

Fractionation Coupled to Molecular Networking: Towards Identification of Anthelmintic Molecules in Terminalia leiocarpa (DC.) Baill

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cytotoxicity and Anthelminthic Activities of Fractions
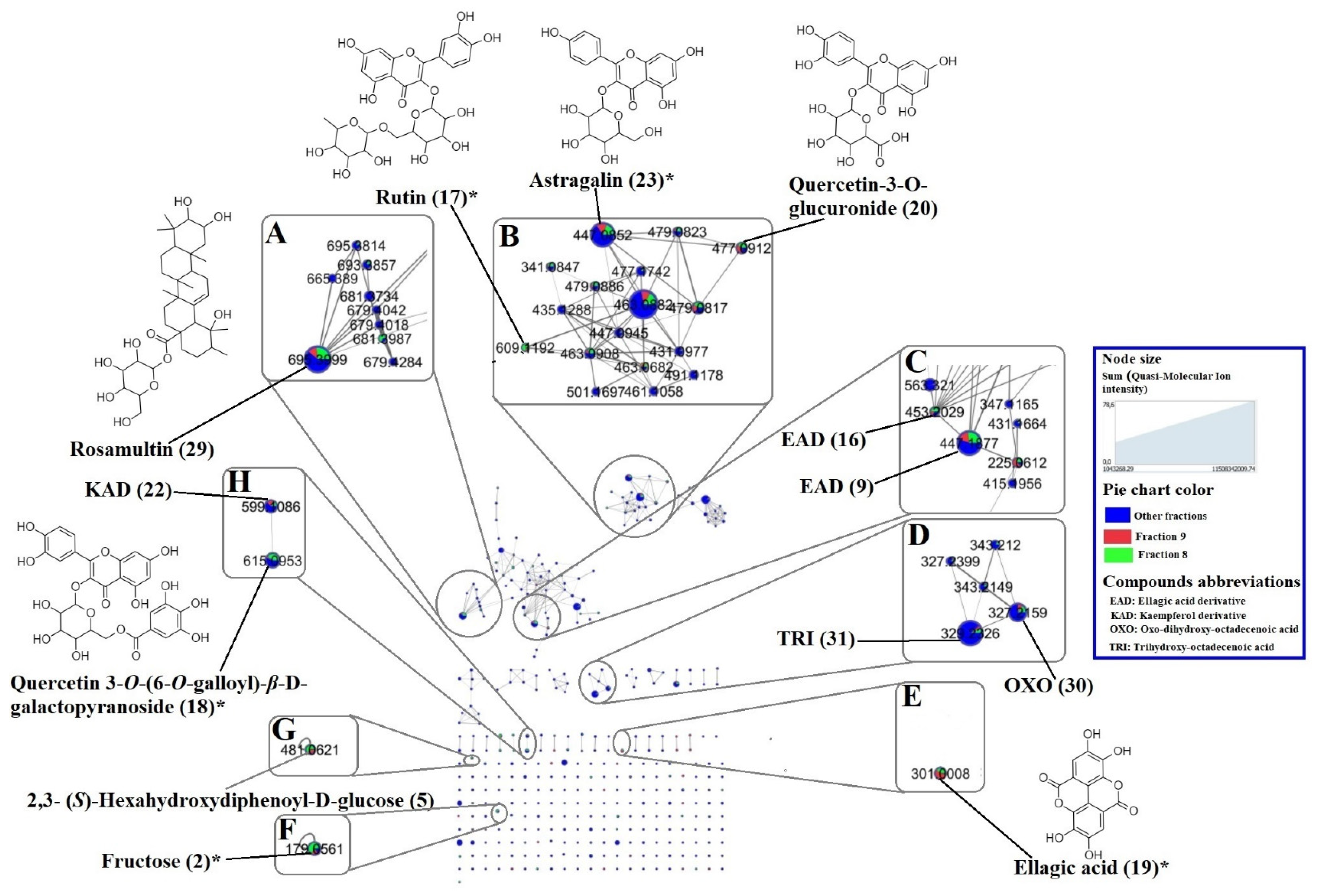
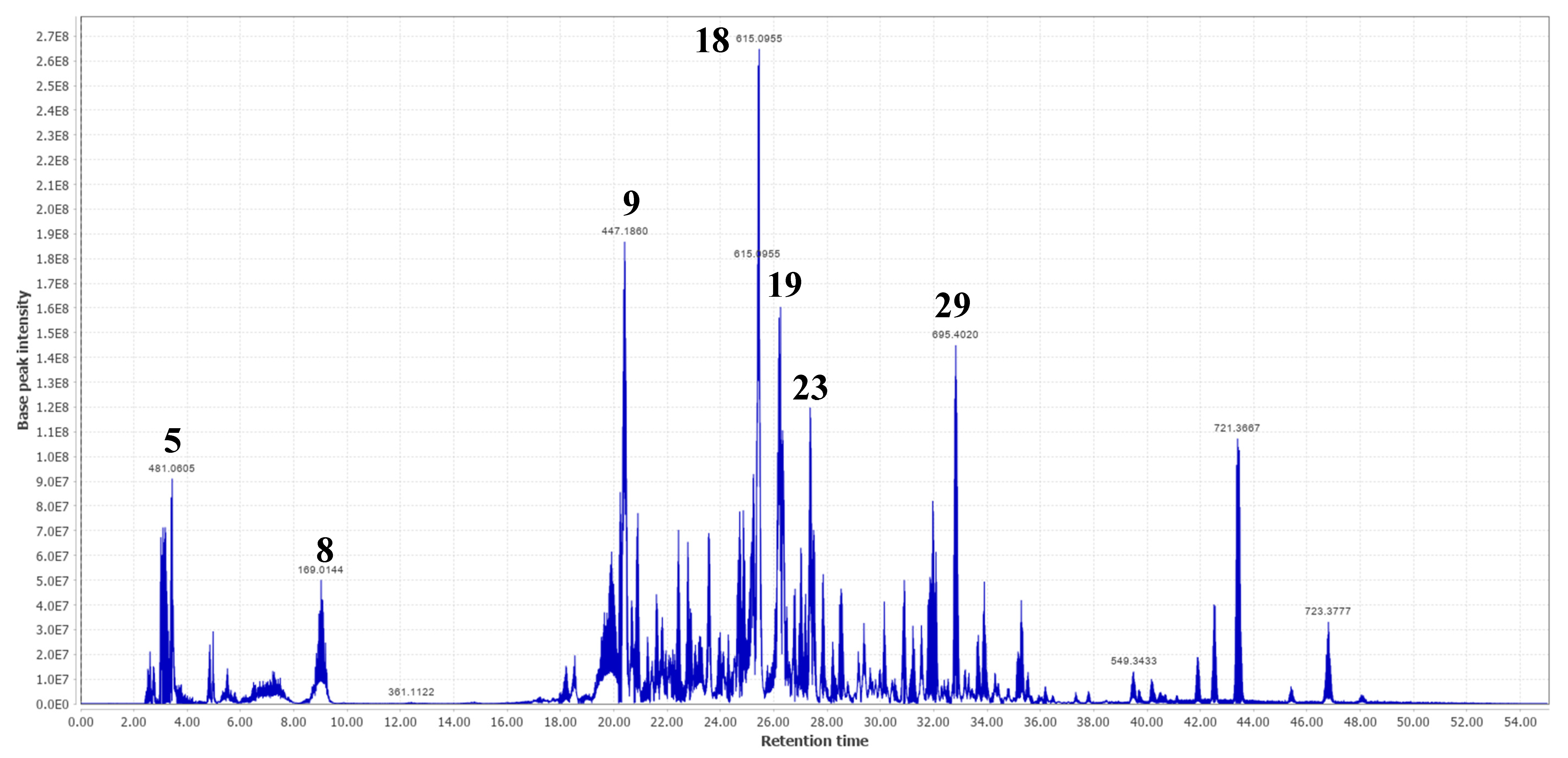
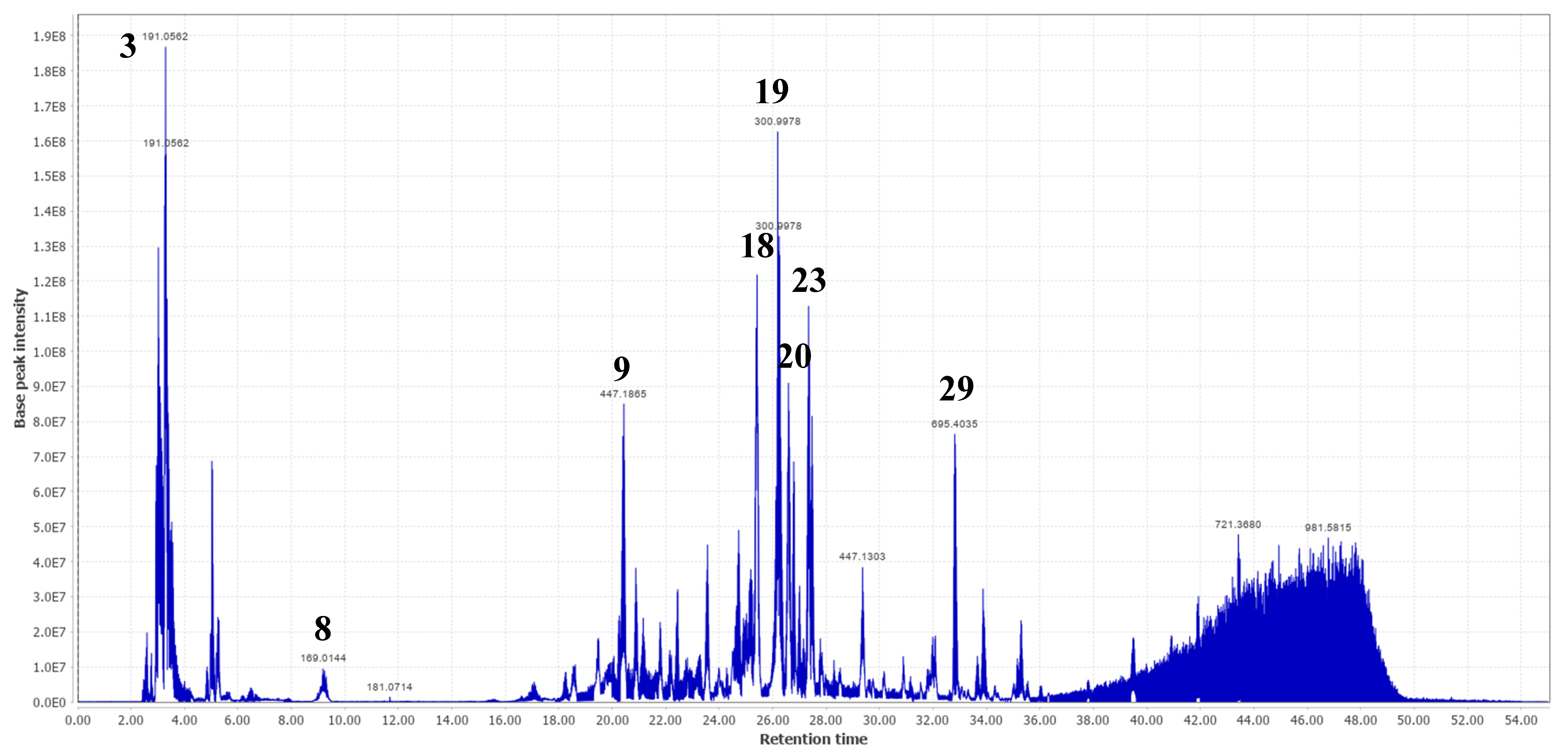
2.2. Molecular Networking and Major Compounds in the Most Active Fractions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | RT (min) | Molecular Formula | Quasi- Molecular Ion | MS/MS Fragment | Molecular Mass | Error (ppm) | Identification | Isolated Previously from | Source | References | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Observed | Theoretical | F8# | F9# | |||||||||
| 1 | 3.02 | C6H14O6 | 181.0715 [M-H]- | 179.0560 144.0665 101.0245 163.0610 | 182.0794 | 182.0790 | 1.99 | Hexitol | √ | |||
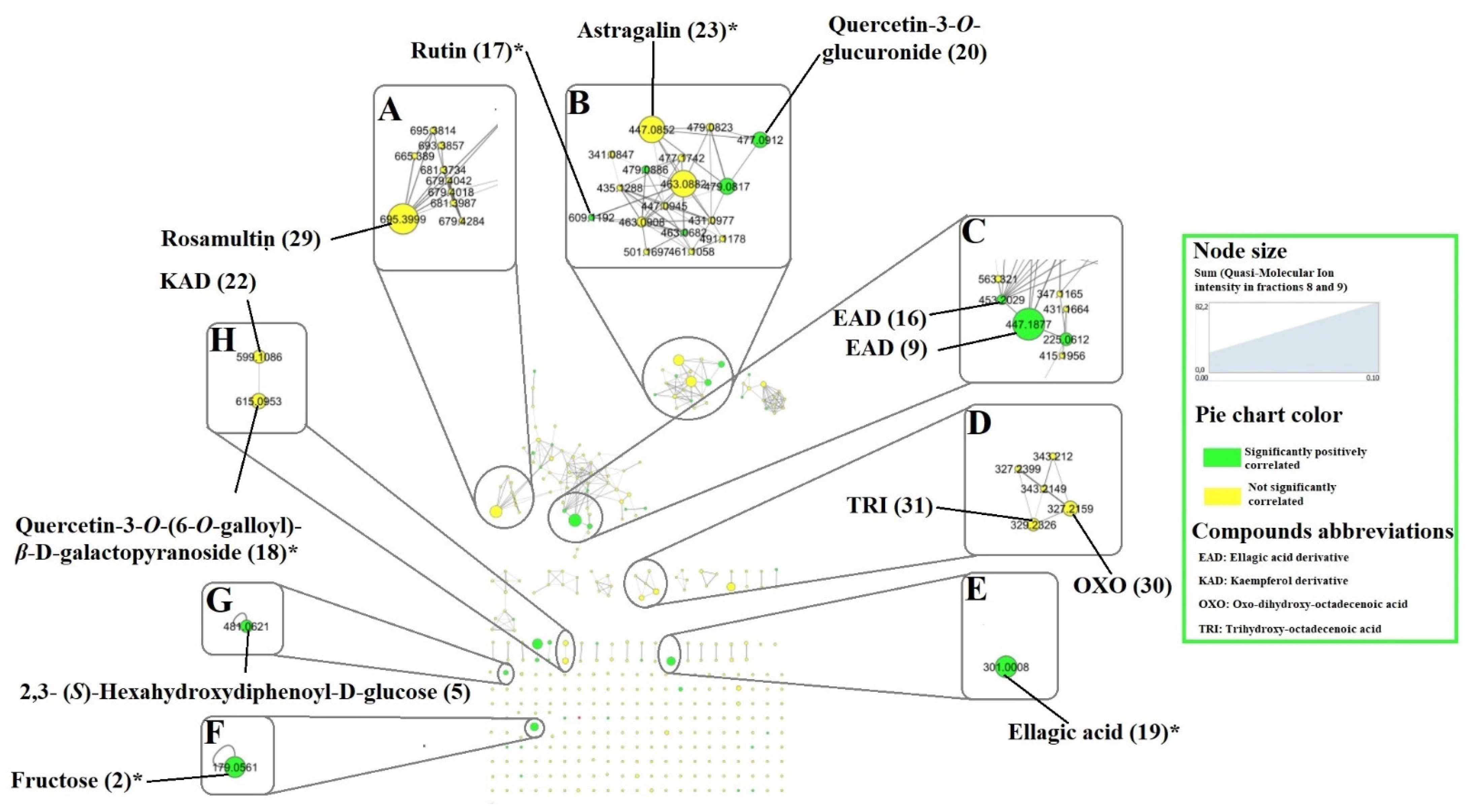
| 2 | 3.17 | C6H12O6 | 179.0564 [M-H]- | 161.0460 113.0248 | 180.0643 | 180.0634 | 5.06 | Fructose * | √ | √ | ||
| 3 | 3.29 | C7H12O6 | 191.0562 [M-H]- | 181.0715 179.0569 173.0465 189.8369 | 192.0641 | 192.0634 | 3.71 | Quinic acid * | Terminalia ferdinandiana | √ | √ | [35] |
| 4 | 3.40 | C7H10O5 | 173.0456 [M-H]- | 119.0353 129.0198 137.0243 155.0348 | 174.0535 | 174.0528 | 3.89 | Shikimic acid | Anogeissus latifolia | √ | √ | [3] |
| 5 | 3.44 | C20H18O14 | 481.0605 [M-H]- | 331.0672 421.1343 173.0456 300.9998 375.1294 | 482.0684 | 482.0697 | −2.70 | 2,3- (S)-Hexahydroxydiphenoyl-D-glucose | T. calamansanai, T. myriocarpa | √ | [29,30] | |
| 6 | 4.97 | C9H18O7 | 283.1037 [M+HCOO]- | 243.0630 273.0739 179.0564 | 238.1061 | 238.1053 | 3.56 | n.i | √ | |||
| 7 | 5.51 | C17H26O12 | 421.1360 [M-H]- | 375.1310 287.0888 267.0739 357.1195 331.0686 | 422.1439 | 422.1424 | 3.49 | n.i | √ | |||
| 8 | 9.02 | C₇H₆O₅ | 169.0144 [M-H]- | 125.0247 168.0070 124.0173 126.0283 | 170.0223 | 170.0215 | 4.57 | Gallic acid * | A. leiocarpa, T. ferdinandiana | √ | √ | [2,25,35] |
| 9 | 20.43 | 447.1860 [M-H]- | 401.1822 300.9978 179.0560 | Ellagic acid derivative | √ | √ | ||||||
| 10 | 20.90 | 477.1626 [M-H]- | 431.1540 445.1712 300.9982 169.0147 | Ellagic acid derivative | √ | |||||||
| 11 | 21.60 | 447.1515 [M-H]- | 300.9987 289.0723 387.1662 169.0150 | Ellagic acid derivative | √ | |||||||
| 12 | 22.43 | 431.1910 [M-H]- | 387.1653 169.0145 300.9980 327.1093 | Ellagic acid derivative | √ | √ | ||||||
| 13 | 22.78 | 387.1660 [M-H]- | 169.0149 301.0005 | Ellagic acid derivative | √ | |||||||
| 14 | 23.56 | 453.1048 [M-H]- | 387.1666 289.0226 439.0686 169.0143 300.9990 125.0252 | Ellagic acid derivative | √ | √ | ||||||
| 15 | 24.73 | 431.1912 [M-H]- | 300.9982 169.0146 289.0718 125.0248 205.1234 | Ellagic acid derivative | √ | √ | ||||||
| 16 | 24.86 | 453.1979 [M-H]- | 433.2072 300.9979 407.1930 169.0145 | Ellagic acid derivative | √ | |||||||
| 17 | 25.24 | C27H30O16 | 609.1453 [M-H]- | 459.1496 301.0353 313.0574 567.2086 169.0144 | 610.1532 | 610.1534 | −0.30 | Rutin * | A. leiocarpa | √ | [21] | |
| 18 | 25.44 | C28H24O16 | 615.0955 [M-H]- | 301.0353 313.0553 565.2844 463.0887 169.0144 | 616.1034 | 616.1064 | −4.93 | Quercetin-3-O-(6-O-galloyl)-β-D-galactopyranoside | T. guianensis | √ | √ | [26] |
| 19 | 26.22 | C14H6O8 | 300.9978 [M-H]- | 163.0398 169.0144 | 302.0057 | 302.0063 | −1.88 | Ellagic acid * | A. leiocarpa Terminalia brownii | √ | √ | [4,7] |
| 20 | 26.59 | C21H18O13 | 477.0677 [M-H]- | 301.0354 302.0383 169.0145 439.0670 151.0035 289.0715 287.0564 | 478.0756 | 478.0747 | 1.80 | Quercetin-3-O-glucuronide | √ | |||
| 21 | 26.79 | C48H68O5 | 723.5013 [M-H]- | 439.0679 169.0140 463.0896 303.0508 289.0721 677.5002 125.0249 | 724.5092 | 724.5067 | 3.48 | n.i | √ | |||
| 22 | 27.02 | C28H24O15 | 599.1047 [M-H]- | 435.1282 285.0406 473.1672 313.0556 | 600.1126 | 600.1115 | 1.80 | Kaempferol linked to gallate and deoxy-hexose | √ | √ | ||
| 23 | 27.35 | C21H20O11 | 447.0932 [M-H]- | 285.0404 284.0323 439.0670 442.7359 289.0715 | 448.1011 | 448.1006 | 1.20 | Astragalin * | P. suberosa | √ | √ | [27] |
| 24 | 27.85 | C37H60O14 | 727.3909 [M-H]- | 565.3358 519.3334 439.0675 477.1035 343.2121 | 728.3988 | 728.3983 | 0.68 | n.i | √ | |||
| 25 | 28.53 | C37H60O13 | 711.3926 [M-H]- | 343.2126 371.1710 531.1526 439.0681 583.1072 289.0722 | 712.4005 | 712.4034 | −4.06 | n.i | √ | |||
| 26 | 29.36 | C22H24O10 | 447.1303 [M-H]- | 439.0685 303.0513 169.0151 287.0574 289.0725 125.0251 | 448.1382 | 448.1369 | 2.80 | n.i | √ | |||
| 27 | 30.90 | C30H62O18 | 709.3831 [M-H]- | 507.2063 547.3296 501.3242 597.1829 461.1088 | 710.3910 | 710.3936 | −3.68 | n.i | √ | √ | ||
| 28 | 31.96 | C37H60O13 | 711.3934 [M-H]- | 549.3431 697.3820 503.3406 695.4033 702.6718 | 712.4013 | 712.4034 | −2.94 | n.i | √ | |||
| 29 | 32.82 | C36H58O10 | 695.4020 [M + HCOO]- | 487.3446 173.9751 686.9651 533.3465 | 650.4043 | 650.4030 | 2.00 | Rosamultin or isomer | T. alata | √ | √ | [31] |
| 30 | 33.88 | C18H32O5 | 327.2178 [M-H]- | 324.4868 211.1343 289.0727 171.1030 229.1447 | 328.2257 | 328.2250 | 2.21 | Oxo-dihydroxy-octadecenoic acid | Globularia spp. Bituminaria bituminosa Sasa veitchii | √ | √ | [32,33,34] |
| 31 | 35.28 | C18H33O5 | 329.2329 [M-H]- | 211.1343 116.0257 229.1447 326.4767 169.0144 | 330.2408 | 330.2406 | 0.53 | Trihydroxy-octadecenoic acid | Globularia spp. B. bituminosa S. veitchii | √ | √ | [32,33,34] |
| 32 | 43.40 | C34H58O16 | 721.3667 [M-H]- | 675.3602 397.1340 712.4544 277.2173 | 722.3746 | 722.3725 | 2.93 | n.i | √ | √ | ||
| 33 | 46.81 | C34H60O16 | 723.3777 [M-H]- | 677.3750 116.0257 397.1347 167.0363 119.0260 | 724.3856 | 724.3881 | −3.50 | n.i | √ | |||
2.3. Prediction of Anthelmintic Activity of Features Detected in T. leiocarpa Fractions by Pearson Correlation
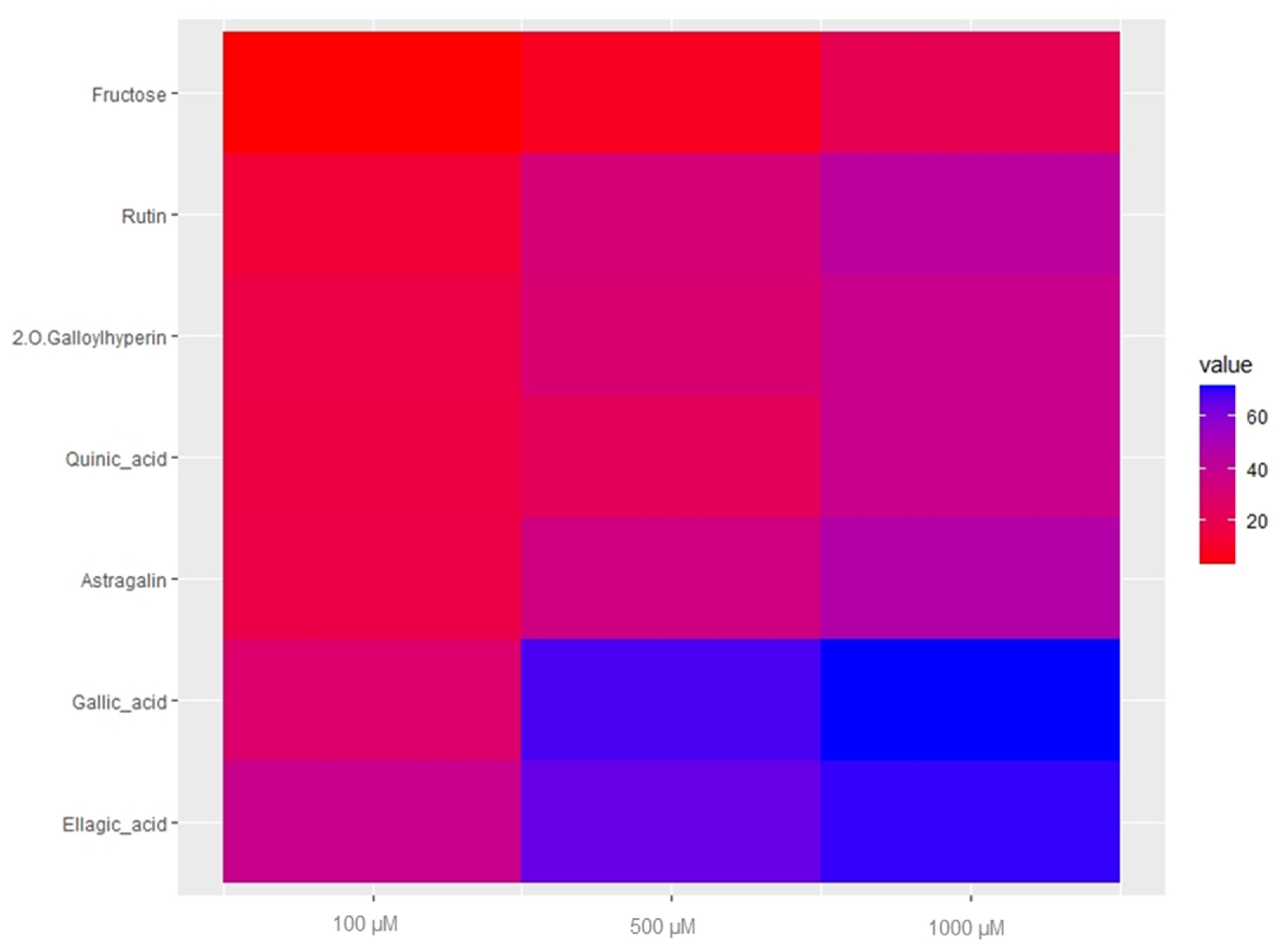
2.4. Anthelmintic Activity of Major Compounds and Their Quantification
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Collection
3.3. Extraction Procedure
3.4. Open Column Chromatography (OCC) Fractionation of T. leiocarpa MeOH Extract
3.5. Fractions Cytotoxicity
3.6. Anthelmintic Activities of Fractions
3.6.1. Viability of C. elegans Adult Worms Treated with Fractions and Pure Compounds
3.6.2. Larval Migration Inhibition Assay (LAMIA)
3.7. HPLC-PDA-HRMS/MS Analysis
3.8. Data Processing on MZmine
3.9. Dereplication on GNPS
3.10. Quantification of Major Compounds
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ademola, I.; Eloff, J. In Vitro Anthelmintic Effect of Anogeissus leiocarpus (Dc.) Guill. & Perr. Leaf Extracts and Fractions on Developmental Stages of Haemonchus contortus. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 134–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterman, C.; Patel, Z.; Kim, S.; Rivera, A.; Pontiggia, L.; Grace, M.H.; Smith, R. Anthelmintic Activity of Punicalagin from Anogeissus leiocarpus. Univers. J. Plant Sci. 2015, 3, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Baghel, U.S.; Gautam, A.; Baghel, D.S.; Yadav, D.; Malik, J.; Yadav, R. The Genus Anogeissus: A Review on Ethnopharmacology, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2016, 194, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Salih, E.Y.A.; Julkunen-Tiitto, R.; Luukkanen, O.; Sipi, M.; Fahmi, M.K.M.; Fyhrquist, P.J. Potential Anti-Tuberculosis Activity of the Extracts and Their Active Components of Anogeissus leiocarpa (DC.) Guill. and Perr. with Special Emphasis on Polyphenols. Antibiotics 2020, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Tchetan, E.; Olounlade, A.P.; Houehanou, T.D.; Azando, E.V.B.; Kaneho, J.A.; Houinato, M.R.B.; Hounzangbe-Adote, S.M.; Quetin-Leclercq, J.; Gbaguidi, F.A. Ethnoveterinary Knowledge of Sheep and Goat Farmers in Benin (West Africa): Effect of Socioeconomic and Environmental Factors. Heliyon 2021, 7, e07656. [Google Scholar] [CrossRef]
- Ndjonka, D.; Ajonina-Ekoti, I.; Djafsia, B.; Lüersen, K.; Abladam, E.; Liebau, E. Anogeissus leiocarpus Extract on the Parasite Nematode Onchocerca ochengi and on Drug Resistant Mutant Strains of the Free-Living Nematode Caenorhabditis elegans. Vet. Parasitol. 2012, 190, 136–142. [Google Scholar] [CrossRef]
- Salih, E.Y.A.; Kanninen, M.; Sipi, M.; Luukkanen, O.; Hiltunen, R.; Vuorela, H.; Julkunen-Tiitto, R.; Fyhrquist, P. Tannins, Flavonoids and Stilbenes in Extracts of African Savanna Woodland Trees Terminalia brownii, Terminalia laxiflora and Anogeissus leiocarpus Showing Promising Antibacterial Potential. S. Afr. J. Bot. 2017, 108, 370–386. [Google Scholar] [CrossRef]
- Kabore, A.; Belem, G.A.M.; Sawadogo, L. In Vitro Anthelmintic Effect of Two Medicinal Plants (Anogeissus leiocarpus and Daniellia oliveri) on Haemonchus contortus, an Abosomal Nematode of Sheep in Burkina Faso. Afr. J. Biotechnol. 2009, 8, 4690–4695. [Google Scholar]
- Soro, D.; Koné, W.M.; Bonfoh, B.; Dro, B.; Toily, K.B.; Kamanzi, K. In Vivo Anthelmintic Activity of Anogeissus leiocarpus Guill & Perr (Combretaceae) against Nematodes in Naturally Infected Sheep. Parasitol. Res. 2013, 112, 2681–2688. [Google Scholar] [CrossRef]
- Tchetan, E.; Olounladé, P.A.; Azando, E.V.B.; Khaliq, H.A.; Ortiz, S.; Houngbeme, A.; Alowanou, G.G.; Koura, B.I.; Akouedegni, G.C.; Houinato, M.R.B.; et al. Anthelmintic Activity, Cytotoxicity, and Phytochemical Screening of Plants Used to Treat Digestive Parasitosis of Small Ruminants in Benin (West Africa). Animals 2022, 12, 2718. [Google Scholar] [CrossRef]
- Ndjonka, D.; Abladam, E.D.; Djafsia, B.; Ajonina-Ekoti, I.; Achukwi, M.D.; Liebau, E. Anthelmintic Activity of Phenolic Acids from the Axlewood Tree Anogeissus leiocarpus on the FIlarial Nematode Onchocerca ochengi and Drug-Resistant Strains of the Free-Living Nematode Caenorhabditis elegans. J. Helminthol. 2014, 88, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Varady, M.; Papadopoulos, E.; Dolinská, M.; Königová, A. Anthelmintic Resistance in Parasites of Small Ruminants: Sheep versus Goats. Helminthologia 2011, 48, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Abongwa, M.; Martin, R.J.; Robertson, A.P. A Brief Review on the Mode of Action of Antinematodal Drugs. Acta Vet. 2017, 67, 137–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef] [PubMed]
- Quinn, R.A.; Nothias, L.-F.; Vining, O.; Meehan, M.; Esquenazi, E.; Dorrestein, P.C. Molecular Networking As a Drug Discovery, Drug Metabolism, and Precision Medicine Strategy. Trends Pharmacol. Sci. 2017, 38, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible Molecular Networking of Untargeted Mass Spectrometry Data Using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.; Ho, R.; Greff, S.; Herbette, G.; Filaire, E.; Ranouille, E.; Berthon, J.-Y.; Raharivelomanana, P. Feature-Based Molecular Networks Identification of Bioactive Metabolites from Three Plants of the Polynesian Cosmetopoeia Targeting the Dermal Papilla Cells of the Hair Cycle. Molecules 2021, 27, 105. [Google Scholar] [CrossRef]
- Abdul Khaliq, H.; Ortiz, S.; Alhouayek, M.; Muccioli, G.G.; Quetin-Leclercq, J. Dereplication and Quantification of Major Compounds of Convolvulus arvensis L. Extracts and Assessment of Their Effect on LPS-Activated J774 Macrophages. Molecules 2022, 27, 963. [Google Scholar] [CrossRef]
- Hahnel, S.R.; Dilks, C.M.; Heisler, I.; Andersen, E.C.; Kulke, D. Caenorhabditis elegans in Anthelmintic Research–Old Model, New Perspectives. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 237–248. [Google Scholar] [CrossRef]
- Coles, G.C.; Jackson, F.; Pomroy, W.E.; Prichard, R.K.; von Samson-Himmelstjerna, G.; Silvestre, A.; Taylor, M.A.; Vercruysse, J. The Detection of Anthelmintic Resistance in Nematodes of Veterinary Importance. Vet. Parasitol. 2006, 136, 167–185. [Google Scholar] [CrossRef]
- Attioua, B.; Lagnika, L.; Yeo, D.; Antheaume, C.; Kaiser, M.; Weniger, B.; Lobstein, A.; Vonthron-Sénécheau, C. In Vitro Antiplasmodial and Antileishmanial Activities of Flavonoids from Anogeissus leiocarpus (Combretaceae). Int. J. Pharm. Sci. Rev. 2011, 11, 7. [Google Scholar]
- Razgonova, M.; Zakharenko, A.; Pikula, K.; Manakov, Y.; Ercisli, S.; Derbush, I.; Kislin, E.; Seryodkin, I.; Sabitov, A.; Kalenik, T.; et al. LC-MS/MS Screening of Phenolic Compounds in Wild and Cultivated Grapes Vitis Amurensis Rupr. Molecules 2021, 26, 3650. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; Destandau, E.; Fougère, L.; Lafosse, M. Isolation by Pressurised Fluid Extraction (PFE) and Identification Using CPC and HPLC/ESI/MS of Phenolic Compounds from Brazilian Cherry Seeds (Eugenia uniflora L.). Food Chem. 2014, 145, 522–529. [Google Scholar] [CrossRef]
- Wu, N.; Li, L.; Cai, Z.-C.; Yuan, J.-H.; Wang, W.-X.; Yin, S.-X.; Liu, S.-J.; Wei, L.-F.; Mei, Y.-Q.; Chen, C.-H.; et al. Quality Evaluation of Taxilli Herba from Different Hosts Based on Simultaneous Determination of Multiple Bioactive Constituents Combined with Multivariate Statistical Analysis. Molecules 2021, 26, 7490. [Google Scholar] [CrossRef]
- Das Neves Costa, F.; Jerz, G.; Hewitson, P.; de Souza Figueiredo, F.; Ignatova, S. Laguncularia Racemosa Phenolics Profiling by Three-Phase Solvent System Step-Gradient Using High-Performance Countercurrent Chromatography with Off-Line Electrospray Mass-Spectrometry Detection. Molecules 2021, 26, 2284. [Google Scholar] [CrossRef] [PubMed]
- Calassara, L.L.; Pinto, S.C.; Condack, C.P.M.; Leite, B.F.; Nery, L.C.d.E.S.; Tinoco, L.W.; Aguiar, F.A.; Leal, I.C.R.; Martins, S.M.; da Silva, L.L.; et al. Isolation and Characterization of Flavonoids from Tapirira guianensis Leaves with Vasodilatory and Myeloperoxidase-Inhibitory Activities. Nat. Prod. Res. 2021, 35, 5480–5483. [Google Scholar] [CrossRef]
- De Leo, M.; Braca, A.; Sanogo, R.; Cardile, V.; DeTommasi, N.; Russo, A. Antiproliferative Activity of Pteleopsis suberosa Leaf Extract and Its Flavonoid Components in Human Prostate Carcinoma Cells. Planta Med. 2006, 72, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.A.; Antunes, A.M.M.; Caldeira, I.; Anjos, O.; de Freitas, V.; Fargeton, L.; Boissier, B.; Catarino, S.; Canas, S. Identification of Gallotannins and Ellagitannins in Aged Wine Spirits: A New Perspective Using Alternative Ageing Technology and High-Resolution Mass Spectrometry. Food Chem. 2022, 382, 132322. [Google Scholar] [CrossRef]
- Tanaka, T.; Morita, A.; Nonaka, G.; Lin, T.; Nishioka, I.; Ho, F. Tannins and Related Compounds. CIII. Isolation and Characterizatino of New Monomeric, Dimeric and Trimeric Ellagitannins, Calamansanin and Calamanins A, B and C, from Terminalia calamansanai (BLANCO) ROLFE. Chem. Pharm. Bull. 1991, 39, 60–63. [Google Scholar] [CrossRef] [Green Version]
- Marzouk, M.S.A.; El-Toumy, S.A.A.; Moharram, F.A. Pharmacologically Active Ellagitannins from Terminalia Myriocarpa. Planta Med. 2002, 68, 523–527. [Google Scholar] [CrossRef]
- Srivastava, S.K.; Chouksey, B.K.; Srivastava, S.D. Triterpenoid Glycoside from the Roots of Terminalia alata. Fitoterapia 2001, 72, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Friščić, M.; Bucar, F.; Hazler Pilepić, K. LC-PDA-ESI-MSn Analysis of Phenolic and Iridoid Compounds from Globularia Spp.: Phytochemical Analysis of Globularia Spp. J. Mass Spectrom. 2016, 51, 1211–1236. [Google Scholar] [CrossRef] [PubMed]
- Llorent-Martínez, E.J.; Spínola, V.; Gouveia, S.; Castilho, P.C. HPLC-ESI-MSn Characterization of Phenolic Compounds, Terpenoid Saponins, and Other Minor Compounds in Bituminaria bituminosa. Ind. Crops Prod. 2015, 69, 80–90. [Google Scholar] [CrossRef]
- Van Hoyweghen, L.; De Bosscher, K.; Haegeman, G.; Deforce, D.; Heyerick, A. In Vitro Inhibition of the Transcription Factor NF-ΚB and Cyclooxygenase by Bamboo Extracts: Bamboo Extracts Inhibit Nf-Κb And Cyclooxygenase. Phytother. Res. 2014, 28, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Sirdaarta, J.; Matthews, B.; White, A.; Cock, I.E. GC-MS and LC-MS Analysis of Kakadu Plum Fruit Extracts Displaying Inhibitory Activity against Microbial Triggers of Multiple Sclerosis. Pharmacogn. Commun. 2015, 5, 100–115. [Google Scholar] [CrossRef] [Green Version]
- Mondal, H.; Hossain, H.; Awang, K.; Saha, S.; Mamun-Ur-Rashid, S.; Islam, K.; Rahman, S.; Jahan, I.A.; Rahman, M.M.; Shilpi, J.A. Anthelmintic Activity of Ellagic Acid, a Major Constituent of Alternanthera sessilis Against Haemonchus contortus. Pak. Vet. J. 2015, 35, 58–62. [Google Scholar]
- Hussein, D.; El-Shiekh, R.A.; Saber, F.R.; Attia, M.M.; Mousa, M.R.; Atta, A.H.; Abdel-Sattar, E.; Mouneir, S.M. Unravelling the Anthelmintic Bioactives from Jasminum grandiflorum L. Subsp. Floribundum Adopting in Vitro Biological Assessment. J. Ethnopharmacol. 2021, 275, 114083. [Google Scholar] [CrossRef]
- Klongsiriwet, C.; Quijada, J.; Williams, A.R.; Mueller-Harvey, I.; Williamson, E.M.; Hoste, H. Synergistic Inhibition of Haemonchus contortus Exsheathment by Flavonoid Monomers and Condensed Tannins. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 127–134. [Google Scholar] [CrossRef]
- Dilrukshi Herath, H.M.P.; Preston, S.; Hofmann, A.; Davis, R.A.; Koehler, A.V.; Chang, B.C.H.; Jabbar, A.; Gasser, R.B. Screening of a Small, Well-Curated Natural Product-Based Library Identifies Two Rotenoids with Potent Nematocidal Activity against Haemonchus contortus. Vet. Parasitol. 2017, 244, 172–175. [Google Scholar] [CrossRef]
- Swargiary, A.; Roy, B. In Vitro Anthelmintic Efficacy of Alpinia nigra and Its Bioactive Compound, Astragalin against Fasciolopsis buski. Int. J. Pharm. Pharm. Sci. 2015, 7, 30–35. [Google Scholar]
- Waldi, D. Spray Reagents for Thin-Layer Chromatography. In Thin-Layer Chromatography: A Laboratory Handbook; Bolliger, H.R., Brenner, M., Gänshirt, H., Mangold, H.K., Seiler, H., Stahl, E., Waldi, D., Stahl, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1965; pp. 483–502. ISBN 978-3-662-01031-0. [Google Scholar]
- Stévigny, C.; Block, S.; De Pauw-Gillet, M.C.; de Hoffmann, E.; Llabrès, G.; Adjakidjé, V.; Quetin-Leclercq, J. Cytotoxic Aporphine Alkaloids from Cassytha filiformis. Planta Med. 2002, 68, 1042–1044. [Google Scholar] [CrossRef] [PubMed]
- Rabel, B.; Mcgregor, R.; Douch, P.G.C. Improved Bioassay for Estimation of Inhibitory Effects of Ovine Gastrointestinal Mucus and Anthelmintics on Nematode Larval Migration. Int. J. Parasitol. 1994, 24, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.-F.; Nothias-Esposito, M.; da Silva, R.; Wang, M.; Protsyuk, I.; Zhang, Z.; Sarvepalli, A.; Leyssen, P.; Touboul, D.; Costa, J.; et al. Bioactivity-Based Molecular Networking for the Discovery of Drug Leads in Natural Product Bioassay-Guided Fractionation. J. Nat. Prod. 2018, 81, 758–767. [Google Scholar] [CrossRef] [PubMed]





| Sample/Control | H. contortus (%) | C. elegans (%) | Cytotoxicity, IC50 (µg/mL) |
|---|---|---|---|
| MeOH | 63.4 ± 0.8 | 70.0 ± 7.1 | >100 |
| 1 | - | - | 64.5 ± 5.7 |
| 2 | 29.8 ± 17.5 | 2.5 ± 4.3 | 58.9 ± 2.9 |
| 3 | - | - | 64.1 ± 2.2 |
| 4 | 15.1 ± 6.2 | 12.5 ± 4.3 | 60.3 ± 10.0 |
| 5 | 21.0 ± 3.9 | 22.5 ± 4.3 | 59.6 ± 4.9 |
| 6 | 35.7 ± 11.7 | 20.0 ± 7.1 | 78.9 ± 2.0 |
| 7 | 39.9 ± 7.9 | 22.5 ± 4.3 | >100 |
| 8 | 69.4 ± 10.4 | 75.0 ± 5.0 | >100 |
| 9 | 76.4 ± 1.8 | 85.0 ± 5.0 | >100 |
| DMSO (0.5%) | 3.1 ± 0.8 | 0 | - |
| LEV (25µM) | 100.0 ± 0.0 | 100.0 ± 0.0 | - |
| Compound (ID) | Equation | R2 | LOD (µg/mL) | LOQ (µg/mL) | F8 | F9 | ||
|---|---|---|---|---|---|---|---|---|
| µg/mL | µg/mg of Fraction | µg/mL | µg/mg of Fraction | |||||
| Ellagic acid (19) | y = 228815x + 926041 | 0.999 | 5.5 | 16.5 | 86.4 ± 6.9 | 8.6 ± 0.7 | 71.4 ± 2.6 | 7.1 ± 0.3 |
| Astragalin (23) | y = 45696x + 15964 | 0.997 | 3.3 | 9.9 | 7.7 ± 1.2 | 0.8 ± 0.1 | 9.6 ± 0.4 | 1.0 ± 0.0 |
| Gallic acid (8) | y = 8197.3x + 26875 | 0.995 | 10.3 | 31.1 | 96.5 ± 7.8 | 9.7 ± 0.8 | 19.9 ± 0.5 | 2.0 ± 0.5 |
| Compound (ID) | F8 | F9 | ||
|---|---|---|---|---|
| µg/mL | µg/mg of Fraction | µg/mL | µg/mg of Fraction | |
| Ellagic derivative (9) | 1.0 ± 0.4 | 0.1 ± 0.0 | 13.6 ± 1.1 | 1.4 ± 0.1 |
| Ellagic derivative (10) | 2.1 ± 0.3 | 0.2 ± 0.0 | - | - |
| Ellagic derivative (11) | 4.2 ± 2.0 | 0.4 ± 0.2 | 35.9 ± 3.6 | 3.6 ± 0.4 |
| Ellagic derivative (12) | - | - | - | - |
| Ellagic derivative (13) | 2.4 ± 0.8 | 0.2 ± 0.1 | 0.5 ± 1.4 | 0.1 ± 0.2 |
| Ellagic derivative (14) | - | - | 8.0 ± 0.1 | 0.8 ± 0.0 |
| Ellagic derivative (15) | 0.3 ± 0.5 | 0.1 ± 0.1 | 0.4 ± 1.0 | <LOQ |
| Ellagic derivative (16) | 11.1 ± 0.6 | 1.1 ± 0.1 | 1.8 ± 0.2 | 0.2 ± 0.0 |
| Solvent System | Ratio | Volume (mL) |
|---|---|---|
| DCM-MeOH | 100–0 | 300 |
| DCM-MeOH | 99.5–0.5 | 400 |
| DCM-MeOH | 99–1 | 600 |
| DCM-MeOH | 98–2 | 600 |
| DCM-MeOH | 96–4 | 600 |
| DCM-MeOH | 94–6 | 1000 |
| DCM-MeOH | 92–8 | 600 |
| DCM-MeOH | 90–10 | 1000 |
| DCM-MeOH | 85–15 | 1200 |
| DCM-MeOH | 80–20 | 1200 |
| DCM-MeOH | 70–30 | 1200 |
| DCM-MeOH | 60–40 | 1200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tchetan, E.; Ortiz, S.; Olounladé, P.A.; Hughes, K.; Laurent, P.; Azando, E.V.B.; Hounzangbe-Adote, S.M.; Gbaguidi, F.A.; Quetin-Leclercq, J. Fractionation Coupled to Molecular Networking: Towards Identification of Anthelmintic Molecules in Terminalia leiocarpa (DC.) Baill. Molecules 2023, 28, 76. https://doi.org/10.3390/molecules28010076
Tchetan E, Ortiz S, Olounladé PA, Hughes K, Laurent P, Azando EVB, Hounzangbe-Adote SM, Gbaguidi FA, Quetin-Leclercq J. Fractionation Coupled to Molecular Networking: Towards Identification of Anthelmintic Molecules in Terminalia leiocarpa (DC.) Baill. Molecules. 2023; 28(1):76. https://doi.org/10.3390/molecules28010076
Chicago/Turabian StyleTchetan, Esaïe, Sergio Ortiz, Pascal Abiodoun Olounladé, Kristelle Hughes, Patrick Laurent, Erick Virgile Bertrand Azando, Sylvie Mawule Hounzangbe-Adote, Fernand Ahokanou Gbaguidi, and Joëlle Quetin-Leclercq. 2023. "Fractionation Coupled to Molecular Networking: Towards Identification of Anthelmintic Molecules in Terminalia leiocarpa (DC.) Baill" Molecules 28, no. 1: 76. https://doi.org/10.3390/molecules28010076