
Nutraceutic Potential of Bioactive Compounds of Eugenia dysenterica DC in Metabolic Alterations

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Nutritional Properties of Eugenia dysenterica
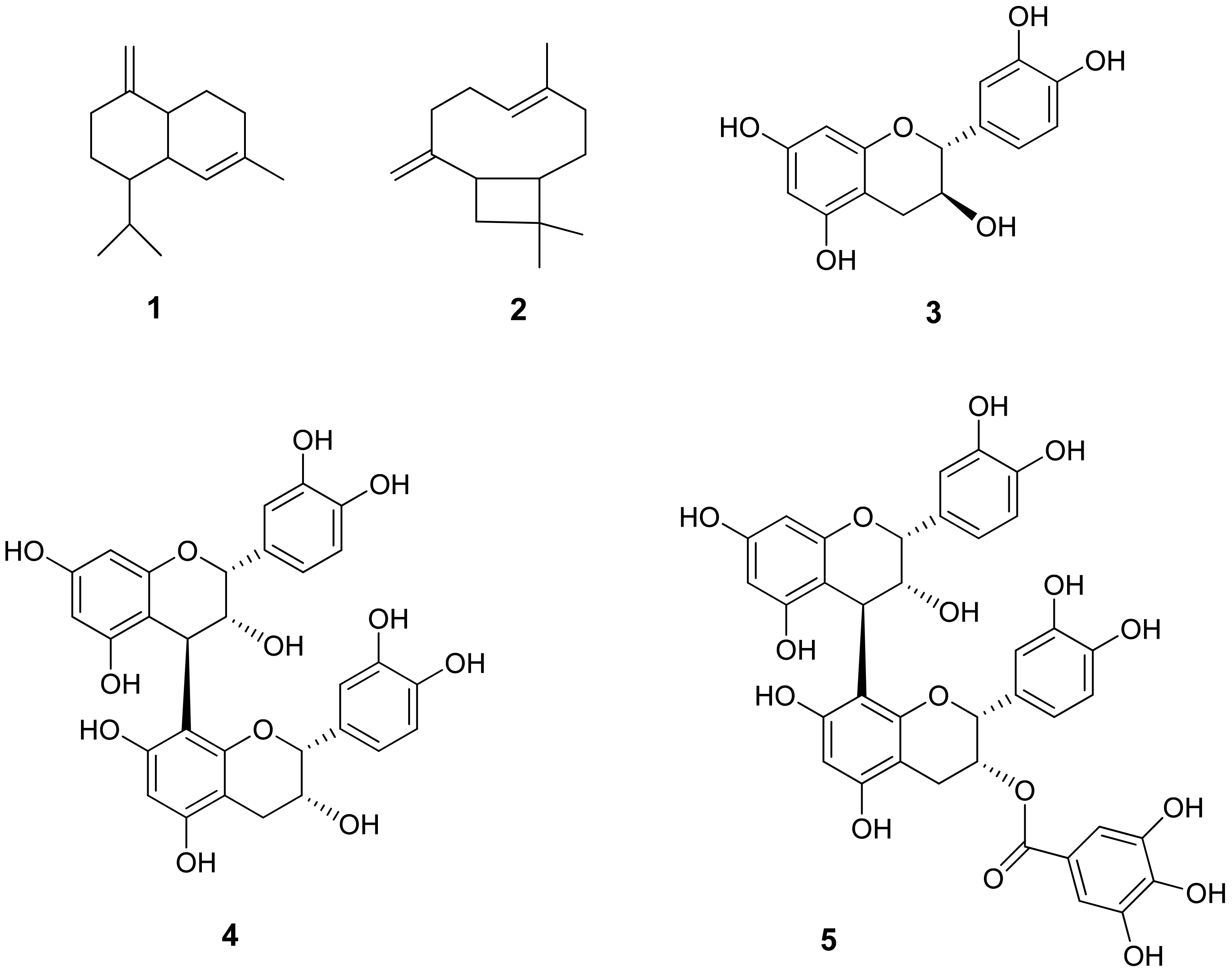
Chemical and Phytochemical Composition
3. Therapeutic Properties of E. dysenterica
3.1. Effects of E. dysenterica on Oxidative Stress
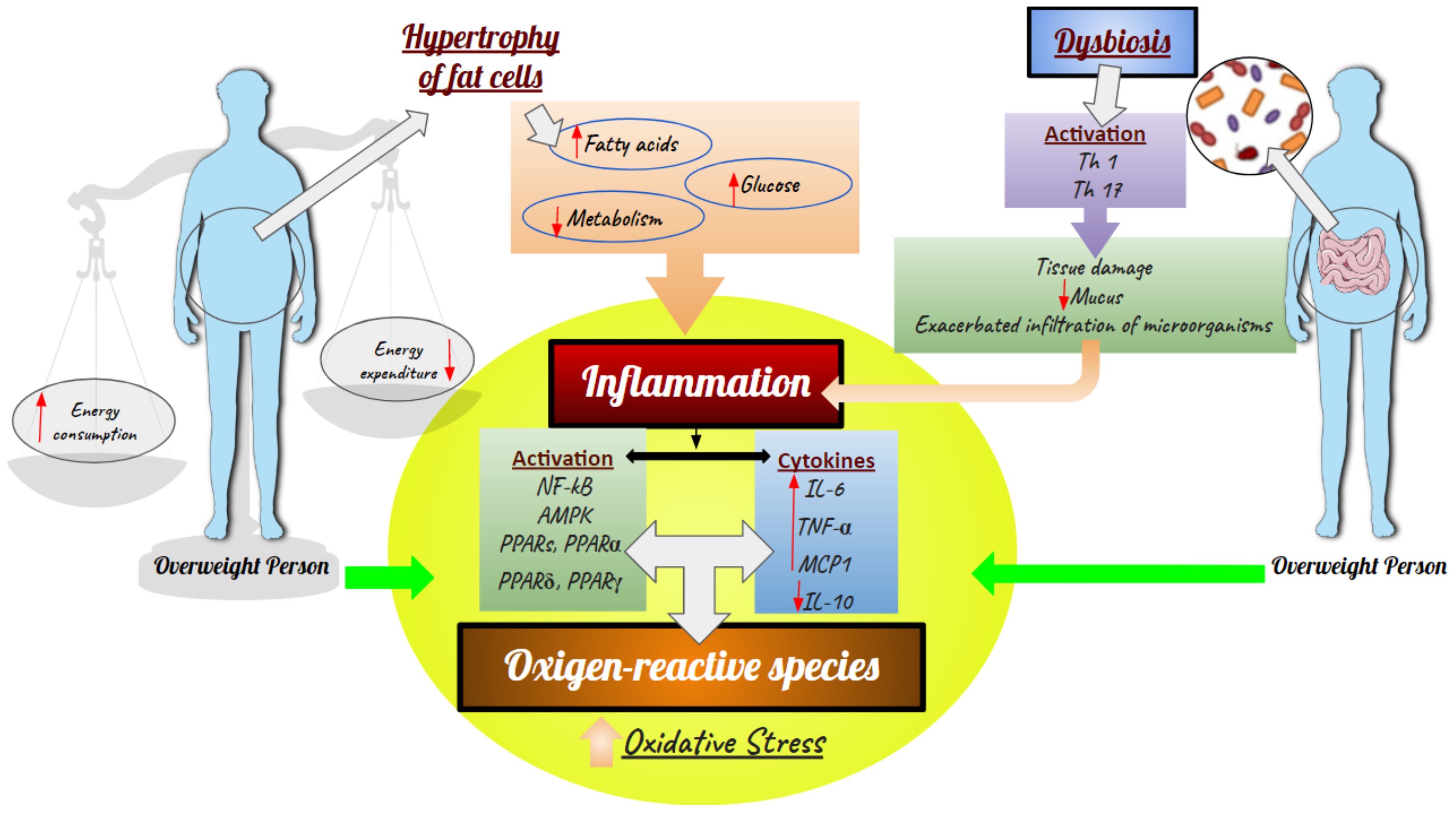
3.2. Effects of E. dysenterica on Diabetes and Inflammation
3.3. Effects of Eugenia dysenterica on Obesity
3.4. Effects of E. dysenterica on Diarrhea, Antimicrobial Activity and Gut Inflammatory Diseases
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Seraglio, S.K.T.; Schulz, M.; Nehring, P.; Della Betta, F.; Valese, A.C.; Daguer, H.; Gonzaga, L.V.; Fett, R.; Costa, A.C.O. Nutritional and Bioactive Potential of Myrtaceae Fruits during Ripening. Food Chem. 2018, 239, 649–656. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.M.; Humberto, M.M.D.S.; Da Silva, J.M.; Rocha, R.D.F.D.A.; Sant’Ana, A.E.G. Estudo Fitoquímico e Avaliação das Atividades Moluscicida e Larvicida dos Extratos da Casca do Caule e Folha de Eugenia Malaccensis L. (Myrtaceae). Rev. Bras. Farm. 2006, 16, 618–624. [Google Scholar] [CrossRef]
- Cole, R.A.; Haber, W.A.; Setzer, W.N. Chemical Composition of Essential Oils of Seven Species of Eugenia from Monteverde, Costa Rica. Biochem. Syst. Ecol. 2007, 35, 877–886. [Google Scholar] [CrossRef]
- Sharma, S.B.; Rajpoot, R.; Nasir, A.; Prabhu, K.M.; Murthy, P.S. Ameliorative Effect of Active Principle Isolated from Seeds of Eugenia Jambolanaon Carbohydrate Metabolism in Experimental Diabetes. Evid.-Based Complement. Altern. Med. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jadeja, R.N.; Thouaojam, M.C.; Sankhari, J.M.; Jain, M.; Devkar, R.V.; Ramachandran, A.V. Standardized Flavonoid-Rich Eugenia Jambolana Seed Extract Retards In Vitro and In Vivo LDL Oxidation and Expression of VCAM-1 and P-Selectin in Atherogenic Rats. Cardiovasc. Toxicol. 2011, 12, 73–82. [Google Scholar] [CrossRef]
- Couto, R.; Valgas, A.; Bara, M.T.; Paula, J.R. Caracterização Físico-Química do pó das Folhas de Eugenia Dysenterica DC. (Myrtaceae). Rev. Eletron. Farm. 2009, 6, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Lima, T.B.; Silva, O.; Silva, L.P.; Rocha, T.L.; Grossi-De-Sa, M.F.; Franco, O.L.; Leonardecz, E. In Vivo Effects of Cagaita (Eugenia Dysenterica, DC.) Leaf Extracts on Diarrhea Treatment. Evid.-Based Complement. Altern. Med. 2010, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lima, T.B.; Silva, O.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Scalabrin, F.B.; Rocha, T.L.; Castro, C.F.S. Identification of E. dysenterica laxative peptide: A novel strategy in the treatment of chronic constipation and irritable bowel syndrome. Peptides 2010, 31, 1426–1433. [Google Scholar] [CrossRef]
- Galheigo, M.R.U.; Prado, L.C.D.S.; Mundin, A.M.M.; Gomes, D.O.; Chang, R.; Lima, A.M.C.; Canabrava, H.A.N.; Bispo-Da-Silva, L.B. Antidiarrhoeic Effect of Eugenia Dysenterica DC (Myrtaceae) Leaf Essential Oil. Nat. Prod. Res. 2015, 30, 1182–1185. [Google Scholar] [CrossRef]
- Souza, P.M.; Elias, S.; Simeoni, L.A.; De Paula, J.E.; Gomes, S.M.; Guerra, E.N.S.; Fonseca, Y.M.; Silva, E.C.; Silveira, D.; Magalhães, P.O. Plants from Brazilian Cerrado with Potent Tyrosinase Inhibitory Activity. PLoS ONE 2012, 7, e48589. [Google Scholar] [CrossRef] [Green Version]
- Costa, T.R.; Fernandes, O.F.; Santos, S.; Oliveira, C.M.; Lião, L.M.; Ferri, P.H.; de Paula, J.R.; Ferreira, H.D.; Sales, B.H.; Silva, M.D.R.R. Antifungal Activity of Volatile Constituents of Eugenia Dysenterica Leaf Oil. J. Ethnopharmacol. 2000, 72, 111–117. [Google Scholar] [CrossRef]
- Genovese, M.I.; Pinto, M.D.S.; Gonçalves, A.E.D.S.S.; Lajolo, F.M. Bioactive Compounds and Antioxidant Capacity of Exotic Fruits and Commercial Frozen Pulps from Brazil. Food Sci. Technol. Int. 2008, 14, 207–214. [Google Scholar] [CrossRef]
- Cecílio, A.B.; de Faria, D.B.; Oliveira, P.D.C.; Caldas, S.; de Oliveira, D.A.; Sobral, M.E.G.; Duarte, M.G.R.; Moreira, C.P.D.S.; Silva, C.G.; de Almeida, V.L. Screening of Brazilian Medicinal Plants for Antiviral Activity against Rotavirus. J. Ethnopharmacol. 2012, 141, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Haslam, E. Natural Polyphenols (Vegetable Tannins) as Drugs: Possible Modes of Action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef]
- De Bruyne, T.; Pieters, L.; Deelstra, H.; Vlietinck, A. Condensed Vegetable Tannins: Biodiversity in Structure and Biological Activities. Biochem. Syst. Ecol. 1999, 27, 445–459. [Google Scholar] [CrossRef]
- Dufresne, C.J.; Farnworth, E.R. A Review of Latest Research Findings on the Health Promotion Properties of Tea. J. Nutr. Biochem. 2001, 12, 404–421. [Google Scholar] [CrossRef]
- Santana, L.F.; Santo, B.L.S.E.; Sasso, S.; Aquino, D.F.S.; Cardoso, C.A.L.; Pott, A.; Soares, F.L.P.; Freitas, K.C. Efeitos do Extrato Etanólico das Folhas de Cagaiteira (Eugenia dysenterica DC.) em Camundongos Diabéticos Induzidos por Estreptozotocina. Int. J. Dev. Res. 2018, 8, 1355–1362. [Google Scholar]
- Nascimento, A.L.A.A.; Brandi, I.V.; Durães, C.A.F.; Lima, J.P.; Soares, S.B.; Mesquita, B.M.A.C. Chemical characterization and antioxidant potential of native fruits of the Cerrado of northern Minas Gerais. Braz. J. Food Technol. 2020, 23, e2019296. [Google Scholar] [CrossRef]
- Jorge, N.; Moreno, D.M.; Bertanha, B.J. Eugenia Dysenterica DC: Actividad Antioxidante, Perfil de Ácidos Grasos y Determinación de Tocoferoles. Rev. Chil. Nutr. 2010, 37, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Martinotto, C.; Paiva, R.; Soares, F.P.; Santos, B.R.; Nogueira, R.C. Cagaiteira (Eugenia Dysenterica DC.). Lavras-MG: Universidade Federal de Lavras. Bol. Téc. 2008, 78, 1–21. [Google Scholar]
- Brito, M.A.; Pereira, E.B.C.; Pereira, A.V.; Ribeiro, J.F. Cagaita: Biologia e Manejo. Empresa Brasileira de Pesquisa Agropecuária CPAC; Planaltina, D.F., Ed.; EMBRAPA Cerrados: Brasilia, Brazil, 2003. [Google Scholar]
- Silvia, M.R.; Lacerda, D.B.C.L.; Santos, G.G.; Martins, D.M.O. Caracterização Química de Frutos Nativos do Cerrado. Cien. Rural 2008, 38, 1790–1793. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, E.M.G.; De Carvalho, L.M.J.; Ortiz, G.M.D.; Cardoso, F.D.S.N.; Soares, D.; De Carvalho, J.L.V.; Barros, P.; Machado, N. An Overview on Cagaita (Eugenia Dysenterica DC) Macro and Micro Components and a Technological Approach. Food Ind. 2013, 9, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Santana, L.F.; Espirito Santo, B.L.S.; do Sasso, S.; Aquino, D.F.S.; Cardoso, C.A.L.; Pott, A.; Soares, F.L.P.S.; Freitas, K.C.F. Ethanolic Extract Leaves of Eugenia dysenterica DC Improves the Lipid Profile in Mice with Diabetes Induced by Streptozotocin. Int. J. Dev. Res. 2018, 8, 1355–1362. Available online: https://www.journalijdr.com/ethanolic-extract-leaves-eugenia-dysenterica-dc-improves-lipid-profile-mice-diabetes-induced (accessed on 23 January 2022).
- Bailão, E.F.L.C.; Devilla, I.A.; Da Conceição, E.C.; Borges, L.L. Bioactive Compounds Found in Brazilian Cerrado Fruits. Int. J. Mol. Sci. 2015, 16, 23760–23783. [Google Scholar] [CrossRef]
- Donado-Pestana, C.M.; Belchior, T.; Genovese, M.I. Phenolic Compounds from Cagaita (Eugenia Dysenterica DC.) Fruit Prevent Body Weight and Fat Mass Gain Induced by a High-Fat, High-Sucrose Diet. Food Res. Int. 2015, 77, 177–185. [Google Scholar] [CrossRef]
- Daza, L.D.; Fujita, A.; Granato, D.; Fávaro-Trindade, C.S.; Genovese, M.I. Functional Properties of Encapsulated Cagaita (Eugenia Dysenterica DC.) Fruit Extract. Food Biosci. 2017, 18, 15–21. [Google Scholar] [CrossRef]
- Hwang, K.-A.; Hwang, Y.-J.; Song, J. Antioxidant Activities and Oxidative Stress Inhibitory Effects of Ethanol Extracts from Cornus Officinalis on Raw 264.7 Cells. BMC Complement. Altern. Med. 2016, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.-L. Novel Insights of Dietary Polyphenols and Obesity. J. Nutr. Biochem. 2013, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- De Souza, A.M.; De Oliveira, C.F.; Oliveira, V.; Betim, F.C.M.; Miguel, O.G.; Miguel, M.D. Traditional Uses, Phytochemistry, and Antimicrobial Activities of Eugenia Species—A Review. Planta Med. 2018, 84, 1232–1248. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Vitamin E As a Potential Interventional Treatment for Metabolic Syndrome: Evidence from Animal and Human Studies. Front. Pharmacol. 2017, 8, 444. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Vega, J.A.; Arteaga-Badillo, D.A.; Sánchez-Gutiérrez, M.; Morales-González, J.A.; Vargas-Mendoza, N.; Gómez-Aldapa, C.A.; Castro-Rosas, J.; Delgado-Olivares, L.; Madrigal-Bujaidar, E.; Madrigal-Santillán, E. Organic Acids from Roselle (Hibiscus sabdariffa L.)—A Brief Review of Its Pharmacological Effects. Biomedicines 2020, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Olefsky, J.M. Inflammation and Lipid Signaling in the Etiology of Insulin Resistance. Cell Metab. 2012, 15, 635–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geloneze, B.; Tambascia, M.A. Laboratorial Evaluation and Diagnosis of Insulin Resistance. Arq. Bras. Endocrinol. Metab. 2006, 50, 208–215. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Cardiovascular Disease and Risk Management. Diabetes Care 2014, 38, S49–S57. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, M. Biochemistry and Molecular Cell Biology of Diabetic Complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative Stress and Diabetic Complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.S.; Harris, A.K.; Rychly, D.J.; Ergul, A. Oxidative Stress and the Use of Antioxidants in Diabetes: Linking Basic Science to Clinical Practice. Cardiovasc. Diabetol. 2005, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.-H.; Liu, J.-J.; Yeh, C.-L.; Chiu, W.-C.; Yeh, S.-L. Effects of Glutamine Supplementation on Oxidative Stress-Related Gene Expression and Antioxidant Properties in Rats with Streptozotocin-Induced Type 2 Diabetes. Br. J. Nutr. 2011, 107, 1112–1118. [Google Scholar] [CrossRef] [Green Version]
- Taniyama, Y.; Griendling, K. Reactive Oxygen Species in the Vasculature. Hypertension 2003, 42, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Palhares, D. Caracterização Farmacognóstica das Folhas de Eugenia Dysenterica DC (Myrtaceae Jussieu). Rev. Lecta 2003, 21, 29–36. [Google Scholar]
- Araujo, N.M.P.; Arruda, H.S.; Farias, D.D.P.; Molina, G.; Pereira, G.A.; Pastore, G.M. Plants from the Genus Eugenia as Promising Therapeutic Agents for the Management of Diabetes Mellitus: A Review. Food Res. Int. 2021, 142, 110182. [Google Scholar] [CrossRef]
- Gonçalves, A.E.D.S.S.; Lajolo, F.M.; Genovese, M.I. Chemical Composition and Antioxidant/Antidiabetic Potential of Brazilian Native Fruits and Commercial Frozen Pulps. J. Agric. Food Chem. 2010, 58, 4666–4674. [Google Scholar] [CrossRef]
- Sano, S.M.; da Fonseca, C.E.L.; Ribeiro, J.F.; Oga, F.M.; Luiz, A.J.B. Folhação, Floração, Frutificação e Crescimento Inicial da Cagaiteira em Planatina; DF. Pesq. Agropec. Bras. 1995, 30, 5–14. [Google Scholar]
- Donado-Pestana, C.M.; dos Santos-Donado, P.R.; Daza, L.D.; Belchior, T.; Festuccia, W.T.; Genovese, M.I. Cagaita Fruit (Eugenia Dysenterica DC.) and Obesity: Role of Polyphenols on Already Established Obesity. Food Res. Int. 2018, 103, 40–47. [Google Scholar] [CrossRef]
- Sousa, F.J.S.A.; Do Espirito Santo, B.L.S.; Sasso, S.; Santana, L.F.; Guércio, A.C.; Cardoso, C.A.L.; De Cássia Freitas, K. Efetividade do Extrato da Folha da Cagaiteira (Eugenia Dysenterica DC.) no Perfil Metabólico em Animais Alimentados com Dieta Hipercalórica. PECIBES 2016, 2, 122–123. [Google Scholar]
- Da Silva, S.M.M.; Costa, C.R.R.; Gelfuso, G.M.; Guerra, E.N.S.; Nóbrega, Y.K.D.M.; Gomes, S.M.; Pic-Taylor, A.; Fonseca-Bazzo, Y.M.; Silveira, D.; Magalhães, P.D.O. Wound Healing Effect of Essential Oil Extracted from Eugenia Dysenterica DC (Myrtaceae) Leaves. Molecules 2018, 24, 2. [Google Scholar] [CrossRef] [Green Version]
- Petsiou, E.I.; Mitrou, P.I.; Raptis, S.A.; Dimitriadis, G.D. Effect and Mechanisms of Action of Vinegar on Glucose Metabolism, Lipid Profile, and Body Weight. Nutr. Rev. 2014, 72, 651–661. [Google Scholar] [CrossRef]
- Liu, Q.; Tang, G.-Y.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Antioxidant Activities, Phenolic Profiles, and Organic Acid Contents of Fruit Vinegars. Antioxidants 2019, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Costa, C.R.R.; Amorim, B.R.; Da Silva, S.M.M.; Acevedo, A.C.; Magalhães, P.D.O.; Guerra, E.N.S. In Vitro Evaluation of Eugenia Dysenterica in Primary Culture of Human Gingival Fibroblast Cells. Braz. Oral Res. 2019, 33, e035. [Google Scholar] [CrossRef]
- Lee, W.-L.; Huang, J.-Y.; Shyur, L.-F. Phytoagents for Cancer Management: Regulation of Nucleic Acid Oxidation, ROS, and Related Mechanisms. Oxid. Med. Cell. Longev. 2013, 2013, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Chistiane, M.F.; Alisson, D.R.B.; Cassio, H.S.D.M.; Rivelilson, M.F.; Jose, E.D.N.F.; Emmanoel, V.C.; Khaled, N.Z.R.; Joaquim, S.D.C.J. Antioxidant and Anticholinesterase Activities of the Essential Oil of Eugenia Dysenterica DC. Afr. J. Pharm. Pharmacol. 2017, 11, 241–249. [Google Scholar] [CrossRef]
- Sasso, S.; Souza, P.C.S.E.; Santana, L.F.; Cardoso, C.A.L.; Alves, F.M.; Portugal, L.C.; De Faria, B.B.; Da Silva, A.F.; Motta-Castro, A.R.C.; Soares, L.S.; et al. Use of an Extract of Annona Muricata Linn to Prevent High-Fat Diet Induced Metabolic Disorders in C57BL/6 Mice. Nutrients 2019, 11, 1509. [Google Scholar] [CrossRef] [Green Version]
- Her, T.; Lagakos, W.S.; Brown, M.R.; LeBrasseur, N.K.; Rakshit, K.; Matveyenko, A. Dietary Carbohydrates Modulate Metabolic and Beta Cell Adaptation to High Fat Diet Indu Obesity. Am. J. Physiol.-Endocrinol. Metabo. 2020, 318, E856–E865. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.C.; Ceschini, F.L.; Ramallo, B.T. Resistência à Insulina Associada à Obesidade: Efeitos Anti-Inflamatórios do Exercício Físico. Rev. Bras. Ciên. Movim. 2014, 22, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte Dysfunctions Linking Obesity to Insulin Resistance and Type 2 Diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar Singh, A.; Cabral, C.; Kumar, R.; Ganguly, R.; Kumar Rana, H.; Gupta, A.; Rosaria Lauro, M.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial Effects of Dietary Polyphenols on Gut Microbiota and Strategies to Improve Delivery Efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Serrano, A.; Bladé, C.; Suárez, M.; Arola-Arnal, A. Grape Seed Proanthocyanidins Improve White Adipose Tissue Expansion during Diet-Induced Obesity Development in Rats. Int. J. Mol. Sci. 2018, 19, 2632. [Google Scholar] [CrossRef] [Green Version]
- Patil, M.P.; Patil, R.H. Data on the Inhibitory Effect of Endophytic Fungi of Traditional Medicinal Plants against Pancreatic Lipase (PL). Data Brief 2019, 27, 104797. [Google Scholar] [CrossRef]
- Anhê, F.F.; Desjardins, Y.; Pilon, G.; Dudonné, S.; Genovese, M.I.; Lajolo, F.M.; Marette, A. Polyphenols and Type 2 Diabetes: A Prospective Review. PharmaNutrition 2013, 1, 105–114. [Google Scholar] [CrossRef]
- Balisteiro, D.M.; de Araujo, R.L.; Giacaglia, L.R.; Genovese, M.I. Effect of Clarified Brazilian Native Fruit Juices on Postprandial Glycemia in Healthy Subjects. Food Res. Int. 2017, 100, 196–203. [Google Scholar] [CrossRef]
- Brown, J.C.; Gerhardt, T.E.; Kwon, E. Risk Factors for Coronary Artery Disease. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554410/ (accessed on 5 March 2020).
- Sekhon-Loodu, S.; Rupasinghe, H.P.V. Evaluation of Antioxidant, Antidiabetic and Antiobesity Potential of Selected Traditional Medicinal Plants. Front. Nutr. 2019, 6, 1–34. [Google Scholar] [CrossRef]
- Mounien, L.; Tourniaire, F.; Landrier, J.-F. Anti-Obesity Effect of Carotenoids: Direct Impact on Adipose Tissue and Adipose Tissue-Driven Indirect Effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Nagashimada, M.; Zhan, L.; Nagata, N.; Kobori, M.; Sugiura, M.; Ogawa, K.; Kaneko, S.; Ota, T. Prevention and Reversal of Lipotoxicity-Induced Hepatic Insulin Resistance and Steatohepatitis in Mice by an Antioxidant Carotenoid, β-Cryptoxanthin. Endocrinology 2015, 156, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Alcalá, M.; Sánchez-Vera, I.; Sevillano, J.; Herrero, L.; Serra, D.; Ramos, M.P.; Viana, M. Vitamin E Reduces Adipose Tissue Fibrosis, Inflammation, and Oxidative Stress and Improves Metabolic Profile in Obesity. Obesity 2015, 23, 1598–1606. [Google Scholar] [CrossRef]
- Ellulu, M.S. Obesity, Cardiovascular Disease, and Role of Vitamin C on Inflammation: A Review of Facts and Underlying Mechanisms. Inflammopharmacology 2017, 25, 313–328. [Google Scholar] [CrossRef]
- Eseberri, I.; Miranda, J.; Lasa, A.; Mosqueda-Solís, A.; González-Manzano, S.; Santos-Buelga, C.; Portillo, M.P. Effects of Quercetin Metabolites on Triglyceride Metabolism of 3T3-L1 Preadipocytes and Mature Adipocytes. Int. J. Mol. Sci. 2019, 20, 264. [Google Scholar] [CrossRef] [Green Version]
- Torres-Villarreal, D.; Camacho-Morales, A.; Castro, H.; Ortiz-Lopez, R.; De La Garza, A.L. Anti-Obesity Effects of Kaempferol by Inhibiting Adipogenesis and Increasing Lipolysis in 3T3-L1 Cells. J. Physiol. Biochem. 2018, 75, 83–88. [Google Scholar] [CrossRef]
- Wang, L.; Wei, Y.; Ning, C.; Zhang, M.; Fan, P.; Lei, D.; Du, J.; Gale, M.; Ma, Y.; Yang, Y. Ellagic Acid Promotes Browning of White Adipose Tissues in High-Fat Diet-Induced Obesity in Rats through Suppressing White Adipocyte Maintaining Genes. Endocr. J. 2019, 66, 923–936. [Google Scholar] [CrossRef] [Green Version]
- González-Sarrías, A.; Villalba, R.G.; Vaquero, M.R.; Alasalvar, C.; Örem, A.; Zafrilla, P.; Tomas-Barberan, F.A.; Selma, M.V.; Espín, J.C. Clustering According to Urolithin Metabotype Explains the Interindividual Variability in the Improvement of Cardiovascular Risk Biomarkers in Overweight-Obese Individuals Consuming Pomegranate: A Randomized Clinical Trial. Mol. Nutr. Food Res. 2017, 61, 1600830. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; González-Sarrías, A.; Salas-Salvadó, J.; Andres-Lacueva, C.; Alasalvar, C.; Örem, A.; Tomás-Barberán, F.A.; Espín, J.C. The Gut Microbiota Metabolism of Pomegranate or Walnut Ellagitannins Yields Two Urolithin-Metabotypes that Correlate with Cardiometabolic Risk Biomarkers: Comparison between Normoweight, Overweight-Obesity and Metabolic Syndrome. Clin. Nutr. 2017, 37, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Yang, J.; Lee, R.-P.; Huang, J.; Hsu, M.; Thames, G.; Gilbuena, I.; Long, J.; Xu, Y.; Park, E.H.; et al. Pomegranate Juice and Extract Consumption Increases the Resistance to UVB-Induced Erythema and Changes the Skin Microbiome in Healthy Women: A Randomized Controlled Trial. Sci. Rep. 2019, 9, 14528. [Google Scholar] [CrossRef] [PubMed]
- Montalto, M.; D’Onofrio, F.; Gallo, A.; Cazzato, A.; Gasbarrini, G. Intestinal Microbiota and Its Functions. Dig. Liver Dis. Suppl. 2009, 3, 30–34. [Google Scholar] [CrossRef]
- Caruso, R.; Lo, B.C.; Núñez, G. Host–Microbiota Interactions in Inflammatory Bowel Disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef]
- Fidelis-De-Oliveira, P.; Aparecida-Castro, S.; Silva, D.B.; Morais, I.B.D.M.; De Miranda, V.H.M.; De Gobbi, J.; Canabrava, H.A.N.; Bispo-Da-Silva, L.B. Hypotensive Effect of Eugenia Dysenterica Leaf Extract Is Primarily Related to Its Vascular Action: The Possible Underlying Mechanisms. J. Ethnopharmacol. 2019, 251, 112520. [Google Scholar] [CrossRef]
- Farré, R.; Fiorani, M.; Rahiman, S.A.; Matteoli, G. Intestinal Permeability, Inflammation and the Role of Nutrients. Nutrients 2020, 12, 1185. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.-U.; Chen, G.Y.; Núñez, G. Role of the Gut Microbiota in Immunity and Inflammatory Disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Yarnell, E.; Abascal, K. Herbs for Gastroesophageal Reflux Disease. Altern. Complement. Ther. 2010, 16, 344–346. [Google Scholar] [CrossRef]
- Neurath, M.F. Cytokines in Inflammatory Bowel Disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef]
- Prado, L.C.D.S.; da Silva, D.; de Oliveira-Silva, G.L.; Hiraki, K.R.N.; Canabrava, H.A.N.; Bispo-Da-Silva, L.B. The Gastroprotective Effects of Eugenia Dysenterica (Myrtaceae) Leaf Extract: The Possible Role of Condensed Tannins. Biol. Pharm. Bull. 2014, 37, 722–730. [Google Scholar] [CrossRef] [Green Version]
- de Araújo, F.F.; Neri-Numa, I.A.; Farias, D.; da Cunha, G.R.M.C.; Pastore, G.M. Wild Brazilian Species of Eugenia Genera (Myrtaceae) as an Innovation Hotspot for Food and Pharmacological Purposes. Food Res. Int. 2019, 121, 57–72. [Google Scholar] [CrossRef]
- Vieira, P.M.; Veronezi, E.; Silva, C.R.; Chen-Chen, L. Detection of Genotoxic, Cytotoxic, and Protective Activities of Eugenia dysenterica DC. (Myrtaceae) in Mice. J. Med. Food 2012, 15, 563–567. [Google Scholar] [CrossRef]
- Corrêa, T.; Rogero, M.M. Polyphenols Regulating MicroRNAs and Inflammation Biomarkers in Obesity. Nutrition 2018, 59, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.-J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.; Piao, M.; Song, Y. Dietary Quercetin Increases Colonic Microbial Diversity and Attenuates Colitis Severity in Citrobacter Rodentium-Infected Mice. Front. Microbiol. 2019, 10, 1092. [Google Scholar] [CrossRef]
- Chen, J.; Xuan, Y.-H.; Luo, M.-X.; Ni, X.-G.; Ling, L.-Q.; Hu, S.-J.; Xu, J.-Y.; Jiang, L.-Y.; Si, W.-Z.; Xu, L.; et al. Kaempferol Alleviates Acute Alcoholic Liver Injury in Mice by Regulating Intestinal Tight Junction Proteins and Butyrate Receptors and Transporters. Toxicology 2019, 429, 152338. [Google Scholar] [CrossRef]
- Kang, I.; Buckner, T.; Shay, N.F.; Gu, L.; Chung, S. Improvements in Metabolic Health with Consumption of Ellagic Acid and Subsequent Conversion into Urolithins: Evidence and Mechanisms. Adv. Nutr. Int. Rev. J. 2016, 7, 961–972. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Components | Pulp | Seeds |
|---|---|---|
| Proteins (g) | 1.140.63 | 0.59 |
| Carbohydrates (g) | 6.33 | 14.85 |
| Lipids (g) | 0.05 | 0.26 |
| Fibers (g) | 1.05 | |
| Total energetic value (kcal) | 30.33 | 64.10 |
| Eugenia dysenterica DC. | |||
|---|---|---|---|
| Phytochemical Compounds | Pulp | Seeds | Leaves |
| Ascorbic acid Folates Carotenoid β-carotene Lycopene Tannins α-, β-, γ- e δ-Tocopherol | 24.53 mg 25.74 μg 0.77 mg − − − − | − − − + + − − | − + − + + + + |
| Minerals | Pulp | Seeds | Leaves |
| Calcium Iron Zinc Phosphorous Potassium Magnesium Sulphur Boron Copper Manganese | 8.0 mg 0.02 mg − − − − − − − − | − − − − − − − − − − | 0.84% 145 ppm 21.40 ppm 0.14% 1.20% 0.28% 0.06% 32.50 ppm 15 ppm 163 ppm |
| Eugenia dysenterica DC. | ||
|---|---|---|
| Bioactive Compounds | Pulp | Leaves |
| Quercetin Kaempferol Ellagic acid Ellagitannis Proanthocianidins Phenolic acids Flavonoids Anthocyanins Organic acids Polyphenols γ-Cadinene δ-Cadinene | + + + + + + + + + + − − | − − − − + − − − − + + + |
| Plant Part | Host | Treatments | Main Effects | Reference |
|---|---|---|---|---|
| Leaf aqueous extract | Gastric lesion model induced by acidified ethanol | 6 groups of rats that had fasted for 24 h were given oral saline solution, carbenoxolone or E. dysentherica leaf aqueous extract (100, 300, 550, 1000 mg/kg); 50 min after treatments, all received oral HCl 0.3 M/ethanol at 60%. Antioxidant activity of the extract determined in vitro. | ↑ protection of gastric mucosaagainst lesions induced by ethanol/HCl-c, ↓ production of HCl-c, ↓ free radicals, ↑ protective liningagainstharmful agents (aqueous extract 550 and 1000 mg/kg). | [82] |
| Pulp in natura and peptide | Male rats | Positive and negative control groups received oral castor oil and water at the dose of 10 mL/kg. Control groups received pulp at 10 mL/kg and peptideat 60 mg/kg. | ↑ intestinal transit by 14.8% (pulp in natura) and 19% (peptide) without causing diarrhea. | [8] |
| Aqueous infusion and ethanolic extract | Animal model of diarrhea induced by castor oil | Forty rats were separated into five groups of eight animals. The positive control group received loperamide orally at a dose of 2 mg/kg, and the negative group received 1 mL of water; test groups received aqueous extract at doses of 800 mg/kg or ethanolic extract at doses of 400 mg/kg. | ↓ intestinal transit by 24%, ↑ serum chloride levels, ↓ serum phosphorus and magnesium levels, ↑ alanine aminotransferase levels (ethanolic extract). | [7] |
| Pulp Extract | Male mice C57BL/6J, obesity model induced by high fat diet and sucrose | Group 1: A chow-fed group + water administrated by gavage Group 2: high-fat high-sucrose diet + water administered by gavage Group 3: high-fat high-sucrose diet + phenolic-rich extract from E. dysentherica by gavage (7 mg GAE/kg body weight) Group 4: high-fat high-sucrose diet + phenolic-rich extract from E. dysentherica by gavage (14 mg GAE/kg body weight). | ↓ body weight gain, ↓ increase in retroperitoneal, epididymal and brown adipocyte deposits, ↓ fecal lipids, ↓ levels of plasma and hepatic triacylglycerols, ↑ plasma (pulp extract). | [26,84] |
| Pulp extract | Animal model for obesity study | After 6 weeks of feeding with high-fat high-sucrose (HFS) diet or chow diets, mice in the HFS dietary group were randomlydivided into three groups: Group 1: HFS + water given by gavage Group 2: HFS + phenolic-rich extract from E. dysenthericagiven by gavage (7 mg GAE/kg body weight) Group 3: HFS + phenolic-rich extract from E. dysentherica given by gavage (14 mg GAE/kg body weight) Group 4: A chow-fed group + water administered by gavage. | ↓ adipocyte size, ↓ hyperglycemia and dyslipidemia, ↓ serum levels of NEFA and LDL-cholesterol, ↓ aminotransferase activity, ↓ fasting hyperglycemia, ↓glucose intolerance, ↓ pyruvate carboxylase mRNA (pulp extract). | [57,88] |
| Juices from Brazilian native fruit | Healthy volunteer individuals | Each meal consisted of ca. 25 g of available carbohydrate as white bread (corresponding to one unit ca. 50 g) and 300 mL of water (control), or clarified fruit juices, given after 10–12 h of fasting. All underwent tests with water (control) and clarified juices at 7-day intervals in between. | ↓ postprandial glucose, ↓ oxidative stress (juice of pulp) | [62] |
| Aqueous leaf extract | Animal model for cardiovascular study | Procedures in rats under anesthesia. Direct measure of arterial pressure and intravenousadministration of extract or drugs through adequate inserted polyetene catheters in the femoral artery and vein, respectively. | ↓ mean arterial pressure, type-L calcium channel blockade, as well as myoendothelial gap-junction signaling (aqueous leaf extract) | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, L.F.; Sasso, S.; Aquino, D.F.S.; de Cássia Freitas, K.; de Cássia Avellaneda Guimarães, R.; Pott, A.; do Nascimento, V.A.; Bogo, D.; de Oliveira Figueiredo, P.; Hiane, P.A. Nutraceutic Potential of Bioactive Compounds of Eugenia dysenterica DC in Metabolic Alterations. Molecules 2022, 27, 2477. https://doi.org/10.3390/molecules27082477
Santana LF, Sasso S, Aquino DFS, de Cássia Freitas K, de Cássia Avellaneda Guimarães R, Pott A, do Nascimento VA, Bogo D, de Oliveira Figueiredo P, Hiane PA. Nutraceutic Potential of Bioactive Compounds of Eugenia dysenterica DC in Metabolic Alterations. Molecules. 2022; 27(8):2477. https://doi.org/10.3390/molecules27082477
Chicago/Turabian StyleSantana, Lidiani Figueiredo, Sandramara Sasso, Diana Figueiredo Santana Aquino, Karine de Cássia Freitas, Rita de Cássia Avellaneda Guimarães, Arnildo Pott, Valter Aragão do Nascimento, Danielle Bogo, Patrícia de Oliveira Figueiredo, and Priscila Aiko Hiane. 2022. "Nutraceutic Potential of Bioactive Compounds of Eugenia dysenterica DC in Metabolic Alterations" Molecules 27, no. 8: 2477. https://doi.org/10.3390/molecules27082477