
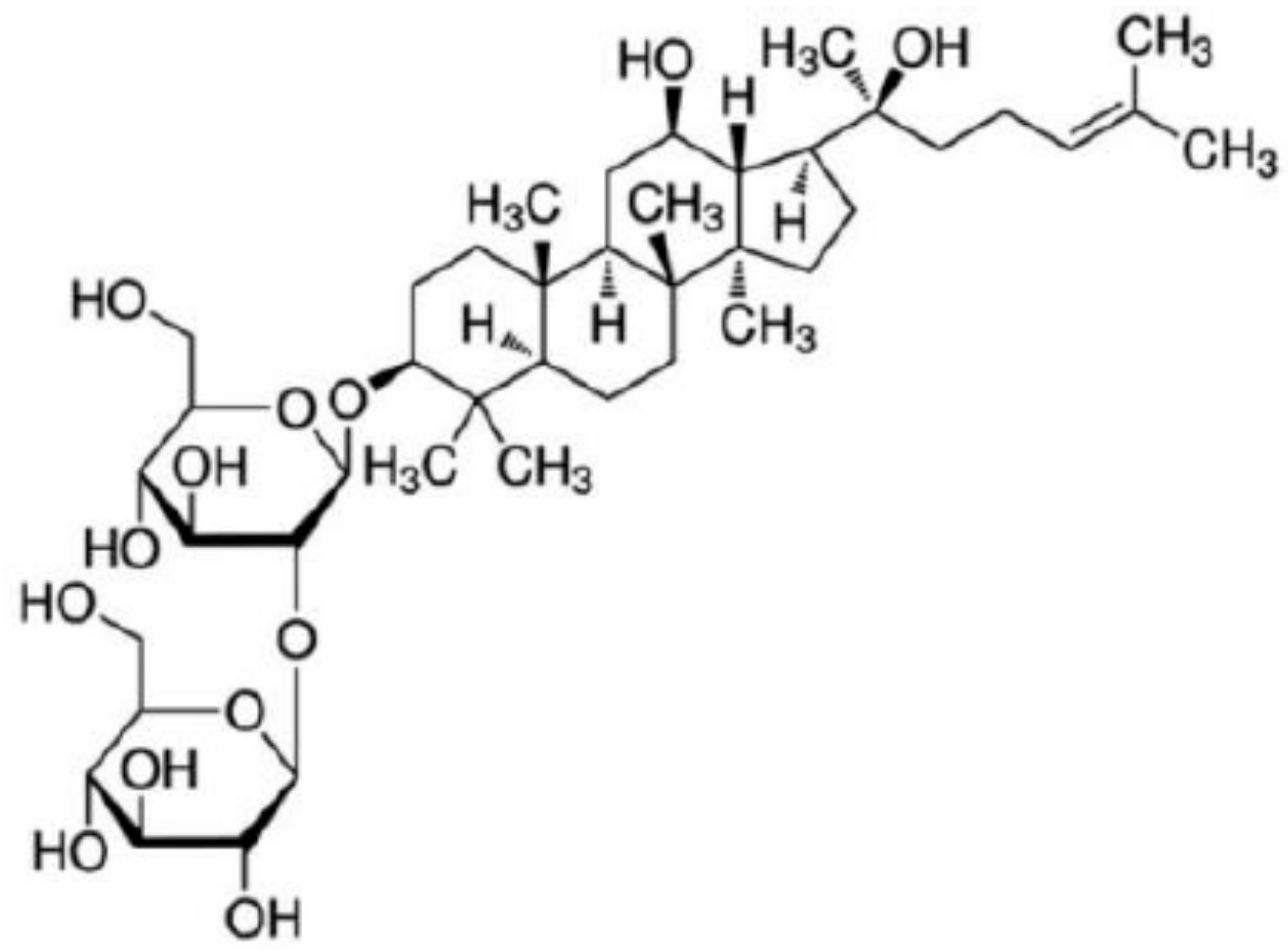
Korean Red Ginseng and Ginsenoside Rg3 Suppress Asian Sand Dust-Induced Epithelial–Mesenchymal Transition in Nasal Epithelial Cells



Abstract
:1. Introduction
2. Results
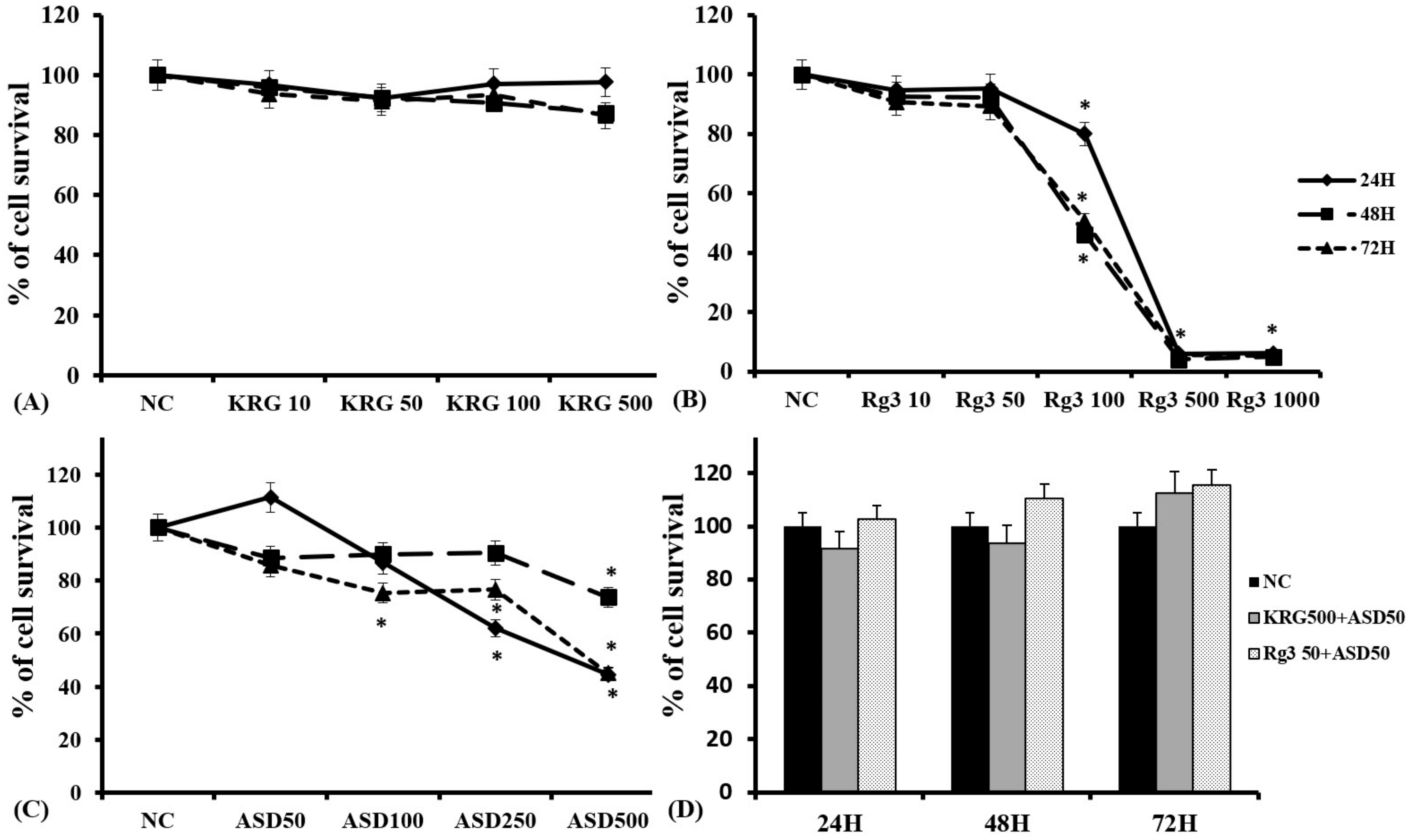
2.1. Cytotoxicity of ASD, KRG, and Ginsenoside Rg3 in Nasal Epithelial Cells
2.2. Effects of KRG and Rg3 on ASD-Induced Production of IL-8 and TGF-β1
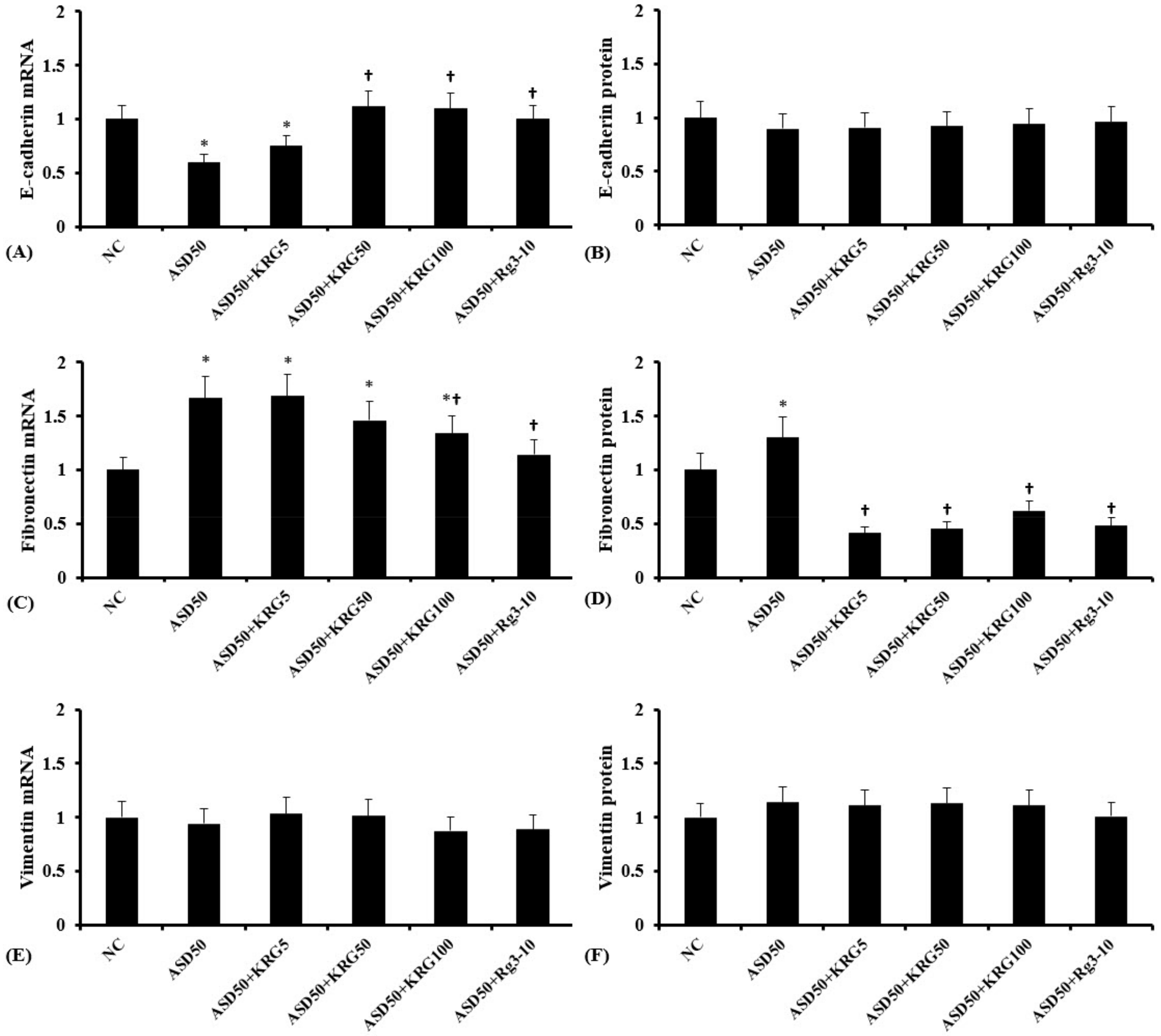
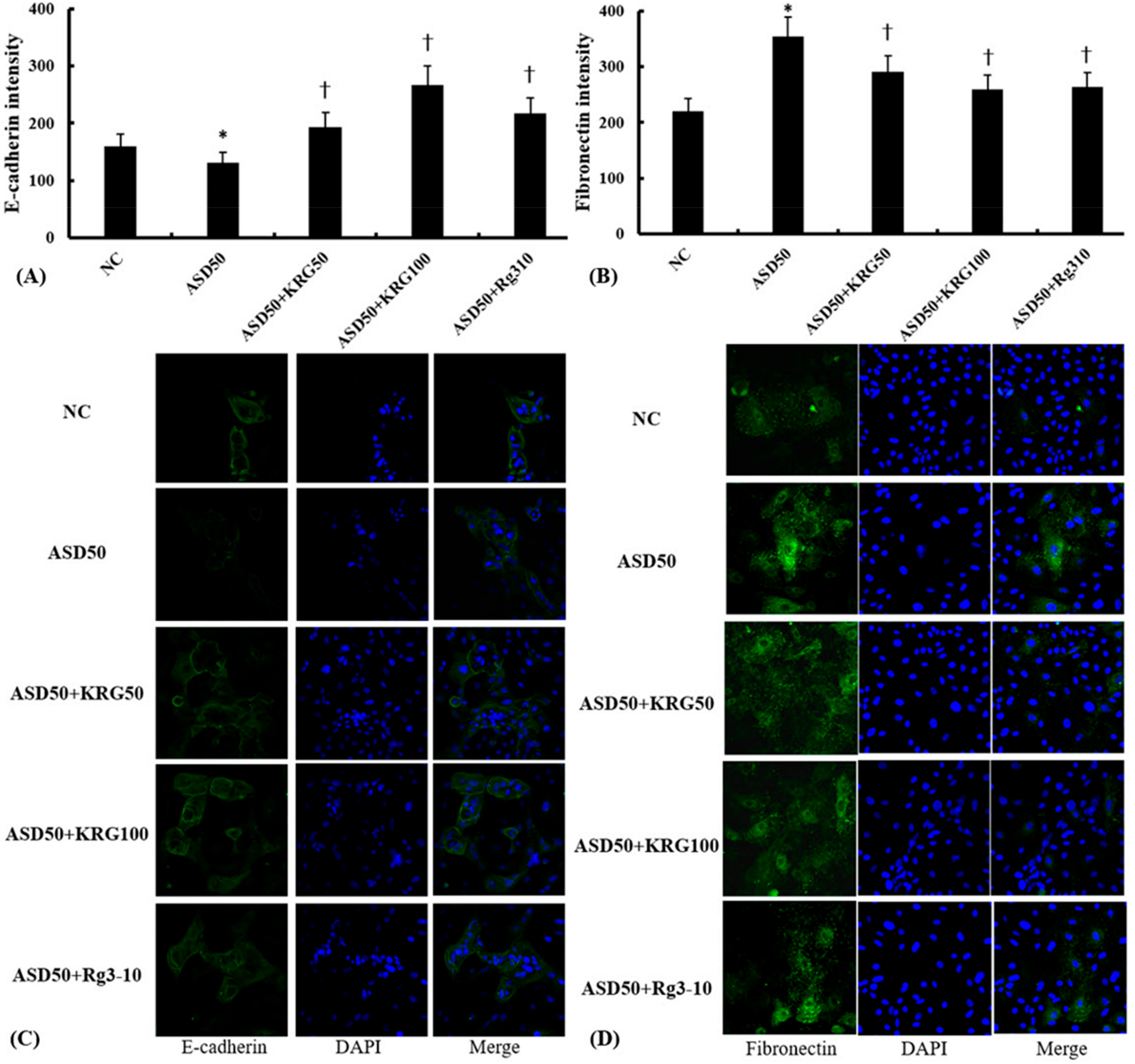
2.3. Effects of KRG and Rg3 on ASD-Induced EMT Marker mRNA and Protein Expression
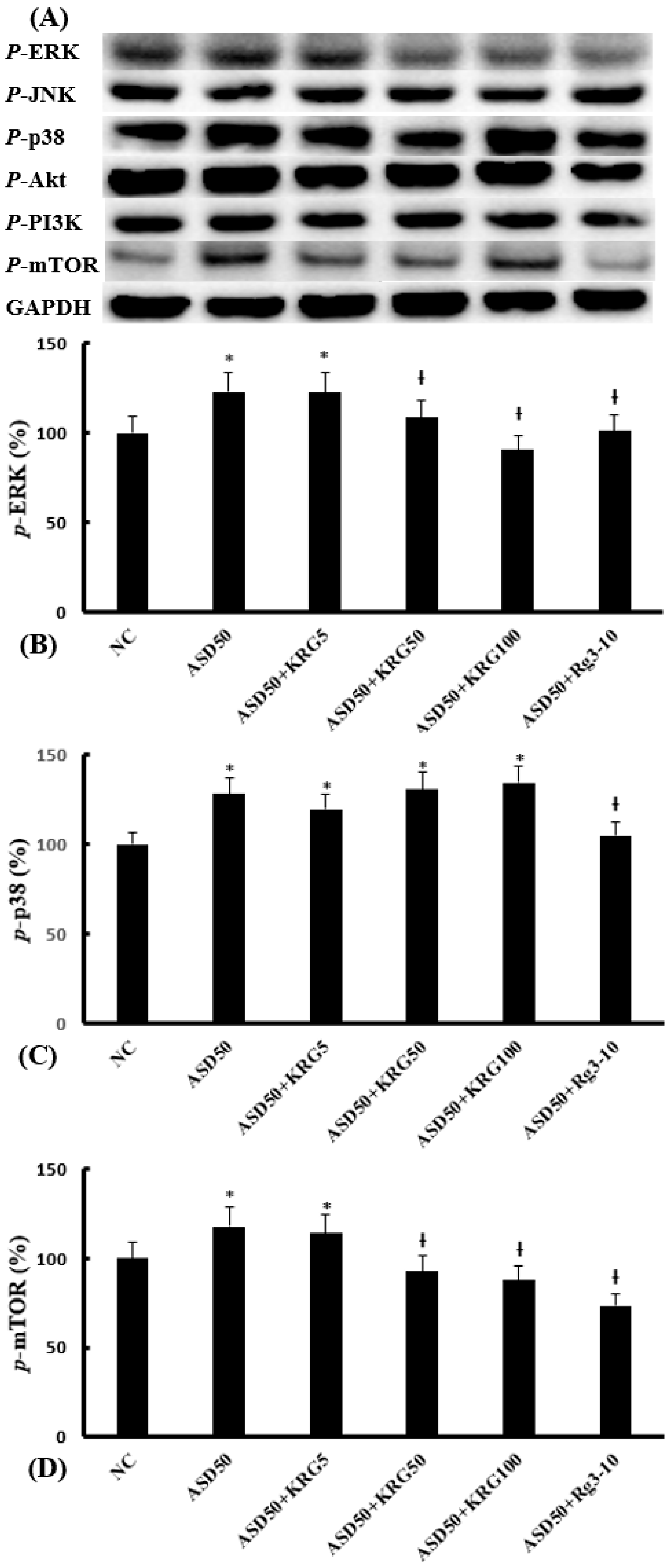
2.4. Effects of KRG and Rg3 on ASD-Induced Transcription Factor Expression
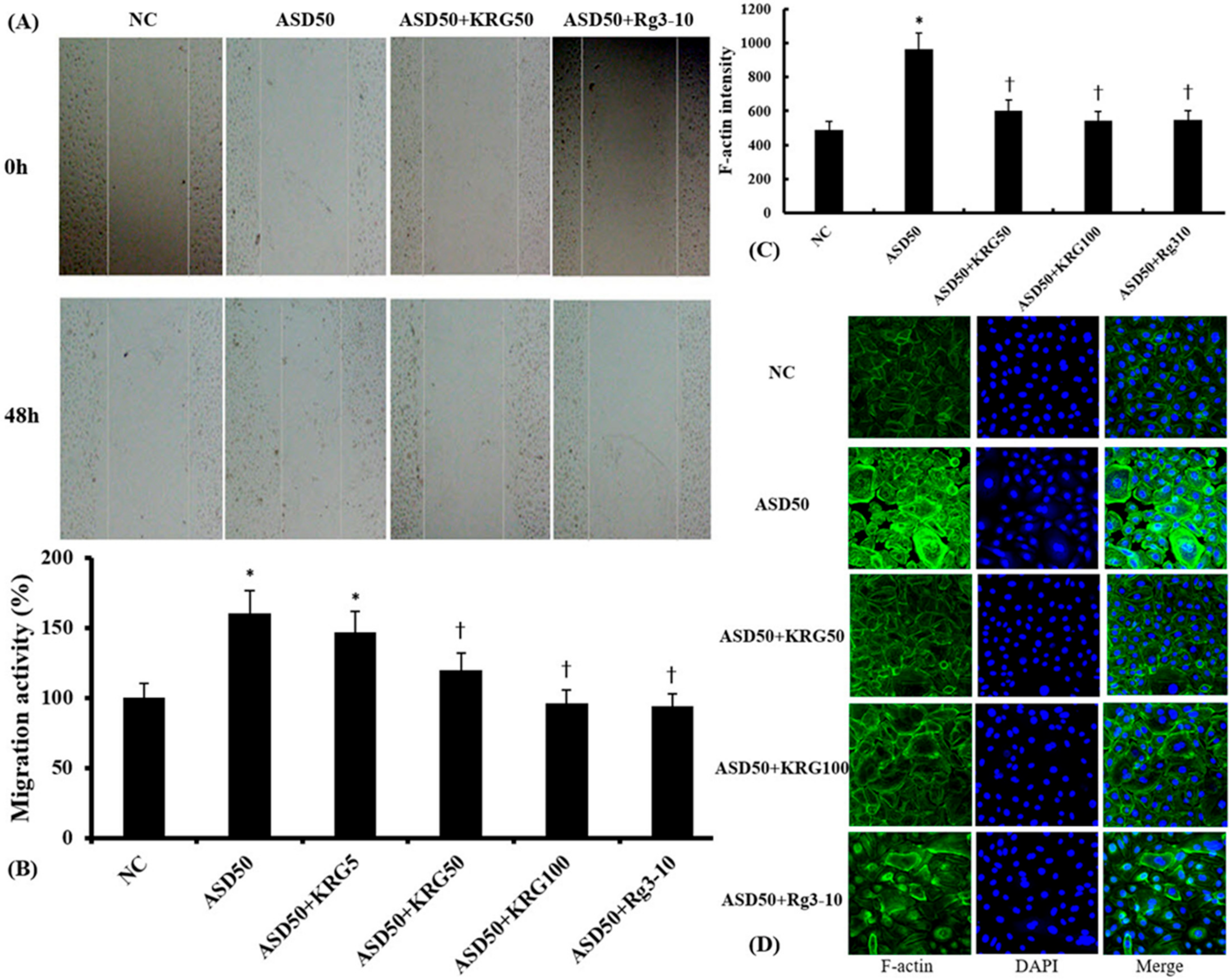
2.5. Effects of KRG and Rg3 on ASD-Induced Migration of Nasal Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Preparation of KRG and ASD
4.2. Primary Nasal Epithelial Cell Culture and Cell Viability Assays
4.3. Analysis of IL-8 and TGF-β1 Production from Nasal Epithelial Cells
4.4. Analysis of the Expression of EMT Marker mRNA and Protein
4.5. Immunofluorescence Study of EMT Marker and F-Actin
4.6. Analysis of Transcription Factors
4.7. Cell Migration Scratch Assays
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Neilson, E.G. Epithelial-mesenchymal transition and its implications for fibrosis. J. Clin. Investig. 2003, 112, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Hupin, C.; Gohy, S.; Bouzin, C.; Lecocq, M.; Polette, M.; Pilette, C. Features of mesenchymal transition in the airway epithelium from chronic rhinosinusitis. Allergy 2014, 69, 1540–1549. [Google Scholar] [CrossRef] [PubMed]
- Konnecke, M.; Burmeister, M.; Pries, R.; Boscke, R.; Bruchhage, K.L.; Ungefroren, H.; Klimek, L.; Wollenberg, B. Epithelial-Mesenchymal Transition in Chronic Rhinosinusitis: Differences Revealed Between Epithelial Cells from Nasal Polyps and Inferior Turbinates. Arch. Immunol. Ther. Exp. 2017, 65, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.W.; Lee, S.A.; Shin, J.M.; Park, I.H.; Lee, H.M. Glucocorticoids ameliorate TGF-beta1-mediated epithelial-to-mesenchymal transition of airway epithelium through MAPK and Snail/Slug signaling pathways. Sci. Rep. 2017, 7, 3486. [Google Scholar] [CrossRef] [Green Version]
- Vetuschi, A.; Pompili, S.; Di Marco, G.P.; Calvaruso, F.; Iacomino, E.; Angelosante, L.; Festuccia, C.; Colapietro, A.; Sferra, R. Can the AGE/RAGE/ERK signalling pathway and the epithelial-to-mesenchymal transition interact in the pathogenesis of chronic rhinosinusitis with nasal polyps? Eur. J. Histochem. 2020, 64, 3079. [Google Scholar] [CrossRef] [Green Version]
- Landis, M.S.; Patrick Pancras, J.; Graney, J.R.; White, E.M.; Edgerton, E.S.; Legge, A.; Percy, K.E. Source apportionment of ambient fine and coarse particulate matter at the Fort McKay community site, in the Athabasca Oil Sands Region, Alberta, Canada. Sci. Total Environ. 2017, 584–585, 105–117. [Google Scholar] [CrossRef]
- Shin, S.H.; Ye, M.K.; Hwang, Y.J.; Kim, S.T. The effect of Asian sand dust-activated respiratory epithelial cells on activation and migration of eosinophils. Inhal. Toxicol. 2013, 25, 633–639. [Google Scholar] [CrossRef]
- Lee, M.; Lim, S.; Kim, Y.S.; Khalmuratova, R.; Shin, S.H.; Kim, I.; Kim, H.J.; Kim, D.Y.; Rhee, C.S.; Park, J.W.; et al. DEP-induced ZEB2 promotes nasal polyp formation via epithelial-to-mesenchymal transition. J. Allergy Clin. Immunol. 2021, 149, 340–357. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cuellar, C.M.; Santibanez-Andrade, M.; Chirino, Y.I.; Quintana-Belmares, R.; Morales-Barcenas, R.; Quezada-Maldonado, E.M.; Sanchez-Perez, Y. Particulate Matter (PM10) Promotes Cell Invasion through Epithelial-Mesenchymal Transition (EMT) by TGF-beta Activation in A549 Lung Cells. Int. J. Mol. Sci. 2021, 22, 2632. [Google Scholar] [CrossRef] [PubMed]
- Kee, J.Y.; Jeon, Y.D.; Kim, D.S.; Han, Y.H.; Park, J.; Youn, D.H.; Kim, S.J.; Ahn, K.S.; Um, J.Y.; Hong, S.H. Korean Red Ginseng improves atopic dermatitis-like skin lesions by suppressing expression of proinflammatory cytokines and chemokines in vivo and in vitro. J. Ginseng Res. 2017, 41, 134–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kee, J.Y.; Han, Y.H.; Mun, J.G.; Park, S.H.; Jeon, H.D.; Hong, S.H. Effect of Korean Red Ginseng extract on colorectal lung metastasis through inhibiting the epithelial-mesenchymal transition via transforming growth factor-beta1/Smad-signaling-mediated Snail/E-cadherin expression. J. Ginseng Res. 2019, 43, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Park, J.H.; Seok, H.; Park, H.J.; Chung, J.H.; Kim, C.J.; Kim, Y.O.; Han, Y.R.; Hong, D.; Kim, Y.S.; et al. The Effects of Korea Red Ginseng on Inflammatory Cytokines and Apoptosis in Rat Model with Chronic Nonbacterial Prostatitis. Biomed. Res. Int. 2019, 2019, 2462561. [Google Scholar] [CrossRef]
- Wei, X.; Chen, Y.; Huang, W. Ginsenoside Rg1 ameliorates liver fibrosis via suppressing epithelial to mesenchymal transition and reactive oxygen species production in vitro and in vivo. Biofactors 2018, 44, 327–335. [Google Scholar] [CrossRef]
- Kim, E.J.; Kwon, K.A.; Lee, Y.E.; Kim, J.H.; Kim, S.H.; Kim, J.H. Korean Red Ginseng extract reduces hypoxia-induced epithelial-mesenchymal transition by repressing NF-kappaB and ERK1/2 pathways in colon cancer. J. Ginseng Res. 2018, 42, 288–297. [Google Scholar] [CrossRef]
- Shimada, A.; Kohara, Y.; Naota, M.; Kobayashi, Y.; Morita, T.; Inoue, K.; Takano, H. Pathological study of chronic pulmonary toxicity induced by intratracheally instilled Asian sand dust (Kosa): Possible association of fibrosis with the development of granulomatous lesions. Folia Histochem. Cytobiol. 2015, 53, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Jacinto, E.; Loewith, R.; Schmidt, A.; Lin, S.; Ruegg, M.A.; Hall, A.; Hall, M.N. Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive. Nat. Cell Biol. 2004, 6, 1122–1128. [Google Scholar] [CrossRef]
- Ichinose, T.; Yoshida, S.; Hiyoshi, K.; Sadakane, K.; Takano, H.; Nishikawa, M.; Mori, I.; Yanagisawa, R.; Kawazato, H.; Yasuda, A.; et al. The effects of microbial materials adhered to Asian sand dust on allergic lung inflammation. Arch. Environ. Contam. Toxicol. 2008, 55, 348–357. [Google Scholar] [CrossRef]
- Hasunuma, H.; Takeuchi, A.; Ono, R.; Amimoto, Y.; Hwang, Y.H.; Uno, I.; Shimizu, A.; Nishiwaki, Y.; Hashizume, M.; Askew, D.J.; et al. Effect of Asian dust on respiratory symptoms among children with and without asthma, and their sensitivity. Sci. Total Environ. 2020, 753, 141585. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Yeon, S.H.; Choi, M.R.; Choi, S.H.; Lee, S.B.; Rha, K.S.; Kim, Y.M. Urban Particulate Matters May Affect Endoplasmic Reticulum Stress and Tight Junction Disruption in Nasal Epithelial Cells. Am. J. Rhinol. Allergy 2021, 35, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Thevenot, P.T.; Saravia, J.; Jin, N.; Giaimo, J.D.; Chustz, R.E.; Mahne, S.; Kelley, M.A.; Hebert, V.Y.; Dellinger, B.; Dugas, T.R.; et al. Radical-containing ultrafine particulate matter initiates epithelial-to-mesenchymal transitions in airway epithelial cells. Am J. Respir. Cell Mol. Biol. 2013, 48, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Min, D.S.; Lee, C.W.; Song, K.H.; Kim, Y.S.; Kim, H.P. Ginsenosides from Korean Red Ginseng ameliorate lung inflammatory responses: Inhibition of the MAPKs/NF-kappaB/c-Fos pathways. J. Ginseng Res. 2018, 42, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Uh, I.; Kim, K.S.; Kim, K.H.; Park, J.; Kim, Y.; Jung, J.H.; Jung, H.J.; Jang, H.J. Anti-Inflammatory Effects of Ginsenoside Rg3 via NF-kappaB Pathway in A549 Cells and Human Asthmatic Lung Tissue. J. Immunol. Res. 2016, 2016, 7521601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, C.; Zhu, G.; Yuan, W.; Xiao, Z.A. TGF-beta1 Induces Epithelial-Mesenchymal Transition of Chronic Sinusitis with Nasal Polyps through MicroRNA-21. Int. Arch. Allergy Immunol. 2019, 179, 304–319. [Google Scholar] [CrossRef] [PubMed]
- Griggs, L.A.; Hassan, N.T.; Malik, R.S.; Griffin, B.P.; Martinez, B.A.; Elmore, L.W.; Lemmon, C.A. Fibronectin fibrils regulate TGF-beta1-induced Epithelial-Mesenchymal Transition. Matrix. Biol. 2017, 60–61, 157–175. [Google Scholar] [CrossRef]
- Shin, H.W.; Cho, K.; Kim, D.W.; Han, D.H.; Khalmuratova, R.; Kim, S.W.; Jeon, S.Y.; Min, Y.G.; Lee, C.H.; Rhee, C.S.; et al. Hypoxia-inducible factor 1 mediates nasal polypogenesis by inducing epithelial-to-mesenchymal transition. Am. J. Respir. Crit. Care Med. 2012, 185, 944–954. [Google Scholar] [CrossRef]
- Qi, L.W.; Wang, C.Z.; Du, G.J.; Zhang, Z.Y.; Calway, T.; Yuan, C.S. Metabolism of ginseng and its interactions with drugs. Curr. Drug Metab. 2011, 12, 818–822. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zou, H.; Gao, Y.; Luo, J.; Xie, X.; Meng, W.; Zhou, H.; Tan, Z. Insights into gastrointestinal microbiota-generated ginsenoside metabolites and their bioactivities. Drug Metab. Rev. 2020, 52, 125–138. [Google Scholar] [CrossRef]
- Cheng, B.; Li, J.; Du, J.; Lv, X.; Weng, L.; Ling, C. Ginsenoside Rb1 inhibits osteoclastogenesis by modulating NF-kappaB and MAPKs pathways. Food Chem. Toxicol. 2012, 50, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M. Anti-inflammatory effects of ginsenosides Rg5, Rz1, and Rk1: Inhibition of TNF-alpha-induced NF-kappaB, COX-2, and iNOS transcriptional expression. Phytother. Res. 2014, 28, 1893–1896. [Google Scholar] [CrossRef]
- Lee, S.M.; Bae, B.S.; Park, H.W.; Ahn, N.G.; Cho, B.G.; Cho, Y.L.; Kwak, Y.S. Characterization of Korean red ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupcsik, L. Estimation of cell number based on metabolic activity: The MTT reduction assay. Methods Mol. Biol. 2011, 740, 13–19. [Google Scholar] [CrossRef]
- Park, I.H.; Kang, J.H.; Shin, J.M.; Lee, H.M. Trichostatin A inhibits epithelial mesenchymal transition induced by TGF-beta1 in airway epithelium. PLoS ONE 2016, 11, e0162058. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IL-8 (pg/mL) | TGF-β1 (pg/mL) | |
|---|---|---|
| Negative control | 180.8 ± 110.5 | 26.0 ± 3.3 |
| ASD50 (μg/mL) | 806.6 ± 386.4 * | 46.6 ± 4.7 * |
| ASD50 + KRG5 (μg/mL) | 816.2 ± 302.8 * | 32.3 ± 5.0 |
| ASD50 + KRG50 (μg/mL) | 842.3 ± 420.7 * | 27.8 ± 2.4 † |
| ASD50 + KRG100 (μg/mL) | 897.1 ± 297.6 * | 26.5 ± 3.4 † |
| ASD50 + Rg3-10 (μg/mL) | 808.3 ± 415.4 * | 27.2 ± 3.7 † |
| Component | Fraction (%) |
|---|---|
| SiO2 | 52.13 |
| Al2O3 | 15.80 |
| Fe2O3 | 5.85 |
| CaO | 4.46 |
| K2O | 2.57 |
| MgO | 2.43 |
| Na2O | 1.59 |
| TiO2 | 0.83 |
| P2O5 | 0.18 |
| MnO | 0.13 |
| ZnO | 0.05 |
| BaO | 0.02 |
| SrO | 0.02 |
| Other elements | 0.22 |
| Loss ignition | 13.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-H.; Ye, M.-K.; Lee, D.-W.; Chae, M.-H.; Hwang, Y.-J. Korean Red Ginseng and Ginsenoside Rg3 Suppress Asian Sand Dust-Induced Epithelial–Mesenchymal Transition in Nasal Epithelial Cells. Molecules 2022, 27, 2642. https://doi.org/10.3390/molecules27092642
Shin S-H, Ye M-K, Lee D-W, Chae M-H, Hwang Y-J. Korean Red Ginseng and Ginsenoside Rg3 Suppress Asian Sand Dust-Induced Epithelial–Mesenchymal Transition in Nasal Epithelial Cells. Molecules. 2022; 27(9):2642. https://doi.org/10.3390/molecules27092642
Chicago/Turabian StyleShin, Seung-Heon, Mi-Kyung Ye, Dong-Won Lee, Mi-Hyun Chae, and You-Jin Hwang. 2022. "Korean Red Ginseng and Ginsenoside Rg3 Suppress Asian Sand Dust-Induced Epithelial–Mesenchymal Transition in Nasal Epithelial Cells" Molecules 27, no. 9: 2642. https://doi.org/10.3390/molecules27092642