
NO in Viral Infections: Role and Development of Antiviral Therapies



Abstract
:1. Introduction
2. Biology of NO
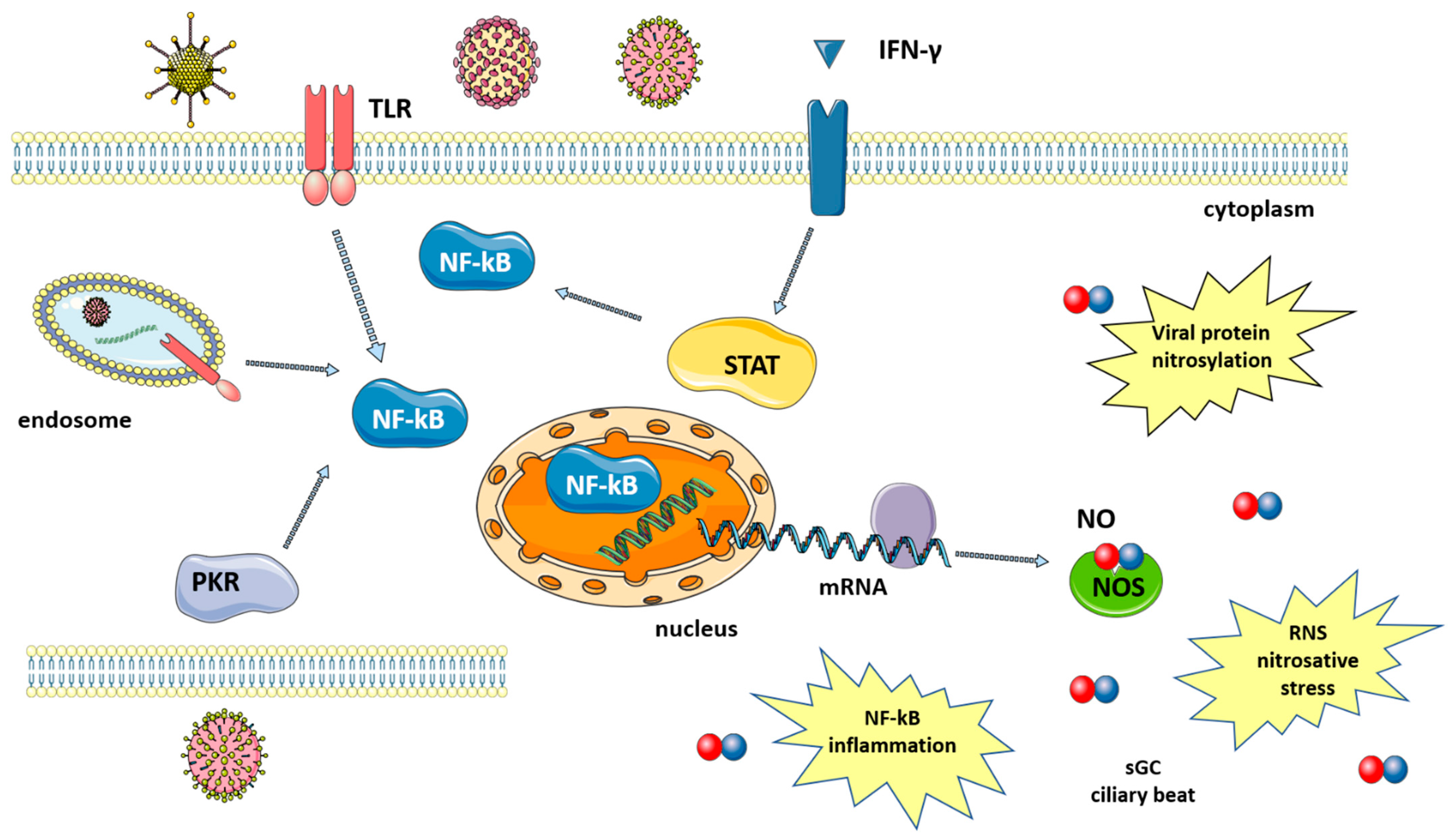
3. Role of NO in Immune Defense against Viruses
4. Role of NO in Vascular Endothelium
Vascular Endothelium Dysfunctions Associated with Viral Infections
5. NO and the Respiratory System
6. NO-Based Antiviral Strategies
6.1. gNO Inhalation
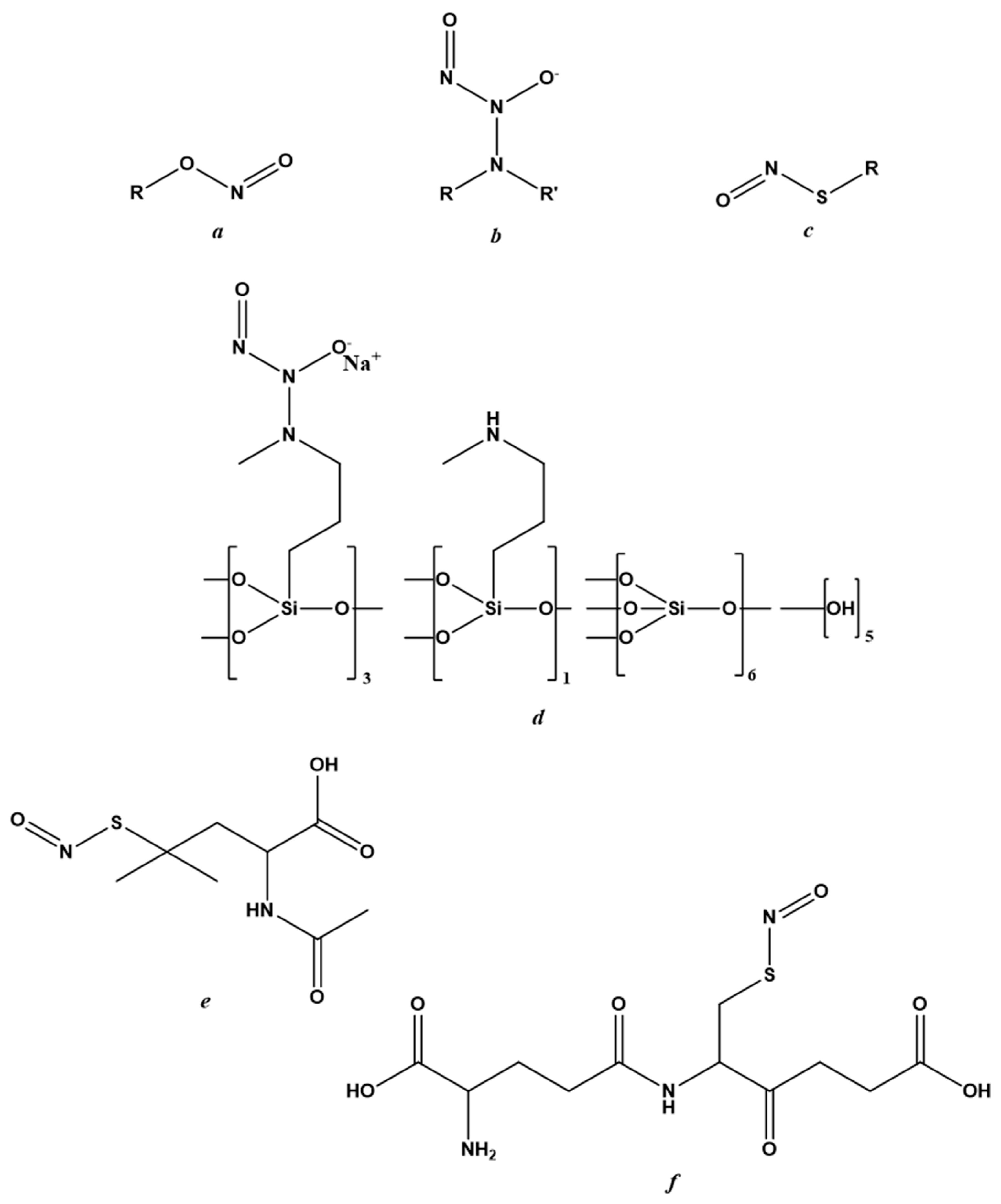
6.2. NO-Donors
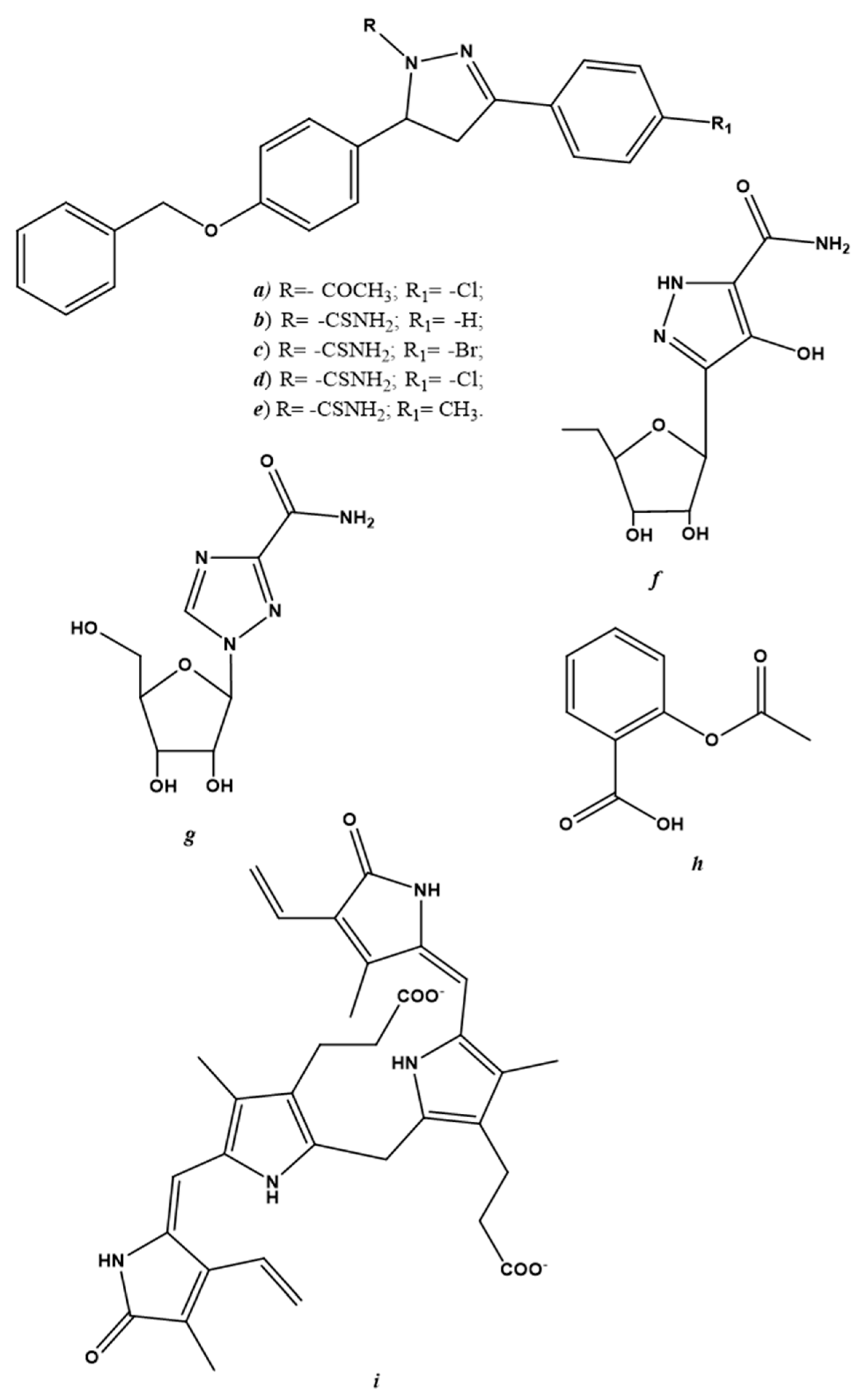
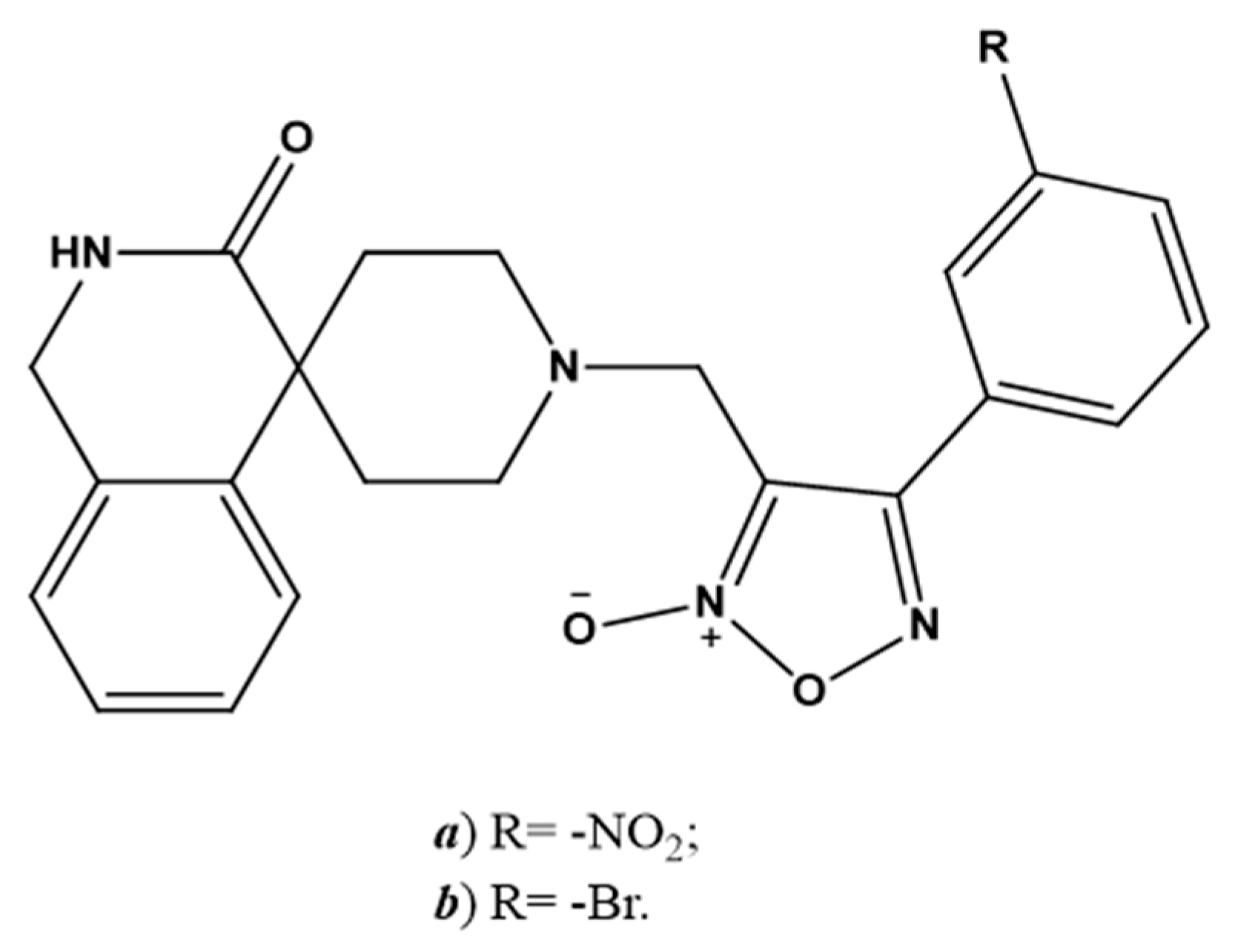
6.3. Endogenous NO-Regulating Drugs
7. Therapeutic Potential of NO for the Treatment of COVID-19
7.1. Short Outline of SARS-CoV-2
7.2. Endogenous NO in COVID-19 Patients
7.3. Applications of gNO-Based Therapy in COVID-19 Patients
7.4. NO-Donors in COVID-19 Patients
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nicola, M.; Alsafi, Z.; Sohrabi, C.; Kerwan, A.; Al-Jabir, A.; Iosifidis, C.; Agha, M.; Agha, R. The socio-economic implications of the coronavirus pandemic (COVID-19): A review. Int. J. Surg. 2020, 78, 185–193. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E.; Li, G. Approved antiviral drugs over the past 50 years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur-Marzec, H.; Cegłowska, M.; Konkel, R.; Pyrć, K. Antiviral Cyanometabolites—A Review. Biomolecules 2021, 11, 474. [Google Scholar] [CrossRef] [PubMed]
- Boroumand, H.; Badie, F.; Mazaheri, S.; Seyedi, Z.S.; Nahand, J.S.; Nejati, M.; Baghi, H.B.; Abbasi-Kolli, M.; Badehnoosh, B.; Ghandali, M.; et al. Chitosan-Based Nanoparticles Against Viral Infections. Front. Cell. Infect. Microbiol. 2021, 11, 643953. [Google Scholar] [CrossRef]
- Lei, J.; Vodovotz, Y.; Tzeng, E.; Billiar, T.R. Nitric oxide, a protective molecule in the cardiovascular system. Nitric Oxide 2013, 35, 175–185. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Jones, M.L.; Ganopolsky, J.G.; Labbé, A.; Wahl, C.; Prakash, S. Antimicrobial properties of nitric oxide and its application in antimicrobial formulations and medical devices. Appl. Microbiol. Biotechnol. 2010, 88, 401–407. [Google Scholar] [CrossRef]
- Garren, M.R.; Ashcraft, M.; Qian, Y.; Douglass, M.; Brisbois, E.J.; Handa, H. Nitric oxide and viral infection: Recent developments in antiviral therapies and platforms. Appl. Mater Today. 2021, 22, 100887. [Google Scholar] [CrossRef]
- Wink, D.A.; Hines, H.B.; Cheng, R.Y.S.; Switzer, C.H.; Flores-Santana, W.; Vitek, M.P.; Ridnour, L.A.; Colton, C.A. Nitric oxide and redox mechanisms in the immune response. J. Leukoc. Biol. 2011, 89, 873–891. [Google Scholar] [CrossRef] [Green Version]
- Dallavalle, S.; Dobričić, V.; Lazzarato, L.; Gazzano, E.; Machuqueiro, M.; Pajeva, I.; Tsakovska, I.; Zidar, N.; Fruttero, R. Im-provement of conventional anti-cancer drugs as new tools against multidrug resistant tumors. Drug Resist. Updat. 2020, 50, 100682. [Google Scholar] [CrossRef]
- Džoljić, E.; Grbatinić, I.; Kostić, V. Why is nitric oxide important for our brain? Funct. Neurol. 2015, 30, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Mattila, J.T.; Thomas, A. Nitric Oxide Synthase: Non-Canonical Expression Patterns. Front. Immunol. 2014, 5, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleli, M.; Zollbrecht, C.; Montenegro, M.F.; Hezel, M.; Zhong, J.; Persson, E.G.; Holmdahl, R.; Weitzberg, E.; Lundberg, J.O.; Carlström, M. Enhanced XOR activity in eNOS-deficient mice: Effects on the nitrate-nitrite-NO pathway and ROS homeostasis. Free Radic Biol. Med. 2016, 99, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.; Jiang, J.; Su, L.; Shu, T.; Liu, H.; Lai, S.; Ghiladi, R.A.; Wang, J. The role of NO in COVID-19 and potential therapeutic strategies. Free Radic. Biol. Med. 2020, 163, 153–162. [Google Scholar] [CrossRef]
- Lisi, F.; Zelikin, A.N.; Chandrawati, R. Nitric Oxide to Fight Viral Infections. Adv. Sci. 2021, 8, 2003895. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Chen, L.; Maes, P.; Hedenstierna, G.; Van Ranst, M. Inhibition of SARS-coronavirus infection in vitro by S-nitroso-N-acetylpenicillamine, a nitric oxide donor compound. Int. J. Infect. Dis. 2004, 8, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Åkerström, S.; Mousavi-Jazi, M.; Klingström, J.; Leijon, M.; Lundkvist, A.; Mirazimi, A. Nitric Oxide Inhibits the Replication Cycle of Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2005, 79, 1966–1969. [Google Scholar] [CrossRef] [Green Version]
- Akerström, S.; Gunalan, V.; Keng, C.T.; Tan, Y.J.; Mirazimi, A. Dual effect of nitric oxide on SARS-CoV replication: Viral RNA production and palmitoylation of the S protein are affected. Virology 2009, 395, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Cader, M.S.; Amarasinghe, A.; Abdul-Careem, M.F. Activation of toll-like receptor signaling pathways leading to nitric oxide-mediated antiviral responses. Arch. Virol. 2016, 161, 2075–2086. [Google Scholar] [CrossRef]
- Xu, W.; Zheng, S.; Dweik, R.A.; Erzurum, S.C. Role of epithelial nitric oxide in airway viral infection. Free Radic. Biol. Med. 2006, 41, 19–28. [Google Scholar] [CrossRef]
- Auch, C.J.; Saha, R.; Sheikh, F.G.; Liu, X.; Jacobs, B.L.; Pahan, K. Role of protein kinase R in double-stranded RNA-induced expression of nitric oxide synthase in human astroglia. FEBS Lett. 2004, 563, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Bayarri, M.A.; Milara, J.; Estornut, C.; Cortijo, J. Nitric Oxide System and Bronchial Epithelium: More Than a Barrier. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Akaberi, D.; Krambrich, J.; Ling, J.; Luni, C.; Hedenstierna, G.; Järhult, J.D.; Lennerstrand, J.; Lundkvist, Å. Mitigation of the replication of SARS-CoV-2 by nitric oxide in vitro. Redox. Biol. 2020, 37, 101734. [Google Scholar] [CrossRef] [PubMed]
- Uehara, E.U.; Shida Bde, S.; De Brito, C.A. Role of nitric oxide in immune responses against viruses: Beyond microbicidal ac-tivity. Inflamm Res. 2015, 64, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Jean, D.; Maitre, B.; Tankovic, J.; Meignan, M.; Adnot, S.; Brun-Buisson, C.; Harf, A.; Delclaux, C. Beneficial effects of nitric oxide inhalation on pulmonary bacterial clearance. Crit. Care Med. 2002, 30, 442–447. [Google Scholar] [CrossRef]
- Wyatt, T.A.; Spurzem, J.R.; May, K.; Sisson, J.H. Regulation of ciliary beat frequency by both PKA and PKG in bovine airway epithelial cells. Am. J. Physiol. Cell. Mol. Physiol. 1998, 275, L827–L835. [Google Scholar] [CrossRef] [PubMed]
- Pober, J.S.; Sessa, W.C. Evolving functions of endothelial cells in inflammation. Nat. Rev. Immunol. 2007, 7, 803–815. [Google Scholar] [CrossRef]
- Perrone, L.A.; Belser, J.A.; Wadford, D.A.; Katz, J.M.; Tumpey, T.M. Inducible Nitric Oxide Contributes to Viral Pathogenesis Following Highly Pathogenic Influenza Virus Infection in Mice. J. Infect. Dis. 2013, 207, 1576–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torre, D.; Ferrario, G. Immunological aspects of nitric oxide in HIV-1 infection. Med Hypotheses 1996, 47, 405–407. [Google Scholar] [CrossRef]
- Klinger, J.R.; Kadowitz, P.J. The Nitric Oxide Pathway in Pulmonary Vascular Disease. Am. J. Cardiol. 2017, 120, S71–S79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.Y.; Ye, Z.X.; Wang, X.F.; Chang, J.; Yang, M.W.; Zhong, H.H.; Hong, F.F.; Yang, S.L. Nitric oxide bioavailability dys-function involves in atherosclerosis. Biomed. Pharmacother. 2018, 97, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-H.; Park, W.J. Endothelial Dysfunction: Clinical Implications in Cardiovascular Disease and Therapeutic Approaches. J. Korean Med. Sci. 2015, 30, 1213–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loscalzo, J. Nitric oxide insufficiency, platelet activation, and arterial thrombosis. Circ. Res. 2001, 88, 756–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marincowitz, C.; Genis, A.; Goswami, N.; De Boever, P.; Nawrot, T.S.; Strijdom, H. Vascular endothelial dysfunction in the wake of HIV and ART. FEBS J. 2019, 286, 1256–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fosse, J.H.; Haraldsen, G.; Falk, K.; Edelmann, R. Endothelial Cells in Emerging Viral Infections. Front. Cardiovasc. Med. 2021, 8, 619690. [Google Scholar] [CrossRef]
- Ritz, T.; Salsman, M.L.; Young, D.A.; Lippert, A.R.; Khan, D.A.; Ginty, A.T. Boosting nitric oxide in stress and respiratory in-fection: Potential relevance for asthma and COVID-19. Brain Behav. Immun. Health. 2021, 14, 100255. [Google Scholar] [CrossRef] [PubMed]
- Dominic, P.; Ahmad, J.; Bhandari, R.; Pardue, S.; Solorzano, J.; Jaisingh, K.; Watts, M.; Bailey, S.R.; Orr, A.W.; Kevil, C.G.; et al. Decreased availability of nitric oxide and hydrogen sulfide is a hallmark of COVID-19. Redox Biol. 2021, 43, 101982. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, B.; Yazici, A. Could the decrease in the endothelial nitric oxide (NO) production and NO bioavailability be the crucial cause of COVID-19 related deaths? Med. Hypotheses. 2020, 144, 109970. [Google Scholar] [CrossRef]
- Berkman, S.A.; Tapson, V.F. COVID-19 and Its Implications for Thrombosis and Anticoagulation. Semin. Respir. Crit. Care Med. 2021, 42, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Friebe, A.; Englert, N. NO—Sensitive guanylyl cyclase in the lung. J. Cereb. Blood Flow Metab. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bolotina, V.M.; Najibi, S.; Palacino, J.J.; Pagano, P.J.; Cohen, R.A. Nitric oxide directly activates calcium-dependent potassium channels in vascular smooth muscle. Nature 1994, 368, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Ricciardolo, F.L.M. Multiple roles of nitric oxide in the airways. Thorax 2003, 58, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antosova, M.; Mokra, D.; Pepucha, L.; Plevkova, J.; Buday, T.; Sterusky, M.; Bencova, A. Physiology of Nitric Oxide in the Respiratory System. Physiol. Res. 2017, 66, S159–S172. [Google Scholar] [CrossRef] [PubMed]
- Price, M.E.; Sisson, J.H. Redox regulation of motile cilia in airway disease. Redox Biol. 2019, 27, 101146. [Google Scholar] [CrossRef]
- Mehta, P.P.; Dhapte-Pawar, V.S. Novel and Evolving Therapies for COVID-19 Related Pulmonary Complications. Am. J. Med. Sci. 2021, 361, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Llorens, S.; Nava, E. Cardiovascular diseases and the nitric oxide pathway. Curr. Vasc. Pharmacol. 2003, 1, 335–346. [Google Scholar] [CrossRef]
- Ignarro, L.J.; Napoli, C.; Loscalzo, J. Nitric Oxide Donors and Cardiovascular Agents Modulating the Bioactivity of Nitric Oxide. Circ. Res. 2002, 90, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Reiss, C.S.; Komatsu, T. Does nitric oxide play a critical role in viral infections? J. Virol. 1998, 72, 4547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMullin, B.B.; Chittock, D.R.; Roscoe, D.L.; Garcha, H.; Wang, L.; Miller, C.C. The antimicrobial effect of nitric oxide on the bacteria that cause nosocomial pneumonia in mechanically ventilated patients in the intensive care unit. Respir. Care 2005, 50, 1451–1456. [Google Scholar] [PubMed]
- Chen, L.; Liu, P.; Gao, H.; Sun, B.; Chao, D.; Wang, F.; Zhu, Y.; Hedenstierna, G.; Wang, C.G. Inhalation of Nitric Oxide in the Treatment of Severe Acute Respiratory Syndrome: A Rescue Trial in Beijing. Clin. Infect. Dis. 2004, 39, 1531–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyring, S.K.; Rosen, T.; Berman, B.; Stasko, N.; Durham, T.; Maeda-Chubachi, T. A Phase 2 Controlled Study of SB206, a Topical Nitric Oxide-Releasing Drug for Extragenital Wart Treatment. J. Drugs Dermatol. 2018, 17, 1100–1105. [Google Scholar]
- Ormerod, A.; White, M.; Shah, S.; Benjamin, N. Molluscum contagiosum effectively treated with a topical acidified nitrite, nitric oxide liberating cream. Br. J. Dermatol. 1999, 141, 1051–1053. [Google Scholar] [CrossRef]
- NOVAN. Novan Reports Top-Line Efficacy Results from Phase 3 Trials of SB206 for Molluscum Contagiosum. 2020. Available online: https://www.globenewswire.com/news-release/2020/01/02/1965841/0/en/Novan-Reports-Top-Line-Efficacy-Results-from-phase-3-Trials-of-SB206-for-Molluscum-Contagiosum.html (accessed on 17 February 2022).
- Darwish, I.; Miller, C.; Kain, K.; Liles, W.C. Inhaled Nitric Oxide Therapy Fails to Improve Outcome in Experimental Severe Influenza. Int. J. Med. Sci. 2012, 9, 157–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Wen, L.; Xiao, Q.; He, K. Nitric oxide-generating compound GSNO suppresses porcine circovirus type 2 infection in vitro and in vivo. BMC Veter. Res. 2016, 13, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regev-Shoshani, G.; Vimalanathan, S.; McMullin, B.; Road, J.; Av-Gay, Y.; Miller, C. Gaseous nitric oxide reduces influenza infectivity in vitro. Nitric Oxide 2013, 31, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Falk, K.; Lundkvist, Å.; Mirazimi, A. Exogenous nitric oxide inhibits Crimean Congo hemorrhagic fever virus. Virus Res. 2006, 120, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Klingstrom, J.; Åkerström, S.; Hardestam, J.; Stoltz, M.; Simon, M.; Falk, K.I.; Mirazimi, A.; Rottenberg, M.; Lundkvist, Å. Nitric oxide and peroxynitrite have different antiviral effects against hantavirus replication and free mature virions. Eur. J. Immunol. 2006, 36, 2649–2657. [Google Scholar] [CrossRef] [PubMed]
- Naimi, E.; Zhou, A.; Khalili, P.; Wiebe, L.I.; Balzarini, J.; De Clercq, A.E.; Knaus, E.E. Synthesis of 3‘- and 5‘-Nitrooxy Pyrimidine Nucleoside Nitrate Esters: “Nitric Oxide Donor” Agents for Evaluation as Anticancer and Antiviral Agents. J. Med. Chem. 2003, 46, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Baars, M.M.J.W.; de Lijster, P.; Fouchier, R.; Osterhaus, A.D.M.E. Inhibition of Influenza Virus Replication by Nitric Oxide. J. Virol. 1999, 73, 8880–8883. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, N.S.; Moore, D.W.; Wang, H.-K.; Broker, T.R.; Chow, L.T. NVN1000, a novel nitric oxide-releasing compound, inhibits HPV-18 virus production by interfering with E6 and E7 oncoprotein functions. Antivir. Res. 2019, 170, 104559. [Google Scholar] [CrossRef]
- Koetzler, R.; Zaheer, R.S.; Newton, R.; Proud, D. Nitric oxide inhibits IFN regulatory factor 1 and nuclear factor-kappaB pathways in rhinovirus-infected epithelial cells. J. Allergy Clin. Immunol. 2009, 124, 551. [Google Scholar] [CrossRef] [PubMed]
- Koetzler, R.; Zaheer, R.S.; Wiehler, S.; Holden, N.; Giembycz, M.A.; Proud, D. Nitric oxide inhibits human rhinovirus-induced transcriptional activation of CXCL10 in airway epithelial cells. J. Allergy Clin. Immunol. 2009, 123, 201–208.e9. [Google Scholar] [CrossRef] [PubMed]
- Sanders, S.P.; Siekierski, E.S.; Porter, J.D.; Richards, S.M.; Proud, D. Nitric oxide inhibits rhinovirus-induced cytokine produc-tion and viral replication in a human respiratory epithelial cell line. J. Virol. 1998, 72, 934. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.I.; Kao, S.J.; Wang, D.; Lee, R.P.; Su, C.F. Acute respiratory distress syndrome. J. Biomed. Sci. 2003, 10, 588. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.P.; Wang, D.; Kao, S.J.; Chen, H.I. The lung is the major site that produces nitric oxide to induce acute pulmonary oe-dema in endotoxin shock. Clin. Exp. Pharmacol. Physiol. 2001, 8, 315. [Google Scholar] [CrossRef]
- Kao, S.J.; Peng, T.C.; Lee, R.P.; Hsu, K.; Chen, C.F.; Hung, Y.K.; Wang, D.; Chen, H.I. Nitric oxide mediates lung injury induced by ischemia-reperfusion in rats. J. Biomed. Sci. 2003, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Gebistorf, F.; Karam, O.; Wetterslev, J.; Afshari, A. Inhaled nitric oxide for acute respiratory distress syndrome (ARDS) in children and adults. Cochrane Database Syst. Rev. 2016, 2016, CD002787. [Google Scholar] [CrossRef] [PubMed]
- Karam, O.; Gebistorf, F.; Wetterslev, J.; Afshari, A. The effect of inhaled nitric oxide in acute respiratory distress syndrome in children and adults: A Cochrane Systematic Review with trial sequential analysis. Anaesthesia 2016, 72, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.; McMullin, B.; Ghaffari, A.; Stenzler, A.; Pick, N.; Roscoe, D.; Ghahary, A.; Road, J.; Av-Gay, Y. Gaseous nitric oxide bactericidal activity retained during intermittent high-dose short duration exposure. Nitric Oxide 2009, 20, 16–23. [Google Scholar] [CrossRef]
- Long, R.; Jones, R.; Talbot, J.; Mayers, I.; Barrie, J.; Hoskinson, M.; Light, B. Inhaled Nitric Oxide Treatment of Patients with Pulmonary Tuberculosis Evidenced by Positive Sputum Smears. Antimicrob. Agents Chemother. 2005, 49, 1209–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, T.S.; Sugimori, K.; Camporesi, E.M.; Anderson, G. Half-life of nitric oxide in aqueous solutions with and without haemoglobin. Physiol. Meas. 1996, 17, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, M.W.; Kuo, L.; Liao, J. Estimation of nitric oxide production and reaction rates in tissue by use of a mathematical model. Am. J. Physiol. Content 1998, 274, H2163. [Google Scholar]
- Hall, J.R.; Rouillard, K.R.; Suchyta, D.J.; Brown, M.D.; Ahonen, M.J.R.; Schoenfisch, M.H. Mode of Nitric Oxide Delivery Af-fects Antibacterial Action. ACS Biomater. Sci. Eng. 2019, 6, 433. [Google Scholar] [CrossRef]
- Rouillard, K.R.; Hill, D.B.; Schoenfisch, M.H. Antibiofilm and mucolytic action of nitric oxide delivered via gas or macromo-lecular donor using in vitro and ex vivo models. J. Cyst. Fibros. 2020, 19, 1004–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, A.A.; Siegfried, E.C.; Durham, T.; De León, E.N.; Reams, T.; Messersmith, E.; Maeda-Chubachi, T. Efficacy and tol-erability of an investigational nitric oxide–releasing topical gel in patients with molluscum contagiosum: A randomized clinical trial. J. Am. Acad. Dermatol. 2020, 82, 887. [Google Scholar] [CrossRef] [Green Version]
- Croen, K.D. Evidence for antiviral effect of nitric oxide. Inhibition of herpes simplex virus type 1 replication. J. Clin. Investig. 1993, 91, 2446–2452. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, C.; Ocampo, C.J.; Saura, M.; McMillan, A.; Lowenstein, C.J. Nitric oxide inhibition of coxsackievirus replication in vitro. J. Clin. Investig. 1997, 100, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Huang, Y.L.; Ma, S.H.; Yeh, C.T.; Chiou, S.Y.; Chen, L.K.; Liao, C.L. Inhibition of Japanese encephalitis virus infection by nitric oxide: Antiviral effect of nitric oxide on RNA virus replication. J. Virol. 1997, 71, 5227–5235. [Google Scholar] [CrossRef] [Green Version]
- Riccio, D.A.; Schoenfisch, M.H. Nitric oxide release: Part I. Macromolecular scaffolds. Chem. Soc. Rev. 2012, 41, 3731–3741. [Google Scholar] [CrossRef]
- Carpenter, A.W.; Schoenfisch, M.H. Nitric oxide release: Part II. Therapeutic applications. Chem. Soc. Rev. 2012, 41, 3742–3752. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, S.P.; Pant, J.; Goudie, M.J.; Schmiedt, C.; Handa, H. Achieving Long-Term Biocompatible Silicone via Covalently Immobilized S-Nitroso-N-acetylpenicillamine (SNAP) That Exhibits 4 Months of Sustained Nitric Oxide Release. ACS Appl. Mater. Interfaces 2018, 10, 27316–27325. [Google Scholar] [CrossRef] [PubMed]
- Lautner, G.; Meyerhoff, M.E.; Schwendeman, S.P. Biodegradable poly(lactic-coglycolic acid) microspheres loaded with S-nitroso-N-acetyl-D-penicillamine for controlled nitric oxide delivery. J. Control. Release 2016, 225, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Pan, S.; Mukherjee, A.; De, P. Nitric Oxide Releasing Delivery Platforms: Design, Detection, Biomedical Applications, and Future Possibilities. Mol. Pharm. 2021, 18, 3181–3205. [Google Scholar] [CrossRef] [PubMed]
- Estes, L.M.; Singha, P.; Singh, S.; Sakthivel, T.S.; Garren, M.; Devine, R.; Brisbois, E.J.; Seal, S.; Handa, H. Characterization of a nitric oxide (NO) donor molecule and cerium oxide nanoparticle (CNP) interactions and their synergistic antimicrobial potential for biomedical applications. J. Colloid Interface Sci. 2020, 586, 163–177. [Google Scholar] [CrossRef]
- Gao, S.; Zhang, W.; Wang, R.; Hopkins, S.P.; Spagnoli, J.C.; Racin, M.; Bai, L.; Li, L.; Jiang, W.; Yang, X.; et al. Nanoparticles encapsulating nitrosylated maytansine to enhance radi-ation therapy. ACS Nano 2020, 14, 1468. [Google Scholar] [CrossRef]
- Singha, P.; Workman, C.D.; Pant, J.; Hopkins, S.P.; Handa, H. Zinc-oxide nanoparticles act catalytically and synergistically with nitric oxide donors to enhance antimicrobial efficacy. J. Biomed. Mater. Res. Part A 2019, 107, 1425–1433. [Google Scholar] [CrossRef]
- Quinn, J.; Whittaker, M.; Davis, T.P. Delivering nitric oxide with nanoparticles. J. Control. Release 2015, 205, 190–205. [Google Scholar] [CrossRef]
- Ansari, A.; Ali, A.; Asif, M.; Shamsuzzaman. Review: Biologically active pyrazole derivatives. New J. Chem. 2017, 41, 16–41. [Google Scholar] [CrossRef]
- El-Sabbagh, O.I.; Baraka, M.M.; Ibrahim, S.M.; Pannecouque, C.; Andrei, G.; Snoeck, R.; Balzarini, J.; Rashad, A.A. Synthesis and antiviral activity of new pyrazole and thiazole derivatives. Eur. J. Med. Chem. 2009, 44, 3746–3753. [Google Scholar] [CrossRef]
- Karrouchi, K.; Radi, S.; Ramli, Y.; Taoufik, J.; Mabkhot, Y.N.; Al-aizari, F.A.; Ansar, M. Synthesis and Pharmacological Ac-tivities of Pyrazole Derivatives: A Review. Molecules 2018, 23, 134. [Google Scholar] [CrossRef] [Green Version]
- Wohl, B.M.; Smith, A.A.A.; Kryger, M.B.L.; Zelikin, A.N. Narrow Therapeutic Window of Ribavirin as an Inhibitor of Nitric Oxide Synthesis is Broadened by Macromolecular Prodrugs. Biomacromolecules 2013, 14, 3916–3926. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Murillo, K.; Rincón-Sánchez, A.R.; Martínez-Rodríguez, H.; Bosques-Padilla, F.; Ramos-Jiménez, J.; Barrera-Saldaña, H.A.; Rojkind, M.; Rivas-Estilla A., M. Acetylsalicylic acid inhibits hepatitis C virus RNA and protein expression through cy-clooxygenase 2 signaling pathways. Hepatology 2008, 47, 1462–1472. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Ibarra, C.P.; Lozano-Sepulveda, S.; Munoz, L.; Rincón-Sánchez, A.R.; Cordova-Fletes, C.; Rivas-Estilla, A.M.G. Downregulation of inducible nitric oxide synthase (iNOS) expression is implicated in the antiviral activity of acetylsalicylic acid in HCV-expressing cells. Arch. Virol. 2014, 159, 3321–3328. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, R.; Mancuso, C.M.; Marchetti, S.; Di Stasio, E.; Pani, G.; Fadda, G. Bilirubin: An Endogenous Molecule with Antiviral Activity in vitro. Front. Pharmacol. 2012, 3, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [PubMed] [Green Version]
- Adusumilli, N.C.; Zhang, D.; Friedman, J.M.; Friedman, A.J. Harnessing nitric oxide for preventing, limiting and treating the severe pulmonary consequences of COVID-19. Nitric Oxide 2020, 103, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Abe, R.; Hattori, N.; Matsumura, Y.; Oshima, T.; Taniguchi, T.; Igari, H.; Nakada, T.-A. Clinical course of a critically ill patient with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). J. Artif. Organs 2020, 23, 397–400. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904. [Google Scholar] [CrossRef]
- Mir, J.M.; Maurya, R.C. Nitric oxide as a therapeutic option for COVID-19 treatment: A concise perspective. New J. Chem. 2020, 45, 1774–1784. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Li, H.; Liu, Z.; Ge, J. Scientific research progress of COVID-19/SARS-CoV-2 in the first five months. J. Cell Mol. Med. 2020, 24, 6558–6570. [Google Scholar] [CrossRef]
- Banu, N.; Panikar, S.S.; Leal, L.R.; Leal, A.R. Protective role of ACE2 and its downregulation in SARS-CoV-2 infection leading to Macrophage Activation Syndrome: Therapeutic implications. Life Sci. 2020, 256, 117905. [Google Scholar] [CrossRef] [PubMed]
- Boscá, L.; Zeini, M.; Traves, P.G.; Hortelano, S. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S. Coronavirus (Covid-19) sepsis: Revisiting mitochondrial dysfunction in pathogenesis, aging, inflammation, and mortality. Agents Actions 2020, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Urso, C.; Caimi, G. Oxidative stress and endothelial dysfunction. Minerva Med. 2011, 102, 59–77. [Google Scholar]
- Martel, J.; Ko, Y.F.; Young, J.D.; Ojcius, D.M. Could nasal nitric oxide help to mitigate the severity of COVID-19? Microbes Infect. 2020, 22, 168–171. [Google Scholar] [CrossRef]
- Berra, L.; Lei, C.; Su, B.; Dong, H.; Fakhr, B.S.; Grassi, L.G.; Di Fenza, R.; Gianni, S.; Pinciroli, R.; Vassena, E.; et al. Protocol for a randomized controlled trial testing inhaled nitric oxide therapy in spontaneously breathing patients with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Lei, C.; Su, B.; Dong, H.; Bellavia, A.; Di Fenza, R.; Fakhr, B.S.; Gianni, S.; Grassi, L.G.; Kacmarek, R.; Morais, C.C.A.; et al. Protocol of a randomized controlled trial testing inhaled Nitric Oxide in mechanically ventilated pa-tients with severe acute respiratory syndrome in COVID-19 (SARS-CoV-2). medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Gianni, S.; Fakhr, B.S.; Morais, C.C.A.; Di Fenza, R.; Larson, G.; Pinciroli, R.; Houle, T.; Mueller, A.L.; Bellavia, A.; Kacmarek, R.; et al. Nitric oxide gas inhalation to prevent COVID-2019 in healthcare providers. medRxiv 2020. [Google Scholar] [CrossRef]
- Road, J. Inhaled Gaseous Nitric Oxide (gNO) Antimicrobial Treatment of Difficult Bacterial and Viral Lung (COVID-19) Infections (NCT03331445); Nitric Solutions-Mobile Unit: Vancouver, BC, Canada, 2020. [Google Scholar]
- Al-Sehemi, A.G.; Pannipara, M.; Parulekar, R.S.; Patil, O.; Choudhari, P.B.; Bhatia, M.S.; Zubaidha, P.K.; Tamboli, Y. Potential of NO donor furoxan as SARS-CoV-2 main protease (M pro) inhibitors: In silico analysis. J. Biomol. Struct. Dyn. 2020, 39, 5804–5818. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Trials | |||
| gNO Treatment | |||
| Model | NO Dosage (ppm) | Duration | References |
| SARS Coronavirus | <30 (stepwise) | 3–7 days | [50] |
| NO Donor Treatment | |||
| Model | NO Dosage (ppm) | Duration | References |
| Human Papillomavirus | Not specified | 1–2 daily, 12 weeks | [51] |
| Molluscum Contagiosum | 5% acidified nitrite donor in cream | 1–83 months | [52] |
| Not specified | 1–2 daily, 12 weeks | [53] | |
| In-Vivo Studies | |||
| gNO Treatment | |||
| Model | NO Dosage (ppm) | Duration | References |
| Human Influenza | 80/160 | Continuous/Intermittent | [54] |
| NO Donor Treatment | |||
| Model | NO Dosage (mM) | Duration | References |
| Porcine Circovirus Type 2 | 10 | 1 daily, 6 days post-infection | [55] |
| In-Vitro and Ex-Vivo Studies | |||
| gNO Treatment | |||
| Model | NO Dosage (ppm) | Duration (hrs) | References |
| Human Influenza | 80/160 | 3 | [56] |
| Nosocomial pneumonia-related clinical isolates | 200 | 2/6 | [49] |
| NO Donor Treatment | |||
| Model | NO Dosage (μM) | Duration (hrs) | References |
| Crimean Congo Hemorrhagic Fever Virus | 50–400 | 24 | [57] |
| Hantaan Hantavirus | 100 | 12 | [58] |
| Herpes Simplex Virus and Vaccinia Virus | 1000 | 12 | [59] |
| Human Influenza | 50–400 | 24 | [60] |
| Human Papillomavirus | Not specified | 1 | [61] |
| Human Rhinovirus | 100–1000 | 4/24 | [62,63,64] |
| SARS Coronavirus-1 | 65–500 | 72 | [16] |
| SARS Coronavirus-2 | 20–500 | 36–72 | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sodano, F.; Gazzano, E.; Fruttero, R.; Lazzarato, L. NO in Viral Infections: Role and Development of Antiviral Therapies. Molecules 2022, 27, 2337. https://doi.org/10.3390/molecules27072337
Sodano F, Gazzano E, Fruttero R, Lazzarato L. NO in Viral Infections: Role and Development of Antiviral Therapies. Molecules. 2022; 27(7):2337. https://doi.org/10.3390/molecules27072337
Chicago/Turabian StyleSodano, Federica, Elena Gazzano, Roberta Fruttero, and Loretta Lazzarato. 2022. "NO in Viral Infections: Role and Development of Antiviral Therapies" Molecules 27, no. 7: 2337. https://doi.org/10.3390/molecules27072337