1. Introduction
Inflammatory bowel diseases (IBDs), such as ulcerative colitis (UC) and Crohn’s disease (CD), are characterized by low-grade chronic inflammation and immune dysregulation of the intestine with alternating relapses and remission and require long-term treatments [
1]. Its etiology is unknown, although it is considered a group of complex heterogeneous gastrointestinal diseases involving genetic variations, intestinal microbiota, immunological factors, and environmental factors, which interact with an inherent genetic predisposition [
2,
3]. The interaction between these factors leads to the disruption in the balance between pro- and anti-inflammatory components and are among the key causes of intestinal inflammation in IBD [
4]. Inflammation is often accompanied by oxidative stress caused by overproduction of reactive oxygen and nitrogen species, which play central roles in the pathophysiology of IBD [
5]. The products of oxidative stress along with intensifying inflammation contribute to the destruction of cell structure by lipid peroxidation, eventually causing cell apoptosis and necrosis. Thus, oxidative stress and inflammation were shown to be key players in the pathogenesis of IBD and represent important therapeutic targets for IBD [
5,
6,
7,
8,
9].
The available therapies for IBD are far from satisfactory for the majority of patients who require continuous medication to keep the disease under control [
10]. In addition, the currently used agents including amino salicylates, corticosteroids, immunomodulators, and biological agents often have various side effects with variable responses and can diminish over time as well as limit their therapeutic benefits [
11]. To date, no cure aside from radical surgery for refractory ulcerative colitis and no preventive measures are available for these conditions. The use of plants in disease management goes back thousands of years and is based on traditional consumption in diets for medicinal benefits [
12,
13]. In recent years, numerous plant extracts and phytochemicals have been shown to ameliorate intestinal inflammation through different molecular pathways, including anti-inflammatory and immunoregulatory mechanisms, antioxidative properties, and modulation of intracellular signaling transduction pathways [
8,
9,
12,
13,
14,
15]. The use of plant-based drugs as complementary and alternative medicines has gained popularity among patients with IBD (30–50%), not only due to their dissatisfaction with the existing IBD therapies but also due to their pharmacological properties attributing to the attenuation of oxidative stress and inflammation as well as correcting nutritional deficits and preventing complications [
13,
16,
17].
Among the numerous plants used in the treatment of intestinal inflammatory diseases, a plant, Lycopodium (LYCO) has garnered attention due to its potent multiple properties including immunomodulation. LYCO (synonymous Huperzia), family
Lycopodiaceae, a plant of tropical and subtropical forests, is also known by other names such as club moss, or devil’s claw or ground pine, and constitutes an important component in traditional medicine for renal diseases, rheumatic arthritis, cystitis, and gastritis [
18]. LYCO has been demonstrated to possess anticancer [
18,
19], antioxidant [
20,
21], and anti-inflammatory [
22,
23] properties. LYCO has been well characterized to contain alkaloids and triterpenoids as major constituents, and these compounds have received attention for their potential pharmacological properties [
24,
25]. To test its therapeutic potential in gastrointestinal diseases, LYCO nano-formulations have been observed experimentally to exhibit potent anticancer activity against colon cancer by inhibiting cell growth and enhancing cell death [
19,
25]. A recent preliminary study demonstrated that the constituents of
Lycopodium clavatum elicit potent anti-inflammatory activity, suggesting its usefulness in IBD [
24,
25]. In another study, LYCO showed potent immunomodulatory actions on digestive pathogenic progression in Chagas disease wherein the infection involved cytokine-mediated inflammatory reactions [
22]. There is no report available to date on the effect of LYCO in ulcerative colitis. Thus, considering the antioxidant and anti-inflammatory role of LYCO, it is worthwhile to investigate the efficacy of LYCO against colitis induced in rats using acetic acid (AA), a classical model widely employed to imitate the pathophysiologic process of IBD and investigate preventive and therapeutic agents [
8,
26].
In this study, the protective effects of LYCO on the macroscopic and microscopic pathologies were investigated in the rat colon. Additionally, the activities of antioxidant enzymes and the levels of proinflammatory cytokines and calprotectin were measured in rat colon tissues.
2. Experimental Animals
Before conducting the animal experiments, written permission (animal ethics approval no. ERA_2019_6000) was obtained from the Institutional Animal Ethical Committee of the United Arab Emirates University, and all experimental procedures were carried out in accordance with ethical guidelines on animal studies. Male adult albino Wistar rats, weighing 225–240 g, were housed in polycarbonate cages at standard animal house conditions viz. photoperiod of 12 h light/dark cycle was used for the experiments. The room temperature ranged between 21 and 24 °C and relative humidity between 40 and 60%. The animals were maintained at the animal research facility, located at the College of Medicine and Health Sciences, United Arab Emirates University, Al Ain, United Arab Emirates. The animals were fed commercially available standard chow diet and fed water ad libitum. They were randomly divided into different animal groups and acclimatized for a week before commencing the experiments. Prior to the induction of colitis, the animals were fasted for 24 h. To induce colitis, 1 mL of 4% AA was administered intrarectal at 8 cm proximal to the anus for 30 s under the influence of light anesthesia with ether. To prevent the leakage of the solution, the rats were kept in Trendelenburg position during rectal instillation for a minute. One ml of phosphate-buffered saline was also administered to flush the colon in similar conditions.
2.1. Chemicals and Kits
Acetic acid (AA) was obtained from BDH Prolabo (Johannesburg, South Africa). The test drug, lycopodium, was procured from Sigma-Aldrich (St. Louis, MO, USA). The protein assay kit, BCA, and protease inhibitors were procured from Thermo Fisher Inc. (Waltham, MA, USA). Malondialdehyde assay kit and MPO kit were obtained from Hycult Biotech (Uden, The Netherlands) and Northwest Life Science (Vancouver, WA, USA). The IL-1β kit DuoSet and GSH assay kit were obtained from R&D Systems (Minneapolis, MN, USA) and Sigma-Aldrich (Sigma Chemical Co., St. Louis, MO, USA), respectively. The kits for calprotectin and IL-23 were obtained from MyBioSource Inc. (San Diego, CA, USA). Sodium deoxycholate and all other chemicals that were not specified were obtained from Sigma-Aldrich (St. Louis, MO, USA).
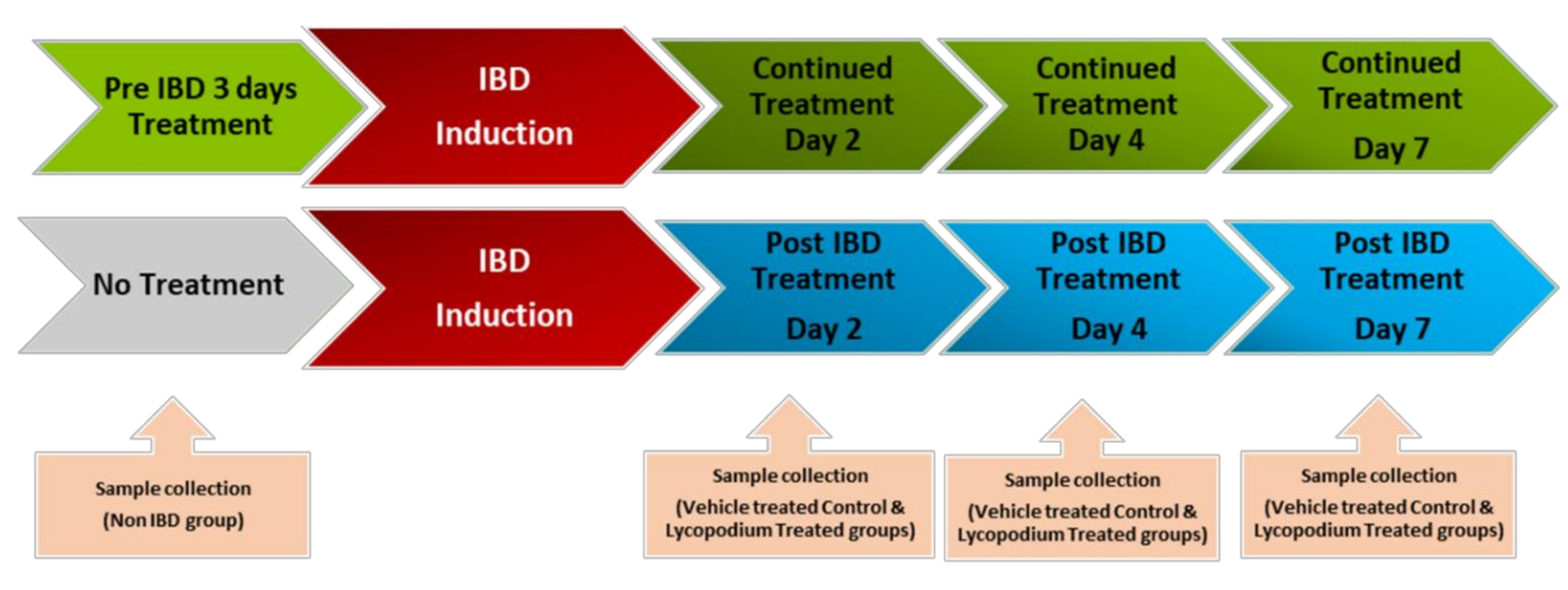
2.2. Experimental Design
The schematic representation of the study design is presented in
Figure 1. The doses of LYCO were chosen on the basis of a pilot study-assessed dose response from our previously published laboratory studies [
20]. The dose of 50 mg/kg prepared in olive oil was administered orally once a day using an intragastric gavage. The rats were lightly anaesthetized with ether after overnight fasting, and 1 mL of 4% AA was used to induce IBD.
LYCO was given under two different regimes: (1) a therapeutic regimen wherein LYCO effects were tested for 1 week after the induction of IBD and (2) a preventive regimen wherein LYCO was given 3 days before induction of IBD and thereafter for 7 days. Briefly, for each dosing regimen, rats were grouped into pre-IBD LYCO-treated (n = 18) and post-IBD LYCO-treated groups (n = 18). LYCO was given orally every day 30 min after the induction of IBD assigned as post-treated groups. The same protocol was used in control animals (olive oil alone). In assessing the protective effect of LYCO on IBD, the animals that were administered LYCO 3 days before induction of IBD were assigned as pretreated groups.
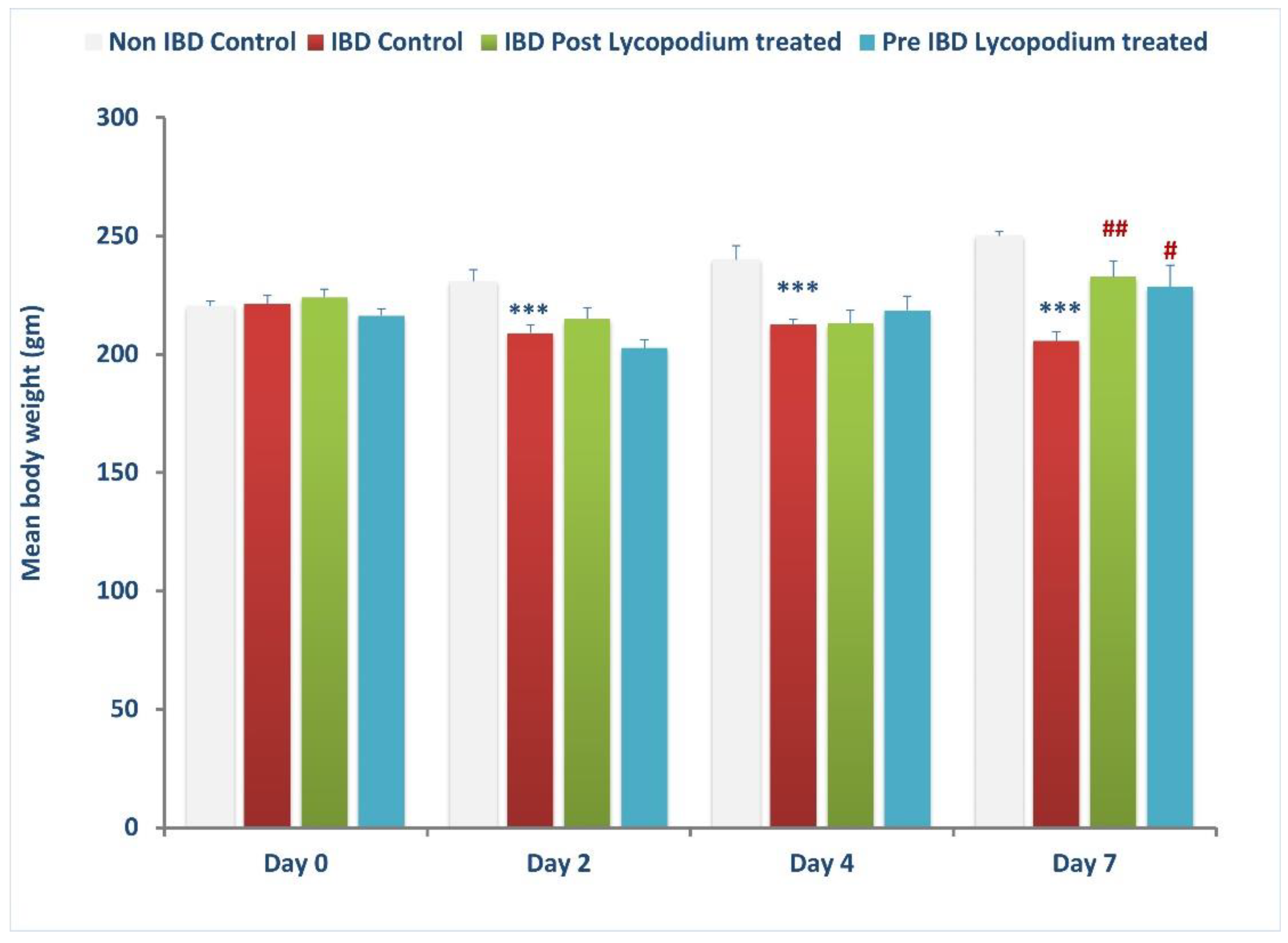
The weight of the animals was taken on days 0 (baseline), 2, 4, and 7 after IBD, and the colon was collected from each group, control, post-treatment, and pre-treatment, and processed for macroscopic, biochemical, and histological assessment and stored accordingly in liquid nitrogen or buffered formalin (4%).
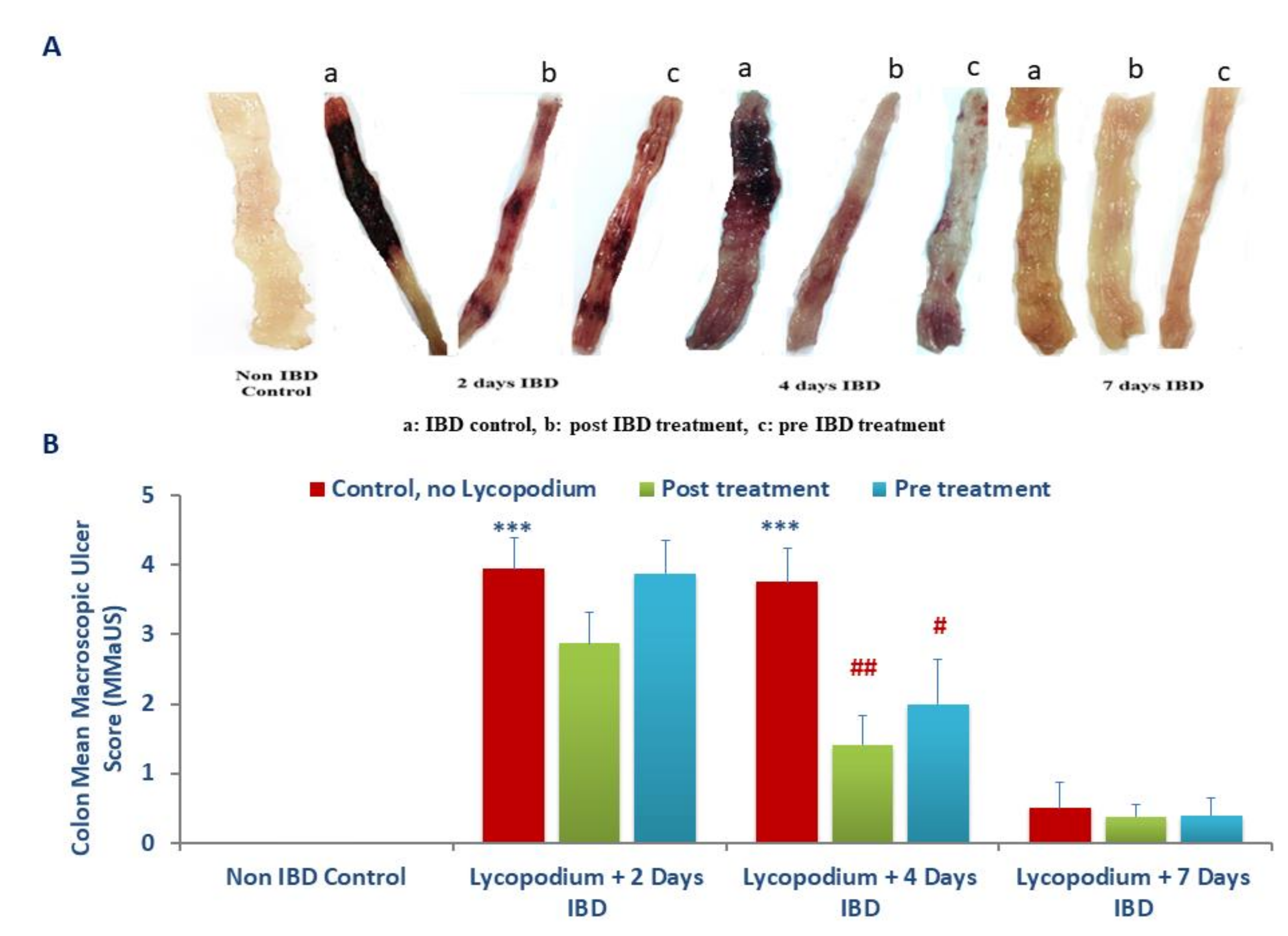
2.3. Assessment of Macroscopic Ulcer Score
The colon damage was assessed by macroscopic and microscopic observations. The samples for histological analysis were collected at days 0 (baseline), 2, 4, and 7 days of IBD with or without LYCO treatment. The rats were euthanized by cervical dislocation after 7 days of IBD. The colon was excised 2 cm above the anal margin following longitudinal opening and washed with saline. Macroscopic injury was examined based on scores following the method described by Jo et al. [
25], wherein the area of inflammation and the presence or absence of ulcers were observed [
27].
The assessing standards for macroscopic injury followed semiquantitative score wherein the observations were graded numerically as follows: no ulcer, no inflammation (0); no ulcer, local hyperemia (1); ulceration without hyperemia (2); ulceration and inflammation one site only (3); ulceration and inflammation on two or more sites (4); ulceration extending more than 2 cm (5).
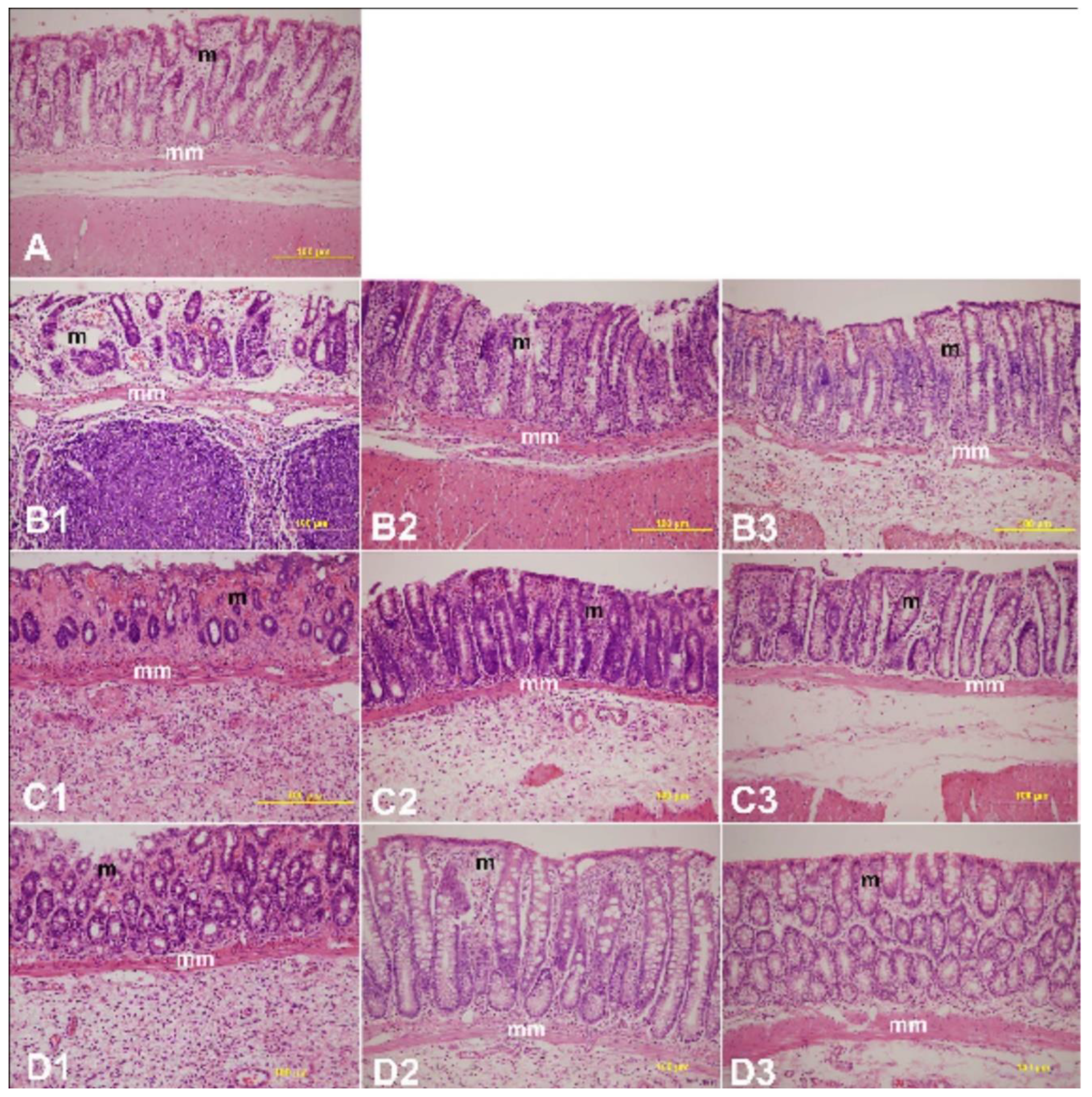
2.4. Histopathology Study
The colon histology was examined after staining with hematoxylin and eosin on days 0, 2, 4, and 7 after IBD induction in rats treated with or without LYCO. Colon samples were retrieved from rats of each experimental group and processed according to the standardized method. In brief, the rats used for the experiment were sacrificed following cervical dislocation, and surgery was performed to remove the colon in proximity to the anal opening.
The colon samples were trimmed of adherent connective tissue and fixed in phosphate-buffered saline-buffered formalin (4%) for a week. They were later washed in tap water for 2 h, dehydrated in graded alcohol, and paraffin embedded following a previously described method [
28]. Using a microtome, the tissue was sectioned (5 µm) and examined after hematoxylin-eosin staining. The scoring system adapted from a previously described method was used to grade the degree of mucosal damage in LYCO-treated IBD rats compared to untreated controls [
29].
2.5. Preparation of Colon Tissue Homogenate
The colon samples of 8 cm length were opened longitudinally and rinsed well with chilled phosphate-buffered saline. After weighing the tissue, homogenization was performed using 28 mm ceramic beads by a bead raptor homogenizer (Omni International, Kennesaw, GA, USA). The homogenizing buffer was ten volumes of chilled high potassium chloride lysis buffer (10 mM Tris-hydrochloric acid, pH 8.0, 140 M sodium chloride, 1M EDTA, 0.5% sodium deoxycholate, 300 mM potassium chloride, and 0.5% Triton X-100) with protease inhibitor cocktail. The homogenates were incubated for 30 min on ice and thereafter centrifuged at 4 °C for 30 min with 15,000 rpm. The obtained supernatant was kept at −40 °C until ELISA was performed. The concentration of protein in each sample was measured using a commercially available kit following a bicinchoninic acid (BCA) protein assay kit (Pierce, Thermo Scientific, Rockford, IL 61101, USA). The content of test markers was normalized to protein content and reported as per mg protein.
2.6. Determination of Myeloperoxidase (MPO)
Myeloperoxidase (MPO) activity was determined following the manufacturer’s instructions of sandwich ELISA kits. Briefly, at room temperature, a 96-well microtiter plate, coated with primary antibody, the sample (100 µL), and standards (100 µL) were added for 1 h. After washing, the biotinylated trace antibody was added at a volume of 100 µL in each well for one hour at room temperature. Following washing to bind/interact with biotinylated trace antibodies, the streptavidin-peroxidase conjugate was added for 1 h at room temperature. The plate was again washed to add TMB-ELISA substrate for 30 min at room temperature. Oxalic acid was added to stop the enzyme reaction. After adding a stop solution, the absorbance was measured at 450 nm using a microplate reader (Tecan Group Ltd., Männedorf, Switzerland). The data were presented as ng/mg of colonic protein.
2.7. Determination of Calprotectin
Calprotectin in colon samples was determined using a commercially available sandwich ELISA kit (MyBioSource, San Diego, CA, USA). Briefly, to a coated 96 microtiter plate, samples (50 µL) and standards (50 µL) and HRP (100 µL) were added for an hour at 37 °C. Following washing and addition of TMB-ELISA substrate, the absorbance was measured at 450 nm using a microplate reader (Tecan Group Ltd., Männedorf, Switzerland), after adding a stop solution. Calprotectin levels were presented as ng/mg of protein.
2.8. Measurements of Reduced Glutathione (GSH), Superoxide dismutase (SOD), and Catalase
Reduced glutathione (GSH) content in colon homogenates was determined following the manufacturer’s instruction of the commercially available kits. To a coated 96-well microtiter plate, the sample and standards were added for 30 min at 37 °C. The absorbance was measured at 450 nm using a microplate reader (Tecan Group Ltd., Männedorf, Switzerland). The data were presented as µM/mg of tissue. SOD assay was performed according to the manufacturer’s protocol. This colorimetric assay utilized the tetrazolium salt for detection of superoxide radicals generated by xanthine oxidase and hypoxanthine. One unit of SOD enzyme was required to inhibit 50% dismutation of the superoxide radical. The inhibition of color development was measured at 450 nm by using Emax Plus microplate reader (Molecular Devices, San Jose, CA 94089, USA). Results are expressed in units per mg of protein. CAT levels were measured by Catalase Assay Kit which utilized the peroxidatic function of CAT for determination of enzyme activity. The method was based on the reaction of the enzyme with methanol in the presence of an optimal concentration of hydrogen peroxide. The formaldehyde produced was measured spectrophotometrically at 540 nm, with 4-amino-3-hydrazino-5-mercapto-1,2,4-trizazole as the chromogen by using EmaxPlus microplate reader (Molecular Devices, San Jose, CA 94089, USA). CAT activity in tissue homogenates is expressed as nmol/min/mg protein.
2.9. Measurement of Malondialdehyde (MDA)
Malondialdehyde (MDA) in the colon samples was measured using a commercially available assay kit based on the principle that MDA reaction with thiobarbituric acid (TBA) forms an adduct: MDA-TBA with absorbance on 532 nm. Briefly, the sample of the deproteinated tissue was mixed with a solution containing ethanol, butylated hydroxytoluene, and phosphoric acid and the resultant mixture was subjected to heat for 60 min at 60 °C. Furthermore, the mixture was kept at room temperature for 15 min. Then, the mixture was centrifuged at 10,000× g for 3 min and the pink color supernatant was collected and the optical density was measured at 532 nm. The concentration of MDA is presented as µM.
2.10. Determination of Inflammatory Cytokines: IL-1β and IL-23
The determination of IL-1β in colon homogenate was carried out by a commercial DuoSet ELISA kit (R&D Systems Inc., Minneapolis, MN, USA). Briefly, the test samples and standards were added to a primary antibody coated 96-well microtiter plate for 30 min at 37 °C. Subsequently, TMB-ELISA substrate was further added, and the resultant mixture’s optical density was read at 450 nm using a microplate reader (Tecan Group Ltd., Männedorf, Switzerland). IL-1β levels are presented as pg per milligram of protein. Similarly, the level of IL-23 in the colon was determined by a commercially available sandwich ELISA kit (MyBioSource, San Diego, CA, USA). Briefly, to the coated 96-well microtiter plates, the sample and standards were added and incubated for 2 h at 37 °C. The detection reagents, provided in the manufacturer kit labelled as reagent A was added after removal of the samples and incubated for 1 h at 37 °C. Next, the detection reagent B provided in the manufacturer kit was added after washing and incubated for 1 h at 37 °C. Following washing, the TMB-ELISA substrate and stop solution was added and the optical density of the resultant mixture was measured at 450 nm, using a microplate reader (Tecan Group Ltd., Männedorf, Switzerland). The levels of IL-23 are presented as pg per milligram of protein.
2.11. Statistical Analysis
The data are presented as mean with standard error of mean (SEM). The data were statistically analyzed using SPSS 23.0 software, and the analysis was performed using an independent t-test to determine the significance of the mean between the groups. The criterion of statistical significance was set on p < 0.05.
4. Discussion
The present study results show that LYCO treatment improved body weight, colonic length, attenuated oxidative stress, and inflammation. Furthermore, the preservation of colonic mucosa clearly indicates the protective and therapeutic effects of LYCO in AA-induced colitis in rats.
Medicinal plants exhibit a variety of pharmacological properties including antioxidant and anti-inflammatory, which shows beneficial effects in IBD. Although many treatment strategies showed beneficial effects for the treatment of IBD, long-term treatments often result in inadequate efficacy and adverse effects. An optimal drug is expected to reduce the disease progression with minimal adverse effects and improve prognosis of the disease. Therefore, in recent years, many research efforts have focused on identifying the health benefits of plants and plant-based medicines in gastrointestinal diseases including IBD [
9,
30]. This is the first report to demonstrate the effect of LYCO on experimental colitis by suppressing the inflammatory process in the AA-induced rat model of IBD. The AA-induced rat model of IBD is a convenient, economical, reproducible, and commonly used experimental model for evaluating the efficacy of potential drug candidates specifically targeting inflammation and oxidative stress [
31]. Enhanced vasopermeability and sustained neutrophil infiltration were seen with this model; in addition, there was an upregulation in inflammatory mediators akin to the pathogenesis of human IBD [
26]. The results showed that LYCO modulated mucosal immunity by bringing about a change in the integrity of the epithelial layer and colon injury following neutrophils and macrophage infiltration. In the pathogenesis, leukocytes and granulocytes migrate to the inflamed mucosal layers and the resultant induction and release of proinflammatory cytokines give rise to a hyperinflammatory state that affects the integrity of the colon mucosa and epithelial tissues [
32].
In the present study, administration of AA resulted in an increase in proinflammatory cytokines in colon tissues demonstrating the involvement of inflammation in IBD pathogenesis that was further confirmed by histopathology observations displaying neutrophil infiltration, swelling, and necrosis of the epithelial cells [
33]. The body weight and colon length of the rats are considered a consistent and sensitive marker of the onset and progression of inflammatory responses [
34]. In the current study, maintenance of colon length along with body weight and reduced diarrhea following treatment with LYCO is suggestive of its protective effect in colitis, in line with other studies wherein similar effects were observed with natural antioxidants [
8,
35,
36,
37].
Administration of AA resulted in severe colonic inflammation, which was characterized by colon thickening, goblet cell hyperplasia, crypt destruction, ulceration, necrosis following inflammatory cell infiltration and depletion of colonic mucus [
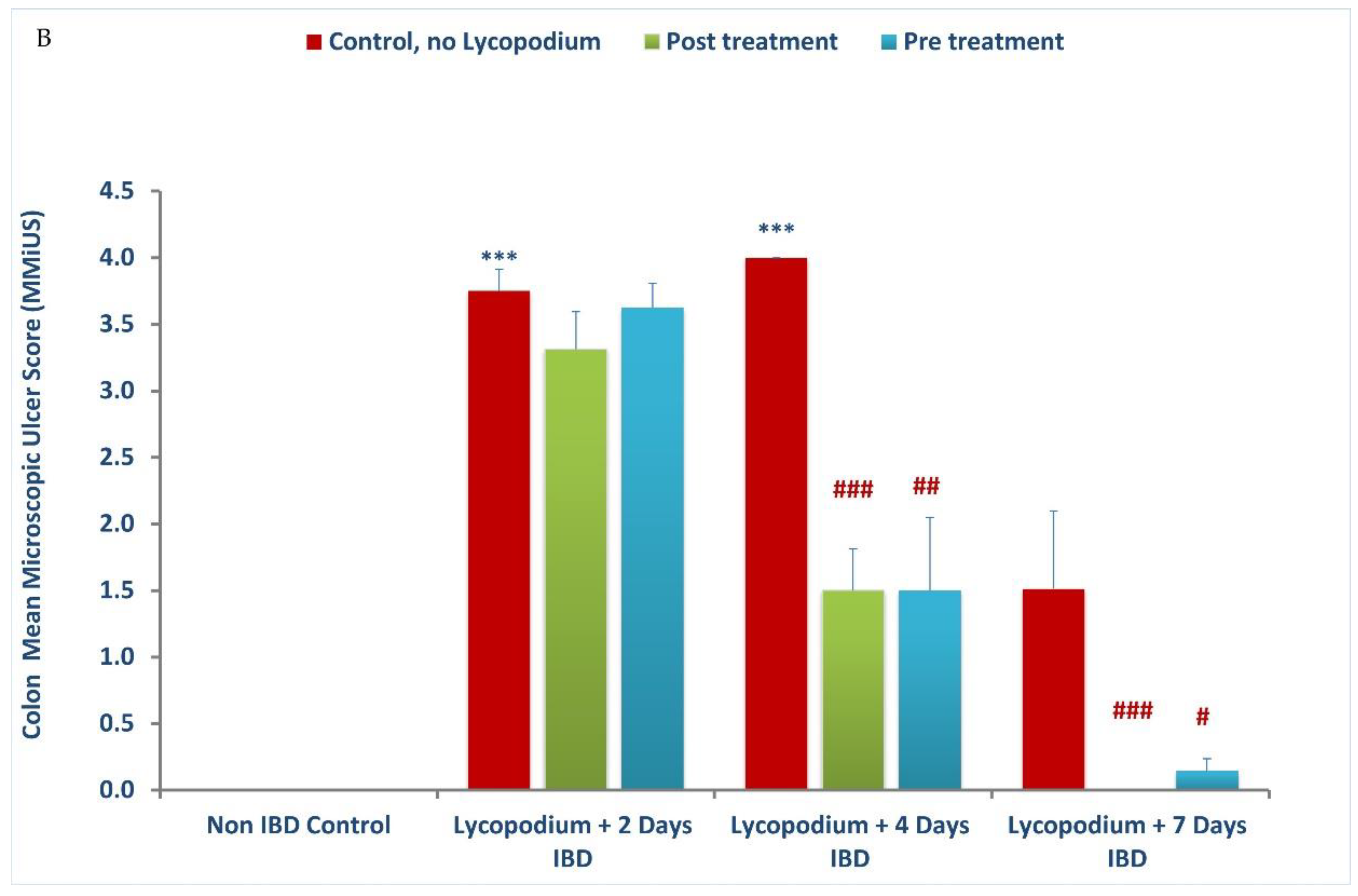
26,
38]. AA generates protons in the intracellular space leading to high acidification that results in epithelial injury. Treatment with LYCO attenuated the macroscopic and microscopic damages in the rat’s colon as observed in histological studies. The plausible reason for the reduced damage is the preservation of epithelial goblet cells, which is responsible for mucin production. Mucins are the predominant glycoproteins that form a barrier, which protect the bowel against antigens present in the lumen. The vascular mechanisms play an important role in promoting healing and protecting the mucosa in the gut [
39,
40]. Reduction in gastric blood flow triggers severe gastric ulceration, after the gastric mucosa is exposed to damaging factors [
39]. The injury caused to the intestinal mucosa in experimental IBD is convincingly associated with both increased production of free radicals and lesser availability of endogenous antioxidants [
6,
7].
Under normal conditions, the colonic mucosa has a weaker antioxidant defense relative to other organ systems [
41]. In colitis, generation of ROS overpowers the endogenous antioxidant network which keeps a check on free radicals, and ROS becomes imbalanced and leads to oxidative stress and DNA damage [
7,
8,
42]. The role of enzymatic and nonenzymatic antioxidants in protecting tissues from pro-oxidants is well known [
9]. The deranged balance in the colonic milieu impairs epithelial cell integrity impedes mucosal recovery and increases intestinal mucosal permeability [
34,
37]. These changes resulted in impaired function of the mucosal barriers and host defense following weakened endogenous antioxidant defense systems [
6,
8,
42]. The endogenous antioxidant network constitutes nonenzymatic antioxidant substrates, GSH, and enzymatic antioxidants such as SOD, CAT, and GPX to regulate the generation of free radicals and ROS [
34,
37]. These enzymes protect cells from free radicals by enhancing resistance to oxidative stress, reducing free radicals, and augmenting antioxidants. SOD is the key enzyme that converts oxygen radicals into oxygen and hydrogen peroxide. Furthermore, the enzymes, catalase, and glutathione peroxidase convert hydrogen peroxide into water [
34,
37]. GSH, an endogenous tripeptide antioxidant, is utilized as a substrate for the enzymatic conversion of hydrogen peroxide into water and oxygen by glutathione peroxidase and plays a critical role in detoxification, increasing amino acid transport, DNA synthesis, and repair [
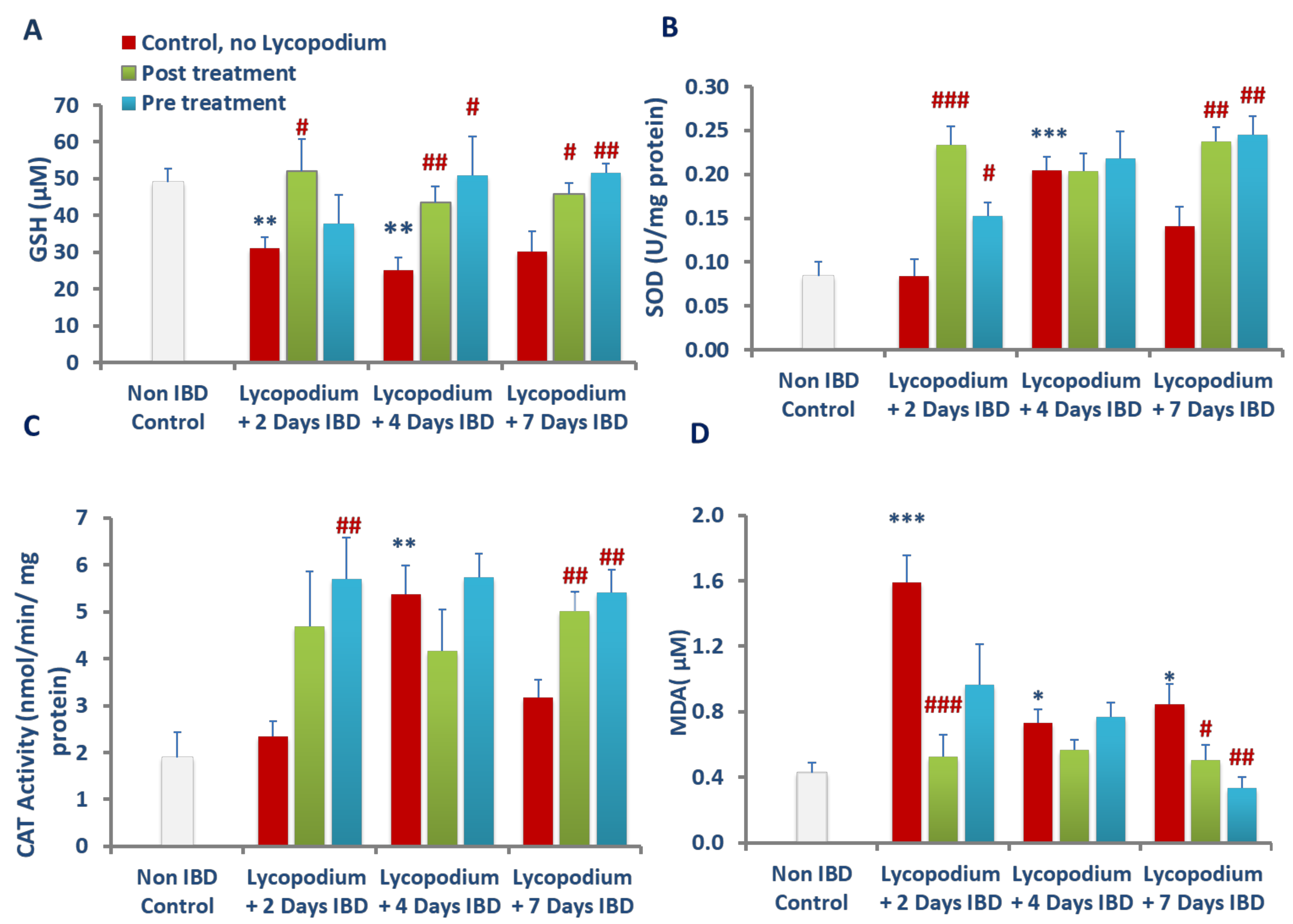
38]. In the present study, LYCO treatment improved SOD and catalase activities and normalized the levels of GSH in colon tissues that demonstrated that LYCO treatment inhibited the excessive generation of free radicals and restored GSH due to its antioxidant properties.
AA-injected animals displayed increased colonic levels of MDA, an indicator of lipid peroxidation following reduced antioxidant defense, and increased formation of lipid peroxides due to lipid peroxidation of polyunsaturated fatty acids in membranes [
43]. MDA formation is aggravated by the generation of cytotoxic free radicals destabilizing membrane lipids, proteins, and DNA and plays a role in IBD [
37,
43]. Furthermore, enhanced lipid peroxidation has been shown in AA-associated tissue injury, which is in line with the observations of the current study. In an earlier study, LYCO was showed to scavenge oxidizing radicals in brain, resulting in an organoprotective effect mediating antioxidant action [
20]. Therefore, it has been suggested that the antioxidant and antilipid peroxidation effects of LYCO afford protection from free radicals by restoring the endogenous antioxidants that result in reduced intestinal inflammation and damage in ulcerative colitis. The present study findings are in agreement with previous studies showing a protective effect of plant-derived antioxidants in attenuating inflammation and oxidative injury in experimental models of colitis [
35,
36,
37,
43].
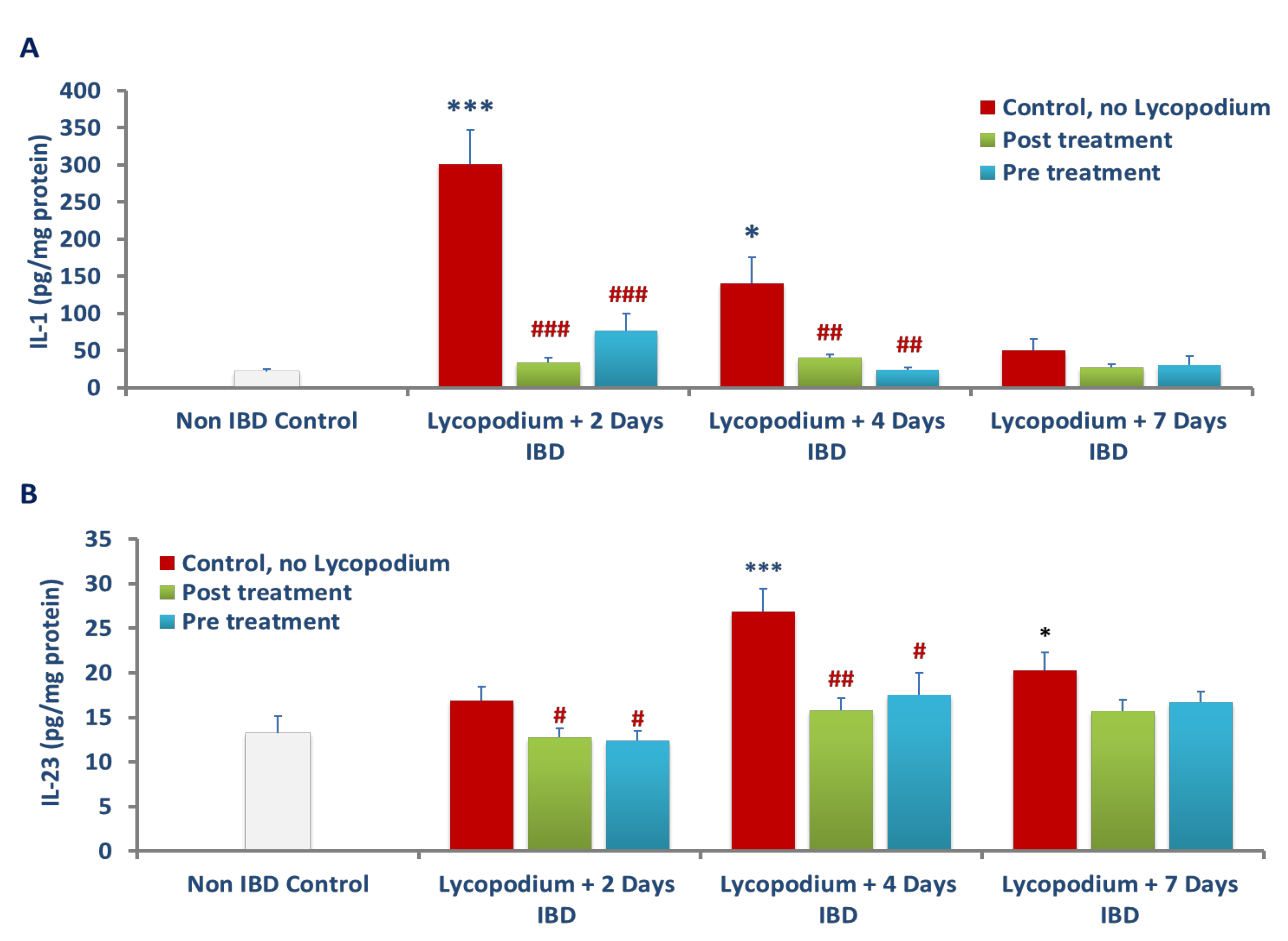
The accrual of ROS in ulcerative colonic tissues leads to the onset of inflammation by induction and release of proinflammatory cytokines, IL-1β and IL-23 [
8,
44,
45]. The elevation in the levels of IL-1β and IL-23 in colon tissues indicates the severity and extent of colitis [
44]. IL-1β is produced by monocytes and tissue macrophages and plays a critical role in the onset and progression of acute and chronic inflammation at both levels, local and systemic. The synthesis of IL-1β from its immature state is converted into an active state by caspase-1-mediated proteolytic reactions. The epithelial cells and macrophages in the colon release inflammatory cytokines, including IL-1β which contribute to the development of colitis by disrupting the intestinal epithelial barrier, inducing apoptosis of epithelial cells, promoting ROS production, activating neutrophils, and instituting inflammation [
44,
45]. IL-1β also stimulates the synthesis and release of inflammatory chemotactic proteases, cytokines, and chemokines by activating canonical pathways resulting in the activation of NF-kB, a redox-sensitive transcription factor usually in an inactive state with Ik-B its inhibitor. The stimulation of NF-kB depends on ROS and associated signaling cascades and participates in dereliction of its inhibitor Ik-B following phosphorylation and ubiquitination [
44,
45]. The NF-kB then activates translocating into the nucleus where its p65 subunit plays a key role in regulating genes controlling the immune-inflammatory cascade. Studies have shown that LYCO suppresses the activation of NF-kB prompted by various inflammatory mediators and exerts genoprotective effects against oxidative and methylation damage in HT-29 colorectal adenocarcinoma cells [
25]. In the present study, inhibition of proinflammatory cytokines such as IL-1β and IL-23 in colonic tissues following LYCO treatment demonstrates the anti-inflammatory effect, usually the first step in the treatment of UC.
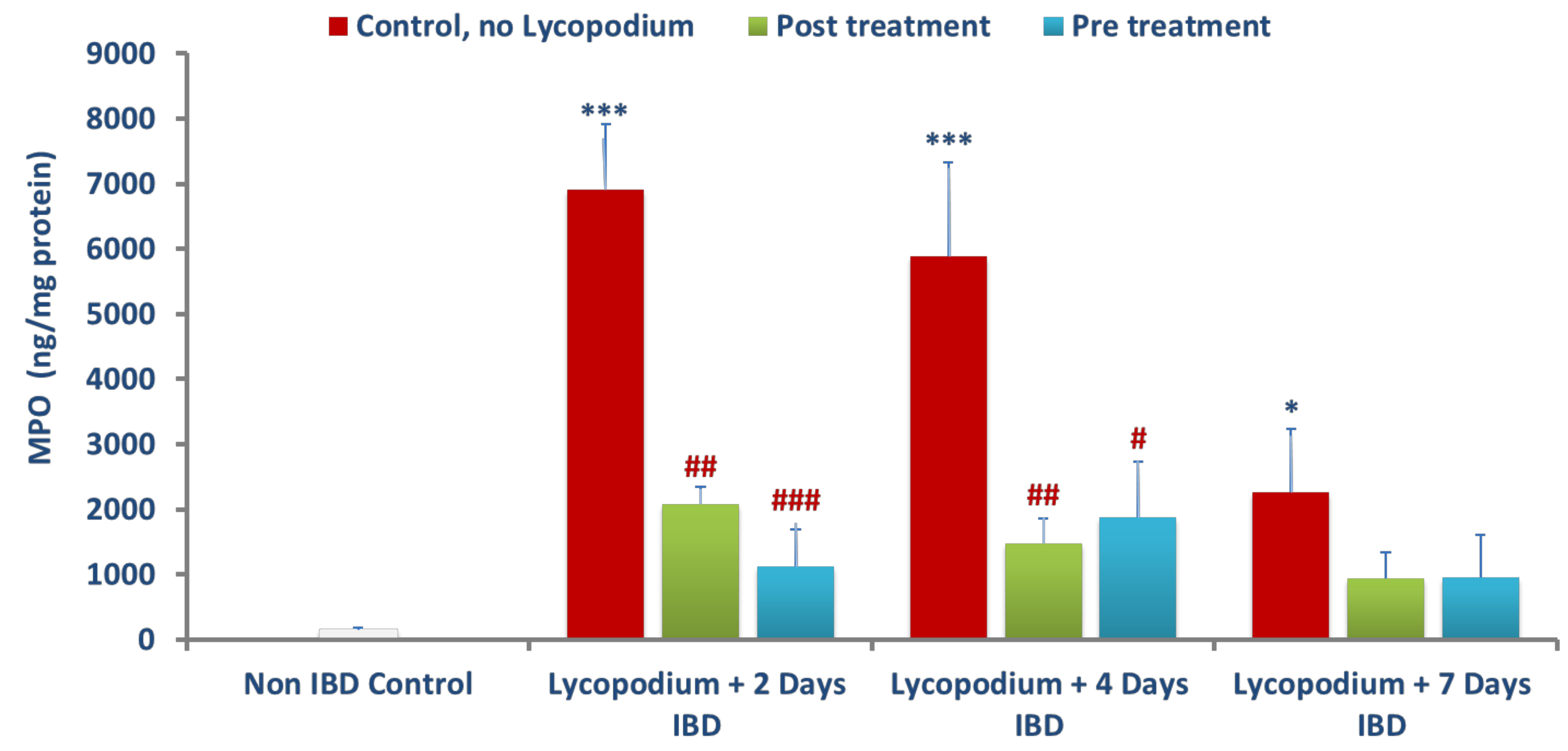
AA-induced damage to the colon involves neutrophil and macrophage infiltration, and the extent of tissue infiltration is proportionally reflected by the activity of an enzyme, MPO, a measure of neutrophil infiltration [
46]. This enzyme is mainly present in azurophilic granules of neutrophils with lesser concentration in monocytes and macrophages and serves as a sensitive and quantitative indicator for acute intestinal inflammation [
38,
41]. MPO is released from neutrophils during inflammation and facilitates the formation of hypochlorous acid and other free radicals, which exert antimicrobial properties. The free radicals formed cause injury to the host cells by targeting proteins, DNA, and lipids [
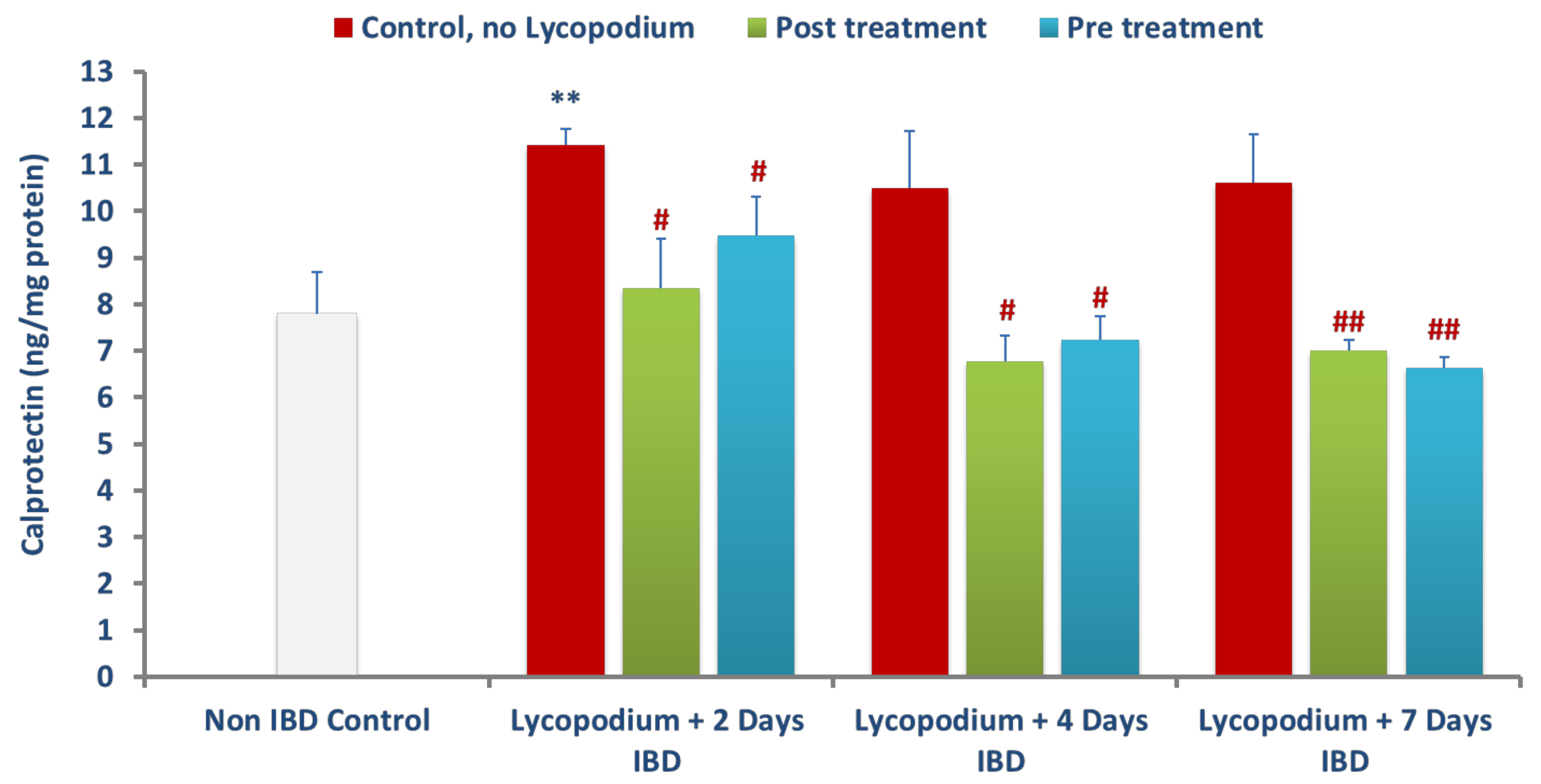
47]. Neutrophils exhibit a decisive role in the etiopathogenesis of colitis by increasing ROS and superoxide anions that further exacerbate the formation of hydroxyl radicals and peroxide leading to mucosal defense impairment and necrosis. LYCO treatment exhibits a decrease in polymorphonuclear infiltration demonstrated by a significant reduction in MPO activity. In addition to MPO, there was a significant rise in the levels of calprotectin, a protein found in the cytosol of inflammatory cells, following activation of leukocytes that display intestinal inflammation and progression of colitis [
48]. Reductions in the levels of calprotectin after treatment with LYCO demonstrate the anti-inflammatory property of LYCO against intestinal inflammation.
The histopathological studies support the macroscopic and biochemical observation on the protective effects of LYCO in colitis. Rectal administration of AA was found to cause tissue ulceration, inflammatory cell infiltration, necrosis, and goblet cell hyperplasia along with reduction in colon length. These observations, which characterize IBD, appear consistent with our previous studies in AA-induced colitis in rats [
37]. Pre and post treatment with LYCO significantly improved the colon histoarchitecture by preserving the microscopic and macroscopic structures of the colon and preventing depletion of the colonic wall mucus. Leukocyte infiltration, edema, and tissue injury, which are histological signs of inflammation, appear alleviated following pre-treatment with LYCO. The observation of the present study wherein LYCO protects against colitis is in support of earlier results in its ability to inhibit inflammation and oxidative stress in brain of rats [
20].
Furthermore, inhibition of NF-kB activation makes it a potentially effective suppressor of cell growth, proliferation, and inflammation [
18]. LYCO species are known to contain characteristic quinolizine or pyridine and alfa-pyridone-type alkaloidal and serratene triterpenoid constituents which contribute to the potent antioxidant and anti-inflammatory properties [
25,
49]. Alkaloids are a class of secondary metabolites in plants containing amino acid-derived nitrogen-containing organic compounds with low molecular weight. Recently, a review highlighted that alkaloid are beneficial in intestinal inflammatory disorders and may provide promising molecules for prophylaxis and treatment of IBD and offer hope for further pharmaceutical development [
50]. Similarly, mainly serratene triterpenoids found in LYCO have been shown to exhibit a variety of potent pharmacological properties including antioxidant and anti-inflammatory properties [
49]. The serratene triterpenoid compound isolated from LYCO has been recently shown to inhibit inflammatory mediators including enzymes and proinflammatory cytokines in HT-29 cells [
25]. It can be reasonably speculated that the triterpenoids and alkaloids attribute to the potential benefits of LYCO in IBD.
In alternative medicine, the whole plant formulation is considered an effective therapeutic strategy because of their constituents acting synergistically and nutritional properties in chronic low-grade inflammatory diseases including IBD [
12]. This concept of using extract particularly reveals a new class of agents, termed as adaptogens, that promote resistance to aversive stimuli perturbing internal homeostasis. The adaptogenic agents have the potential to favorably modulate stress-induced immune and organ impairment by sparing antioxidants and immunomodulation with lesser adverse effects. The plant extract has been found useful for motor abnormalities in children [
51]. The use in children is suggestive of the relative safety of LYCO. The results of the present study demonstrate the therapeutic potential of LYCO in IBD and demonstrated the anti-inflammatory and anticolitis activities of LYCO in the experimental model of acute colitis. LYCO administered via the oral route appears effective in numerous disease models and bioavailable with negligible adverse effects that further indicate its potential for nutritional development and its ease of using orally. The results of the present study demonstrate the anti-inflammatory and anticolitis activities of LYCO in the experimental model of acute colitis.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








