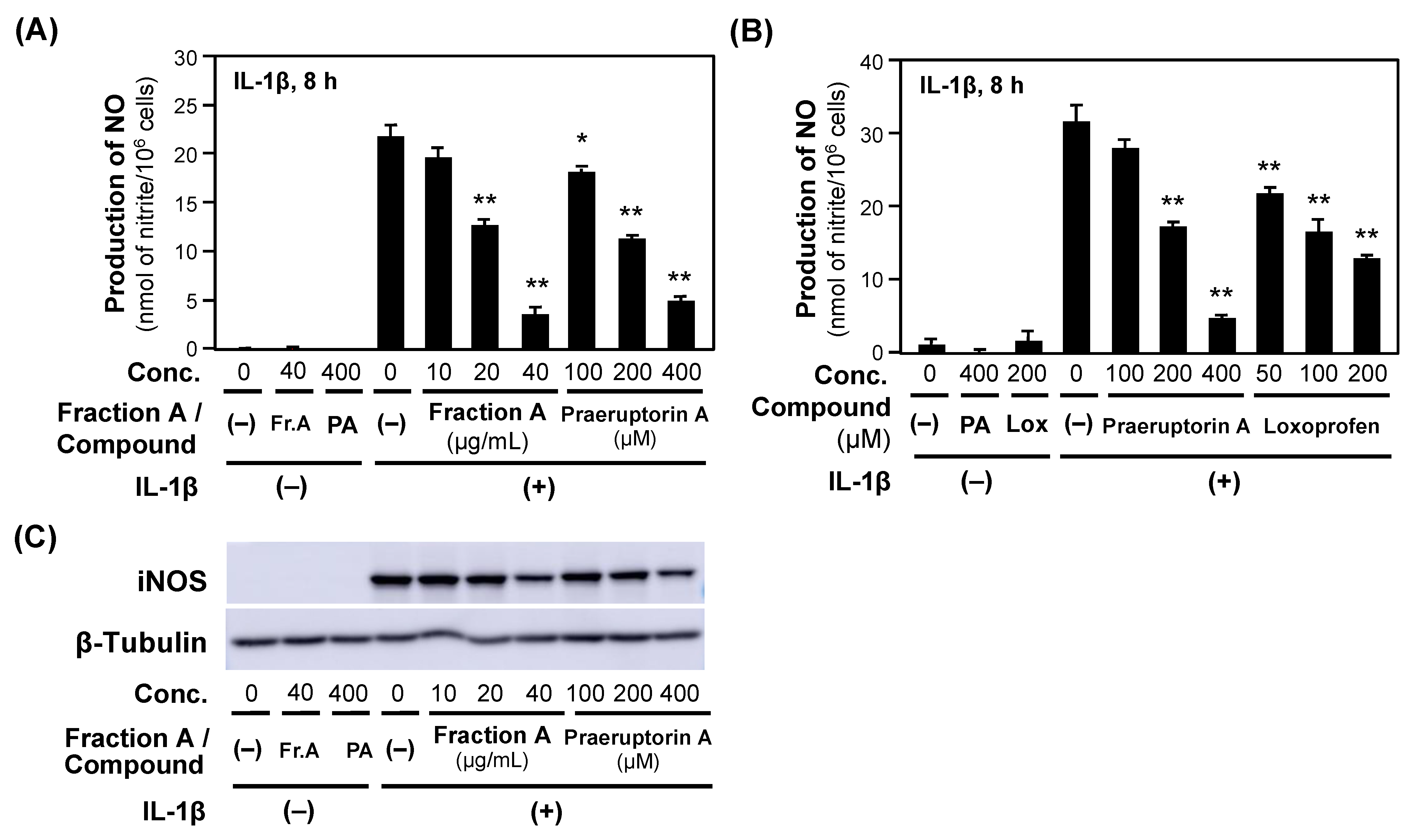
2.3. Identification of Biologically Active Compounds in Fraction A
2.4. NO Production Is Suppressed by Compounds in P. praeruptorum Fraction A
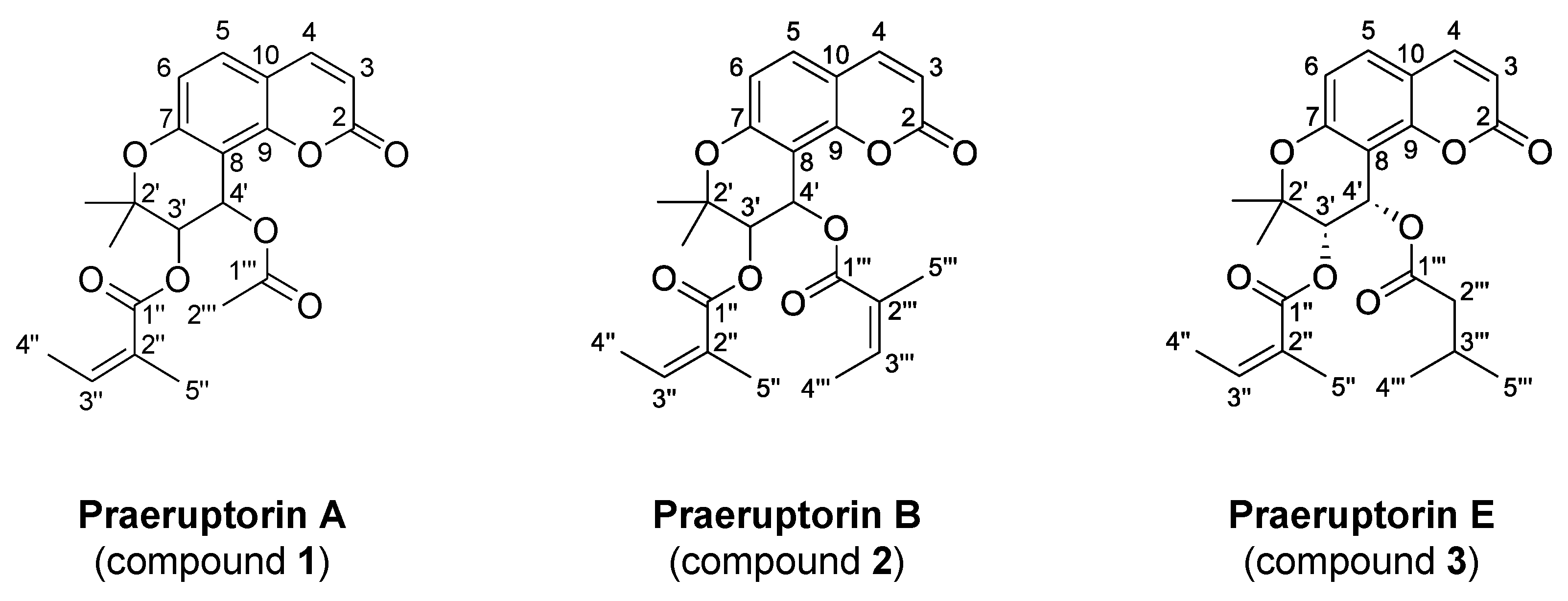
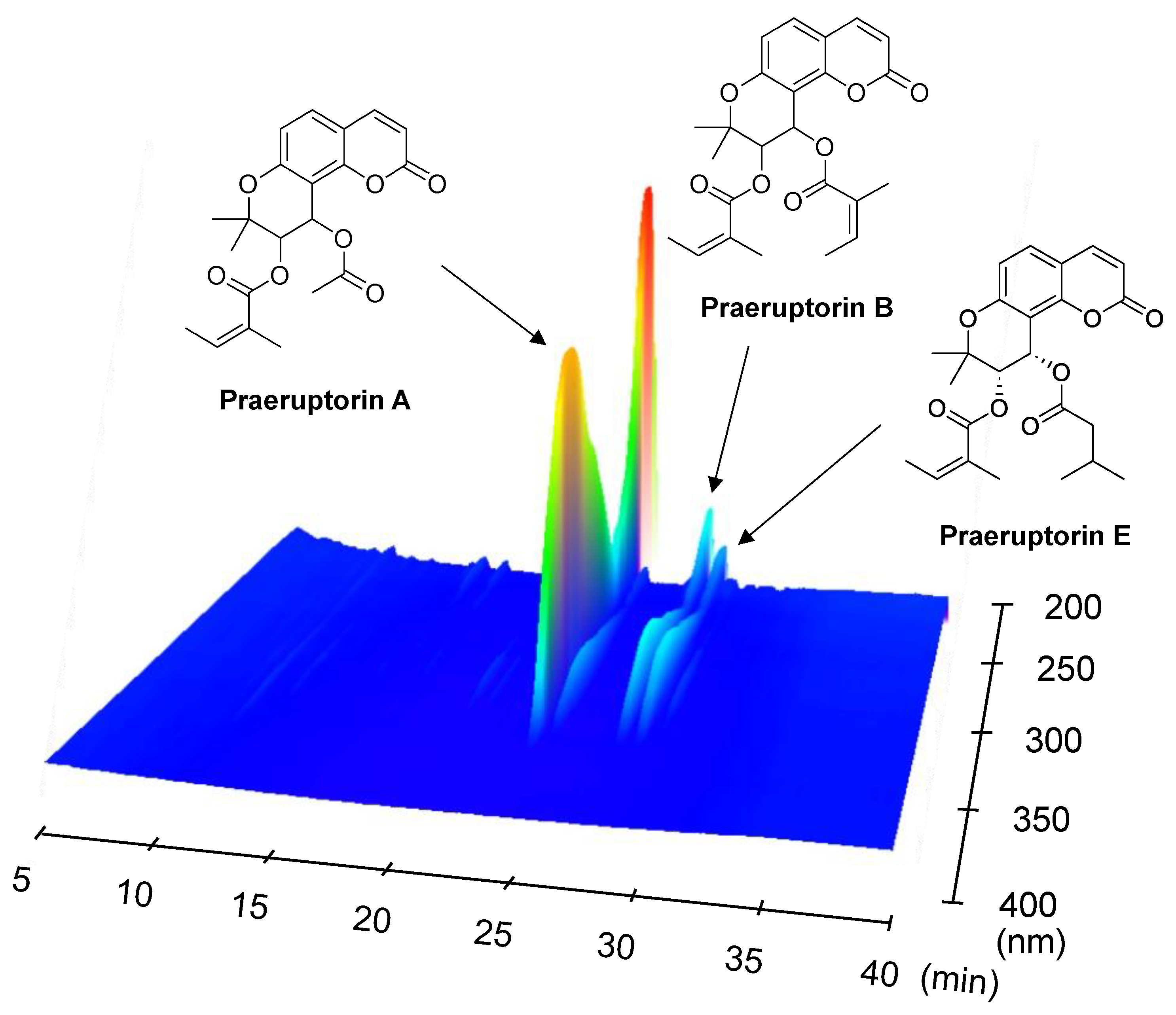
2.5. Analysis of the Compounds in Fraction A of P. praeruptorum Roots
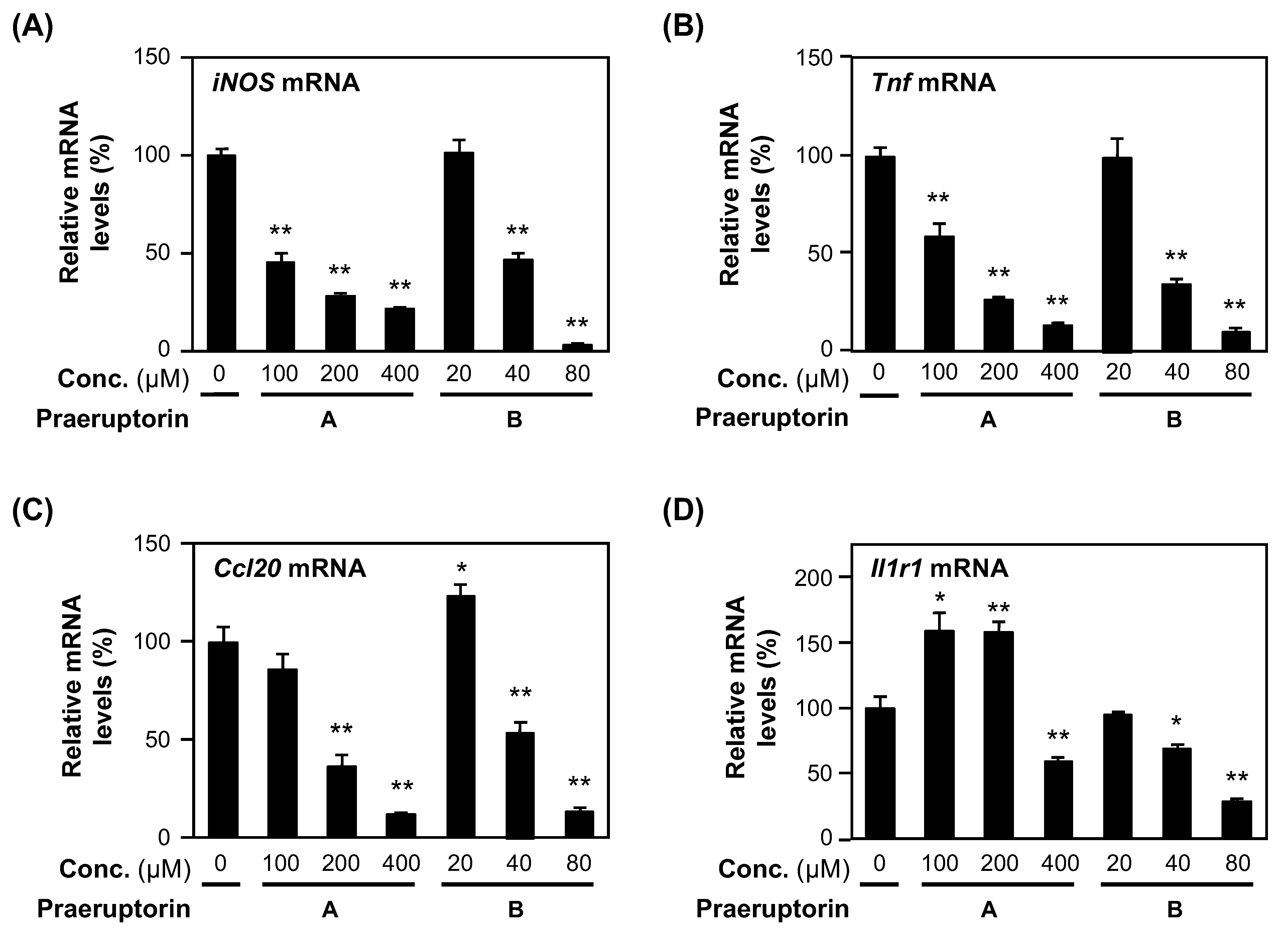
2.6. Praeruptorins A and B Decrease the mRNA Levels of Proinflammatory Genes
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Crude Fractionation of a Methanol Extract
4.4. Purification of Biologically Active Constituents from Fraction A
4.5. Measuring the Content of Compounds by HPLC
4.6. 3D-HPLC Analysis
4.7. Animals and Primary Cultured Rat Hepatocytes
4.8. Rat IL-1β
4.9. RT–qPCR
4.10. NO Assay and LDH Activity
4.11. Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Colasanti, M.; Suzuki, H. The dual personality of NO. Trends Pharmacol. Sci. 2000, 21, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Kitade, H.; Sakitani, K.; Inoue, K.; Masu, Y.; Kawada, N.; Hiramatsu, Y.; Kamiyama, Y.; Okumura, T.; Ito, S. Interleukin 1β markedly stimulates nitric oxide formation in the absence of other cytokines or lipopolysaccharide in primary cultured rat hepatocytes but not in Kupffer cells. Hepatology 1996, 23, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Inaba, H.; Yoshigai, E.; Okuyama, T.; Murakoshi, M.; Sugiyama, K.; Nishino, H.; Nishizawa, M. Antipyretic analgesic drugs have different mechanisms for regulation of the expression of inducible nitric oxide synthase in hepatocytes and macrophages. Nitric Oxide 2015, 44, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Nishizawa, M.; Ozaki, T.; Kimura, T.; Hashimoto, I.; Yamada, M.; Kaibori, M.; Kamiyama, Y.; Ito, S.; Okumura, T. Natural antisense transcript stabilizes inducible nitric oxide synthase messenger RNA in rat hepatocytes. Hepatology 2008, 47, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Okumura, T.; Ikeya, Y. Assessment of anti-inflammatory effects of Japanese Kampo medicine versus functional foods. Funct. Foods Health Dis. 2019, 9, 79–91. [Google Scholar] [CrossRef]
- Takimoto, Y.; Qian, H.Y.; Yoshigai, E.; Okumura, T.; Ikeya, Y.; Nishizawa, M. Gomisin N in the herbal drug gomishi (Schisandra chinensis) suppresses inducible nitric oxide synthase gene via C/EBPβ and NF-κB in rat hepatocytes. Nitric Oxide 2013, 28, 47–56. [Google Scholar] [CrossRef]
- Nishidono, Y.; Okada, R.; Iwama, Y.; Okuyama, T.; Nishizawa, M.; Tanaka, K. Anti-inflammatory kavalactones from Alpinia zerumbet. Fitoterapia 2020, 140, 104444. [Google Scholar] [CrossRef]
- Ulfa, S.M.; Shirako, S.; Sato, M.; Dwijayanti, D.R.; Okuyama, T.; Horie, S.; Watanabe, J.; Ikeya, Y.; Nishizawa, M. Anti-inflammatory effects of anthraquinones of Polygonum multiflorum roots. Bioact. Compd. Health Dis. 2022, 5, 136–148. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Ohno, N.; Yoshigai, E.; Okuyama, T.; Yamamoto, Y.; Okumura, T.; Sato, K.; Ikeya, Y.; Nishizawa, M. Chlorogenic acid from the Japanese herbal medicine Kinginka (Flos Lonicerae japonicae) suppresses the expression of inducible nitric oxide synthase in rat hepatocytes. HOAJ Biol. 2012, 1, 2. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Okuyama, T.; Ishii, T.; Okumura, T.; Ikeya, Y.; Nishizawa, M. Sakuranetin downregulates inducible nitric oxide synthase expression by affecting interleukin-1 receptor and CCAAT/enhancer-binding protein β. J. Nat. Med. 2019, 73, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Fujii, A.; Okuyama, T.; Wakame, K.; Okumura, T.; Ikeya, Y.; Nishizawa, M. Identification of anti-inflammatory constituents in Phellodendri Cortex and Coptidis Rhizoma by monitoring the suppression of nitric oxide production. J. Nat. Med. 2017, 71, 745–756. [Google Scholar] [CrossRef] [PubMed]
- The Committee on the Japanese Pharmacopoeia. Crude Drugs and Related Drugs. In The Japanese Pharmacopoeia, 18th ed.; The Ministry of Health, Labour and Welfare: Tokyo, Japan, 2021. Available online: https://www.mhlw.go.jp/content/11120000/000912390.pdf (accessed on 17 May 2023).
- Ali, M.Y.; Seong, S.H.; Jung, H.A.; Choi, J.S. Angiotensin-I-converting enzyme inhibitory activity of coumarins from Angelica decursiva. Molecules 2019, 24, 3937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Xu, D.; Yao, S.; Luo, J.; Kong, L. An application of high-speed counter-current chromatography coupled with electrospray ionization mass spectrometry for separation and online identification of coumarins from Peucedanum praeruptorum Dunn. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2009, 877, 2571–2578. [Google Scholar] [CrossRef]
- Song, Y.L.; Zhang, Q.W.; Li, Y.P.; Yan, R.; Wang, Y.T. Enantioseparation and absolute configuration determination of angular-type pyranocoumarins from Peucedani Radix using enzymatic hydrolysis and chiral HPLC-MS/MS analysis. Molecules 2012, 17, 4236–4251. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, Y.J.; Kim, J.; Bang, O.S. Pyranocoumarins from root extracts of Peucedanum praeruptorum Dunn with multidrug resistance reversal and anti-Inflammatory activities. Molecules 2015, 20, 20967–20978. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.J.; Ci, W.; Wang, G.F.; Zhang, J.Y.; Wu, S.Y.; Xu, W.; Jin, H.; Zhu, Z.G.; Zhang, J.J.; Pang, J.X.; et al. Praeruptorin A inhibits lipopolysaccharide-induced inflammatory response in murine macrophages through inhibition of NF-κB pathway activation. Phytother. Res. 2011, 25, 550–556. [Google Scholar] [CrossRef]
- Yu, P.J.; Jin, H.; Zhang, J.Y.; Wang, G.F.; Li, J.R.; Zhu, Z.G.; Tian, Y.X.; Wu, S.Y.; Xu, W.; Zhang, J.J.; et al. Pyranocoumarins isolated from Peucedanum praeruptorum Dunn suppress lipopolysaccharide-induced inflammatory response in murine macrophages through inhibition of NF-κB and STAT3 activation. Inflammation 2012, 35, 967–977. [Google Scholar] [CrossRef]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [Green Version]
- Yoshigai, E.; Hara, T.; Inaba, H.; Hashimoto, I.; Tanaka, Y.; Kaibori, M.; Kimura, T.; Okumura, T.; Kwon, A.H.; Nishizawa, M. Interleukin-1β induces tumor necrosis factor-α secretion from rat hepatocytes. Hepatol. Res. 2014, 44, 571–583. [Google Scholar] [CrossRef]
- Yamada, M.; Nishizawa, M.; Nakatake, R.; Habara, K.; Yoshida, H.; Ozaki, T.; Matsui, K.; Hamada, Y.; Kamiyama, Y.; Ito, S.; et al. Characterization of alternatively spliced isoforms of the type I interleukin-1 receptor on iNOS induction in rat hepatocytes. Nitric Oxide 2007, 17, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, M.A.; Oratz, M.; Schreiber, S.S. Serum albumin. Hepatology 1988, 8, 385–401. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Takusagawa, F.; Wakaki, K.; Kishi, H.; Eskandarian, M.R.; Kobayashi, M.; Date, T.; Huh, N.H.; Pitot, H.C. Rat liver serine dehydratase. Bacterial expression and two folding domains as revealed by limited proteolysis. J. Biol. Chem. 1999, 274, 12855–12860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatulian, S.A. Structural dynamics of insulin receptor and transmembrane signaling. Biochemistry 2015, 54, 5523–5532. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.X.; Sun, L.R.; Yu, W.T.; Fan, P.H.; Cui, L.; Gao, Y.H.; Ma, B.; Ren, D.M.; Ji, M. Absolute configuration determination of angular dihydrocoumarins from Peucedanum praeruptorum. J. Asian Nat. Prod. Res. 2004, 6, 177–184. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, C.; Jin, Q.; Yeon, E.T.; Lee, D.; Kim, S.Y.; Han, S.B.; Hong, J.T.; Lee, M.K.; Hwang, B.Y. Pyranocoumarins from Glehnia littoralis inhibit the LPS-induced NO production in macrophage RAW 264.7 cells. Bioorg. Med. Chem. Lett. 2014, 24, 2717–2719. [Google Scholar] [CrossRef]
- Nakano, Y.; Nasu, M.; Kano, M.; Kameoka, H.; Okuyama, T.; Nishizawa, M.; Ikeya, Y. Lignans from guaiac resin decrease nitric oxide production in interleukin 1β-treated hepatocytes. J. Nat. Med. 2017, 71, 190–197. [Google Scholar] [CrossRef]
- Kamino, T.; Shimokura, T.; Morita, Y.; Tezuka, Y.; Nishizawa, M.; Tanaka, K. Comparative analysis of the constituents in Saposhnikoviae Radix and Glehniae Radix cum Rhizoma by monitoring inhibitory activity of nitric oxide production. J. Nat. Med. 2016, 70, 253–259. [Google Scholar] [CrossRef]
- Tanemoto, R.; Okuyama, T.; Matsuo, H.; Okumura, T.; Ikeya, Y.; Nishizawa, M. The constituents of licorice (Glycyrrhiza uralensis) differentially suppress nitric oxide production in interleukin-1β-treated hepatocytes. Biochem. Biophys. Rep. 2015, 2, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Yamanishi, R.; Yoshigai, E.; Okuyama, T.; Mori, M.; Murase, H.; Machida, T.; Okumura, T.; Nishizawa, M. The anti-inflammatory effects of flavanol-rich lychee fruit extract in rat hepatocytes. PLoS ONE 2014, 9, e93818. [Google Scholar] [CrossRef]
- Nurkolis, F.; Purnomo, A.F.; Alisaputra, D.; Gunawan, W.B.; Qhabibi, F.R.; Park, W.; Moon, M.; Taslim, N.A.; Park, M.N.; Kim, B. In silico and in vitro studies reveal a synergistic potential source of novel anti-ageing from two Indonesian green algae. J. Funct. Foods 2023, 104, 105555. [Google Scholar] [CrossRef]
- Kurniawan, R.; Nurkolis, F.; Taslim, N.A.; Subali, D.; Surya, R.; Gunawan, W.B.; Alisaputra, D.; Mayulu, N.; Salindeho, N.; Kim, B. Carotenoids composition of green algae Caulerpa racemosa and their antidiabetic, anti-obesity, antioxidant, and anti-inflammatory properties. Molecules 2023, 28, 3267. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Jeong, J.H.; Park, S.H. The root extract of Peucedanum praeruptorum Dunn exerts anticancer effects in human non-small-cell lung cancer cells with different EGFR mutation statuses by suppressing MET activity. Molecules 2022, 27, 2360. [Google Scholar] [CrossRef] [PubMed]
- Nishidono, Y.; Tanaka, K. Comprehensive characterization of polyacetylenes and diterpenes from the underground parts of Solidago altissima L. and their contribution to the overall allelopathic activity. Phytochemistry 2022, 193, 112986. [Google Scholar] [CrossRef] [PubMed]
- Kanemaki, T.; Kitade, H.; Hiramatsu, Y.; Kamiyama, Y.; Okumura, T. Stimulation of glycogen degradation by prostaglandin E2 in primary cultured rat hepatocytes. Prostaglandins 1993, 45, 459–474. [Google Scholar] [CrossRef]
- Yoshigai, E.; Machida, T.; Okuyama, T.; Mori, M.; Murase, H.; Yamanishi, R.; Okumura, T.; Ikeya, Y.; Nishino, H.; Nishizawa, M. Citrus nobiletin suppresses inducible nitric oxide synthase gene expression in interleukin-1β-treated hepatocytes. Biochem. Biophys. Res. Commun. 2013, 439, 54–59. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract/Crude Fraction | Yield [%] 1 | IC50 [μg/mL] 2 |
|---|---|---|
| Methanol extract | 100.0 | 154.3 ± 26.8 |
| Fraction A (EtOAc-soluble) | 29.3 | 24.1 ± 3.0 |
| Fraction B (n-butanol-soluble) | 8.76 | 74.4 ± 27.3 |
| Fraction C (water-soluble) | 62.0 | NA |
| Constituent (Compound) | Content [%] 1 | IC50 [μM] 2 |
|---|---|---|
| Fraction A | 100.0 | –– |
| Praeruptorin A (1) | 24.0 | 208 ± 77.3 |
| Praeruptorin B (2) | 7.75 | 43.5 ± 7.54 |
| Praeruptorin E (3) | 9.07 | 57.4 ± 7.00 |
| Name 1 | Gene 2 | Primers (5’-to-3’ Direction) |
|---|---|---|
| Albumin | Alb | TGACAAGTCCATTCACACTCTCT |
| AACTTCATGCAAATAGTGTCCCAG | ||
| CCL20 | Ccl20 | CAGCCAGTCAGAAGCAGCAAGC |
| CTTGGTTCTTAGGCTGAGGAGGT | ||
| EF-1α | Eef1a1 | TCTGGTTGGAATGGTGACAACATGC |
| CCAGGAAGAGCTTCACTCAAAGCTT | ||
| GR | Nr3c1 | CAAAGGCGATACCAGGCTTCAGAA |
| GAGACATTCTCTGCTCATTAATAATC | ||
| IL1R1 | Il1r1 | GTTAGTATACCCAGTTCCTGACT |
| GTCTTTCCATCTGAAGCTTTTGG | ||
| iNOS | Nos2 | CCAACCTGCAGGTCTTCGATG |
| GTCGATGCACAACTGGGTGAAC | ||
| Insr | Insr | CTTCCGACATGTGGTCCTTTGG |
| TCAGGTCAGTGAGTCTCTCTGGA | ||
| TNF-α | Tnf | TCCCAACAAGGAGGAGAAGTTCC |
| GGCAGCCTTGTCCCTTGAAGAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozaki, H.; Nishidono, Y.; Fujii, A.; Okuyama, T.; Nakamura, K.; Maesako, T.; Shirako, S.; Nakatake, R.; Tanaka, K.; Ikeya, Y.; et al. Identification of Anti-Inflammatory Compounds from Peucedanum praeruptorum Roots by Using Nitric Oxide-Producing Rat Hepatocytes Stimulated by Interleukin 1β. Molecules 2023, 28, 5076. https://doi.org/10.3390/molecules28135076
Ozaki H, Nishidono Y, Fujii A, Okuyama T, Nakamura K, Maesako T, Shirako S, Nakatake R, Tanaka K, Ikeya Y, et al. Identification of Anti-Inflammatory Compounds from Peucedanum praeruptorum Roots by Using Nitric Oxide-Producing Rat Hepatocytes Stimulated by Interleukin 1β. Molecules. 2023; 28(13):5076. https://doi.org/10.3390/molecules28135076
Chicago/Turabian StyleOzaki, Hiromu, Yuto Nishidono, Airi Fujii, Tetsuya Okuyama, Kaito Nakamura, Takanori Maesako, Saki Shirako, Richi Nakatake, Ken Tanaka, Yukinobu Ikeya, and et al. 2023. "Identification of Anti-Inflammatory Compounds from Peucedanum praeruptorum Roots by Using Nitric Oxide-Producing Rat Hepatocytes Stimulated by Interleukin 1β" Molecules 28, no. 13: 5076. https://doi.org/10.3390/molecules28135076