
Effects of Intra-BLA Administration of PPAR Antagonists on Formalin-Evoked Nociceptive Behaviour, Fear-Conditioned Analgesia, and Conditioned Fear in the Presence or Absence of Nociceptive Tone in Rats

 ,
,  and
and 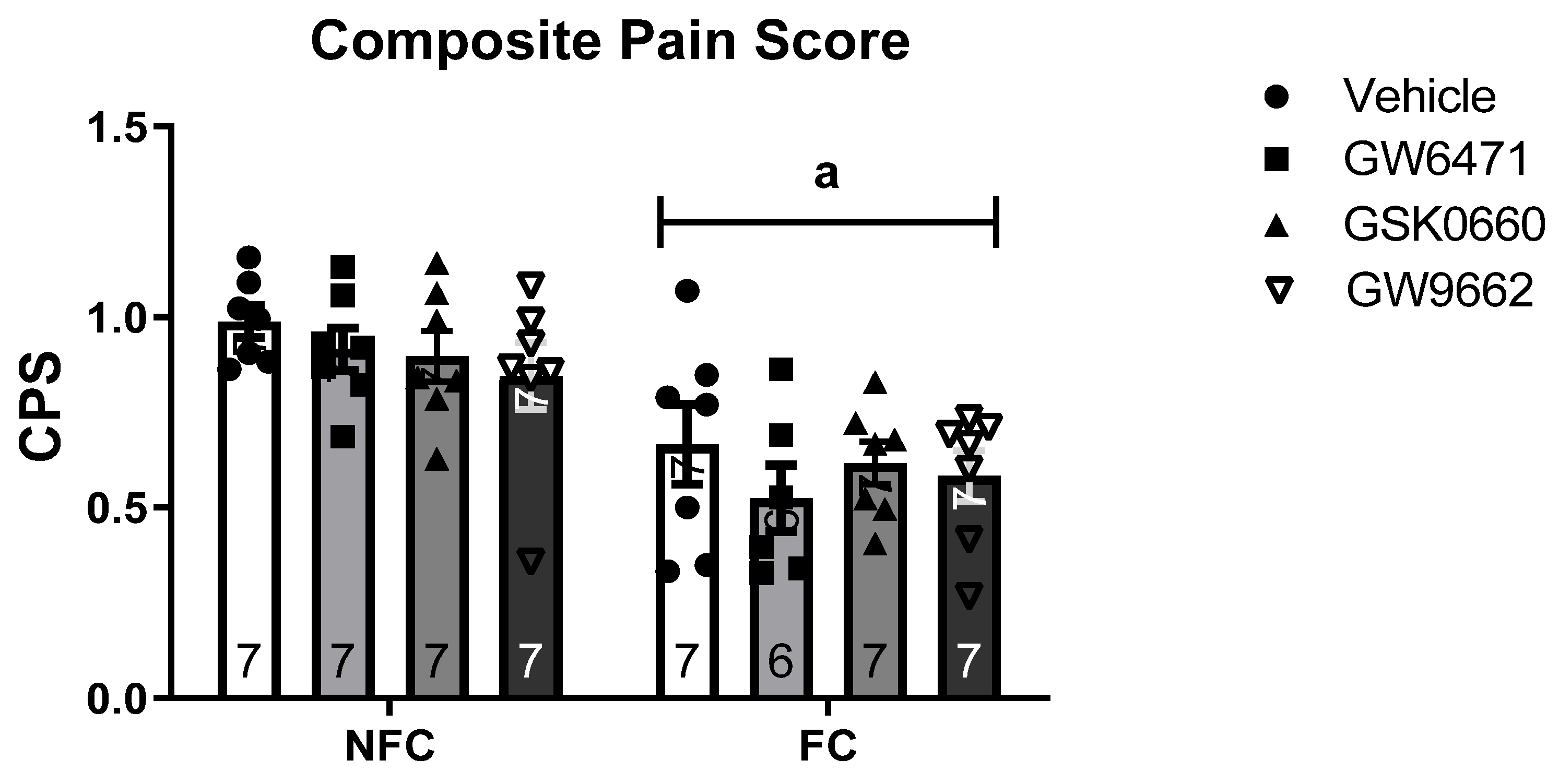{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Experiment 1: Effects of Intra-BLA Administration of PPAR Antagonists on Formalin-Evoked Nociceptive Behaviour, Fear-Conditioned Analgesia, and Conditioned Fear in the Presence of Nociceptive Tone in Rats
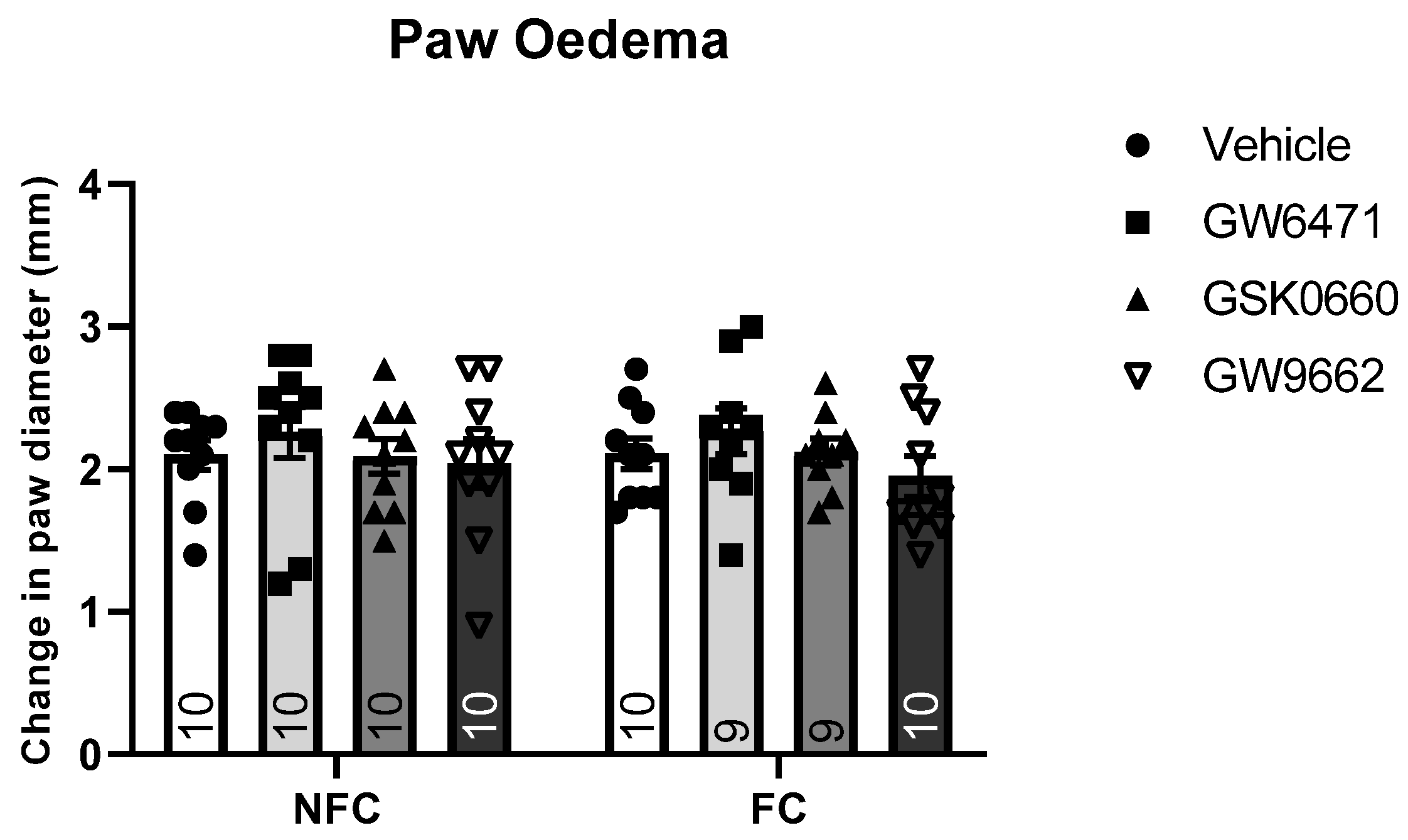
2.1.1. Intra-BLA Administration of GW6471, GSK0660, and GW9662 Had No Effect on Formalin-Evoked Nociceptive Behaviour or FCA
2.1.2. Intra-BLA Administration of GW6471, GSK0660, and GW9662 Increases Fear-Related Behaviour in Formalin-Treated Rats
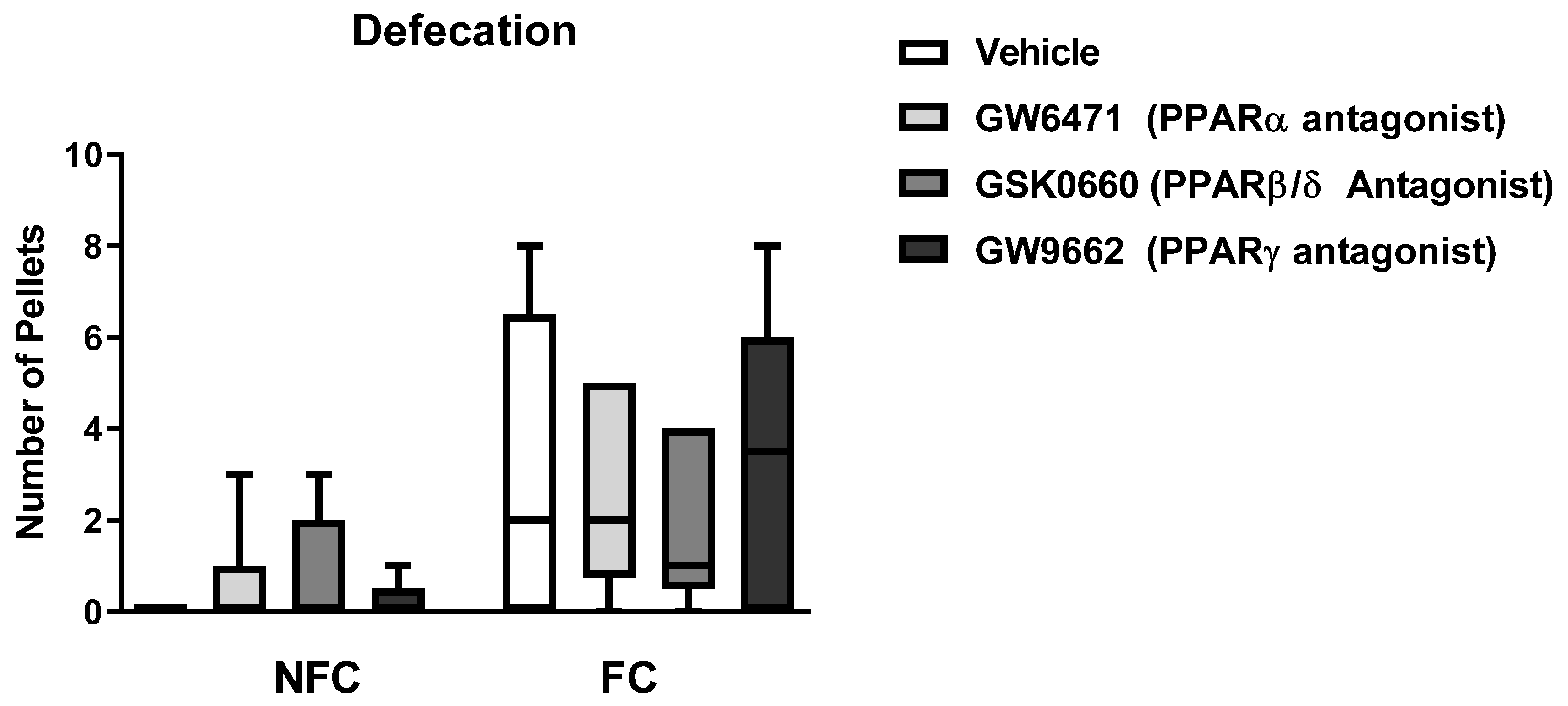
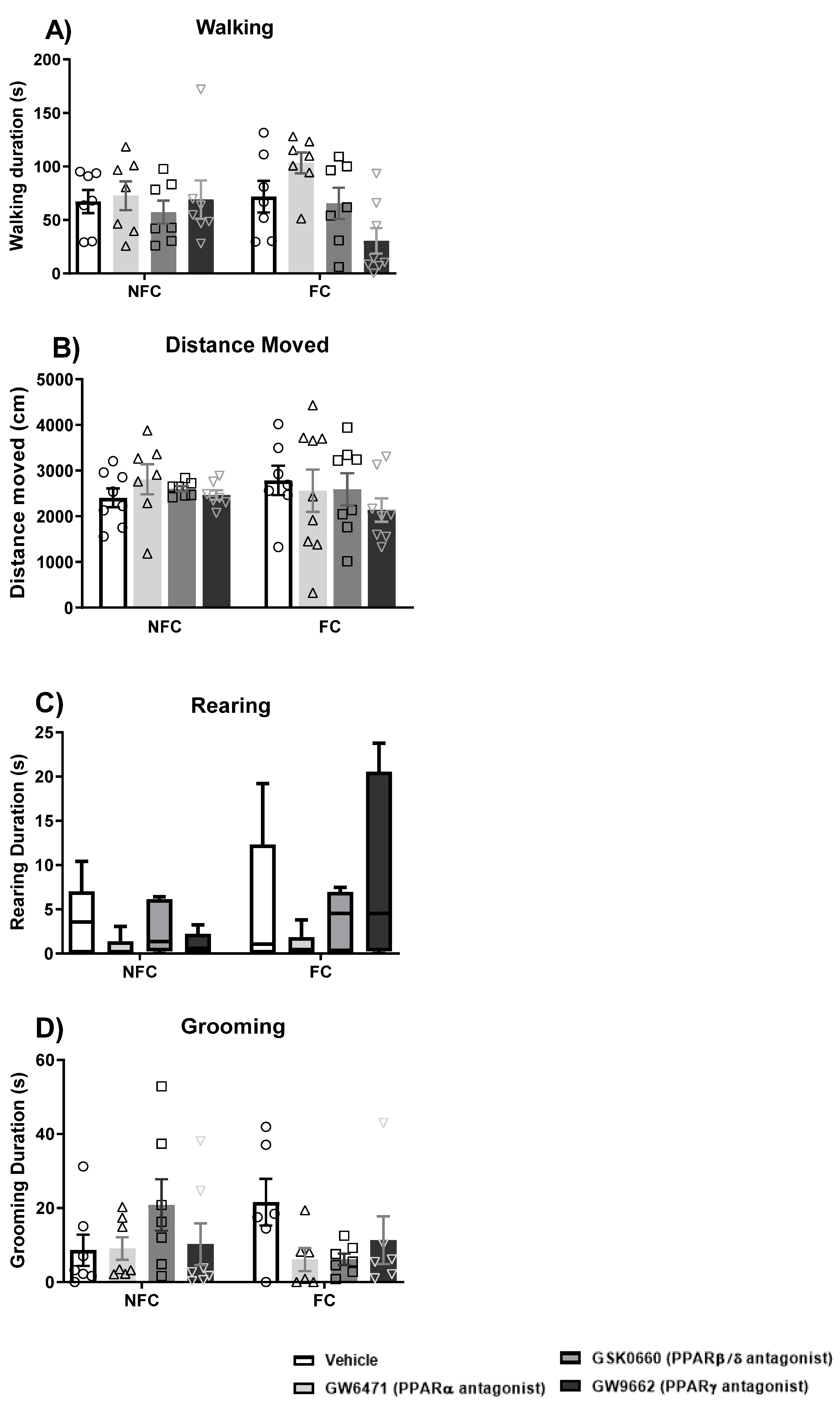
2.1.3. Intra-BLA Administration of GW6471, GSK0660, and GW9662 Does Not Affect General/Motor Behaviour
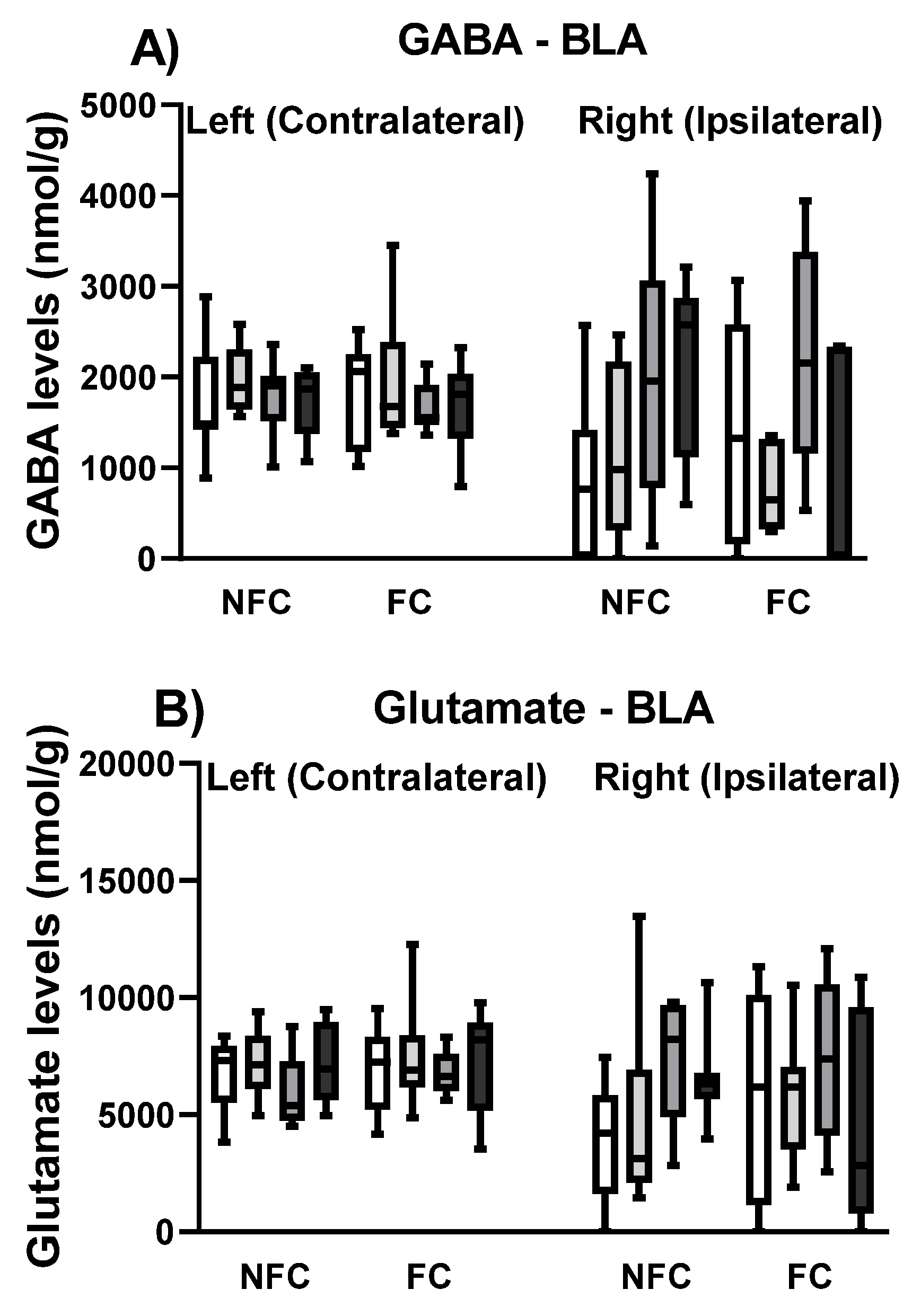
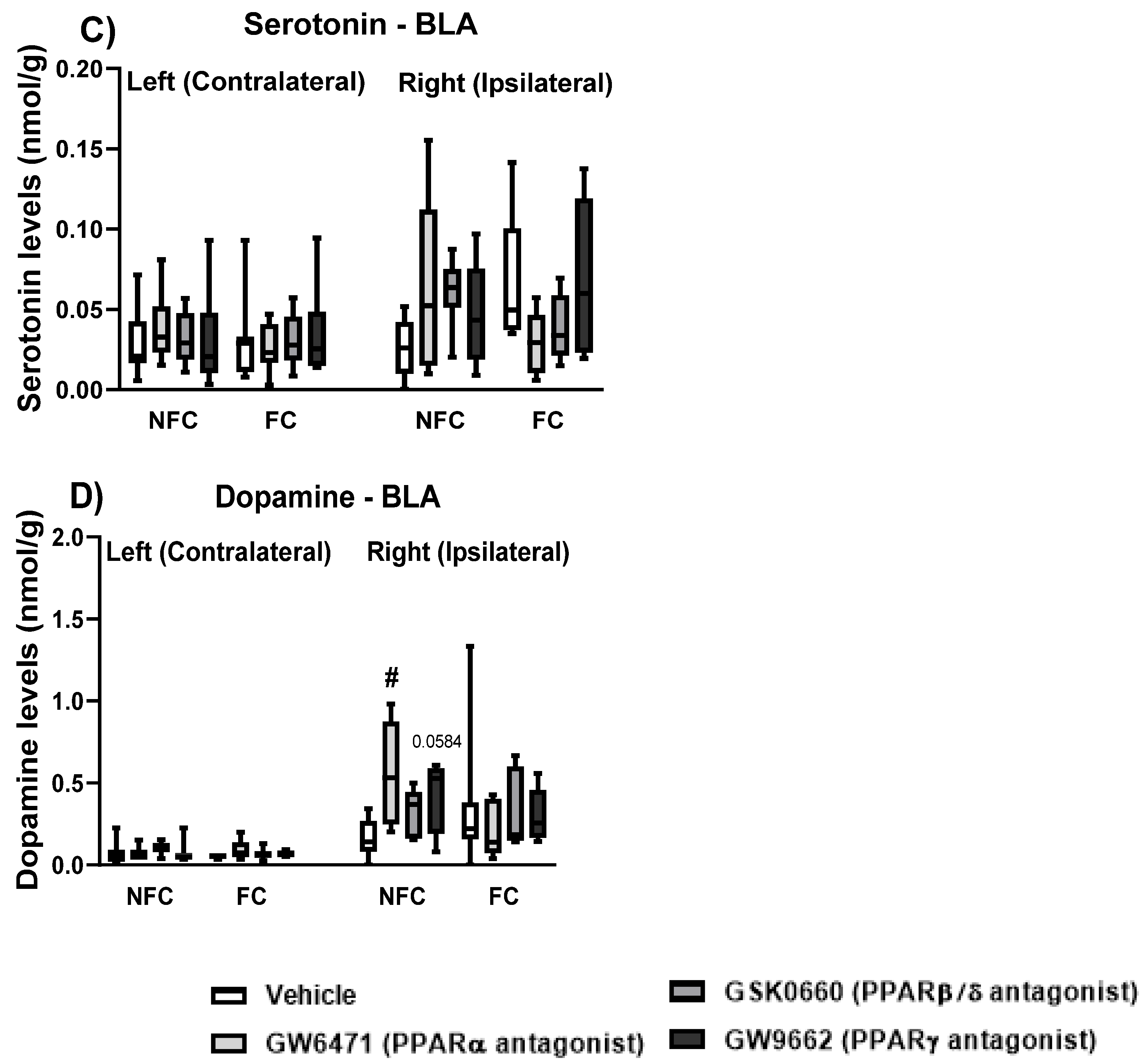
2.1.4. Effect of Fear Conditioning and PPAR Antagonist Administration on Neurotransmitter Levels in the Basolateral Amygdala (BLA)
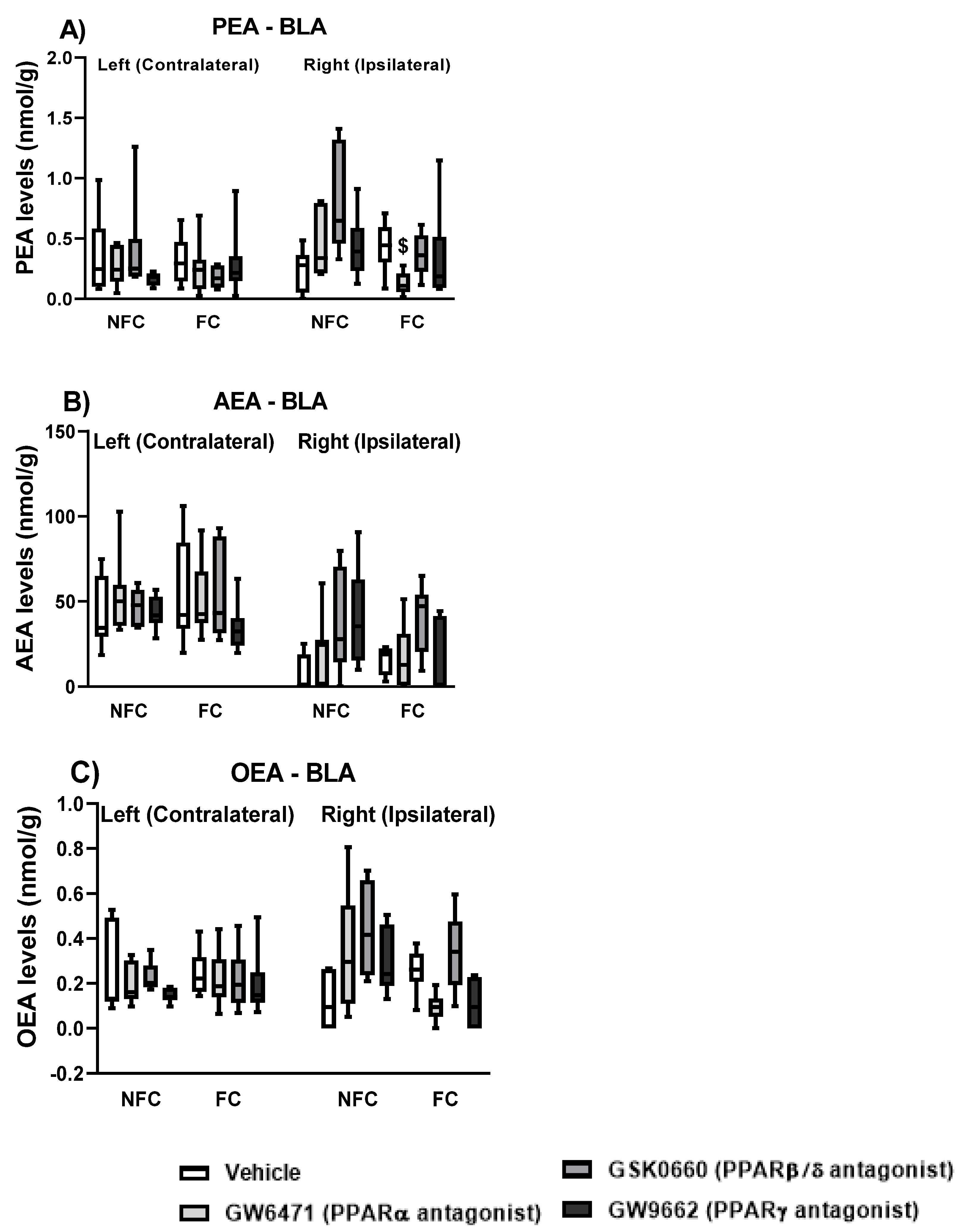
2.1.5. Effect of Fear Conditioning and PPAR Antagonist Administration on Endocannabinoids and NAE Levels in the Basolateral Amygdala (BLA)
2.2. Experiment 2: Effects of Intra-BLA Administration of PPAR Antagonists on Conditioned Fear in the Absence of Nociceptive Tone in Rats
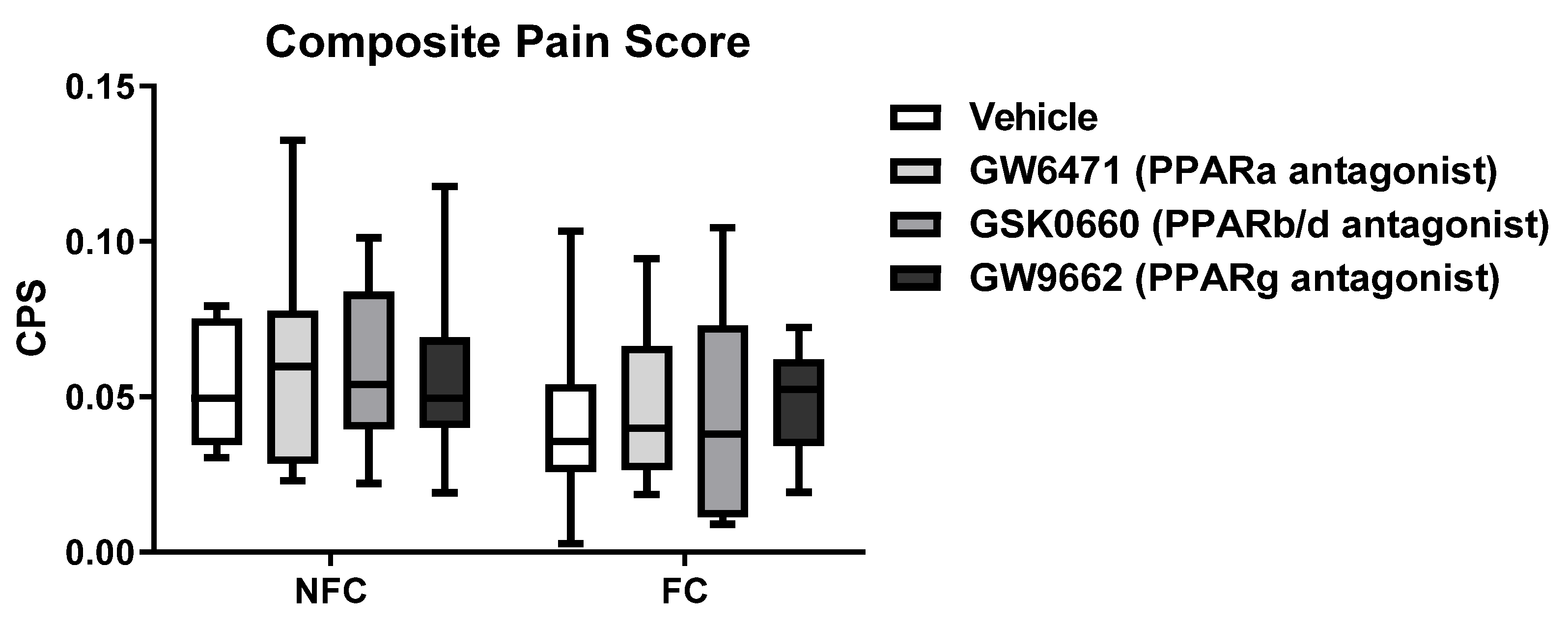
2.2.1. Intra-BLA Administration of PPAR Antagonists Had No Effect on Composite Pain Score in Saline-Injected Rats
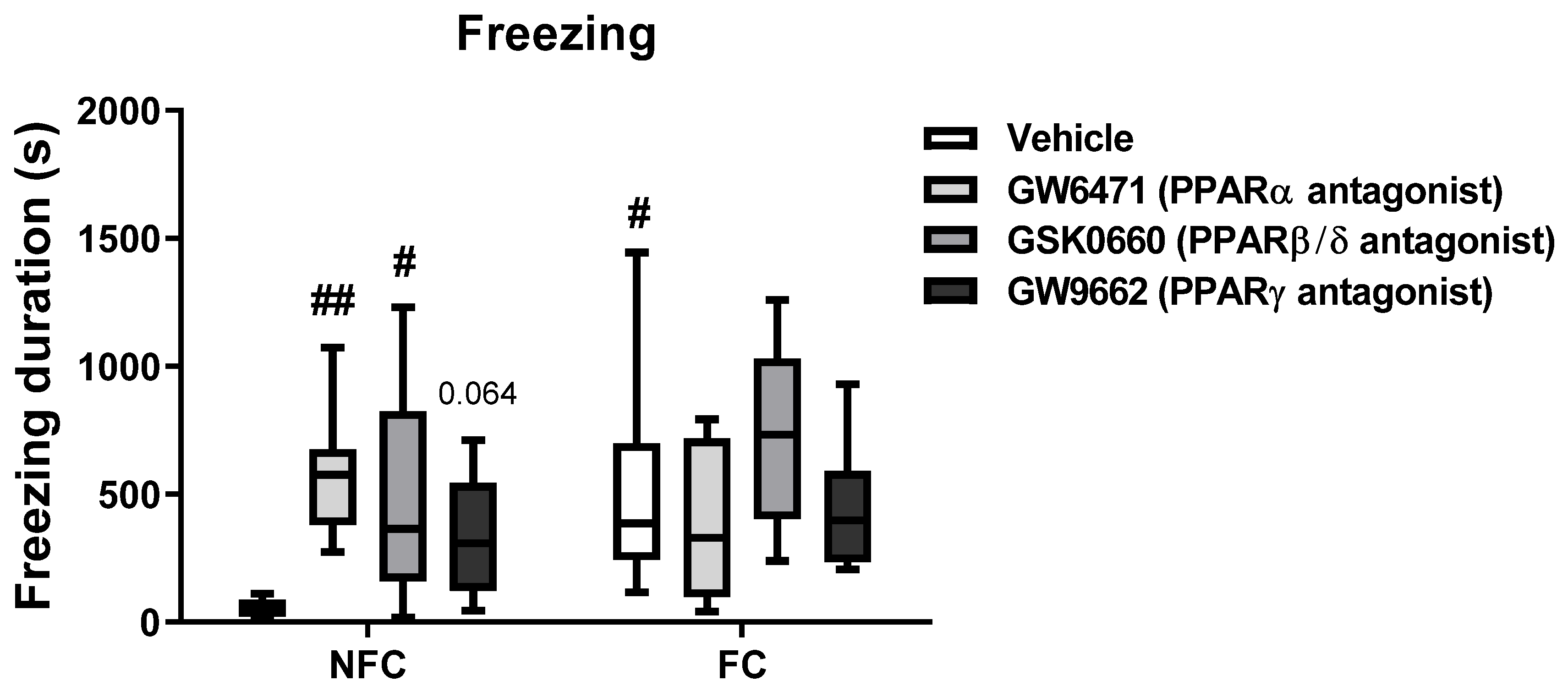
2.2.2. Intra-BLA Administration of PPARs Antagonists Increases Freezing in NFC Rats
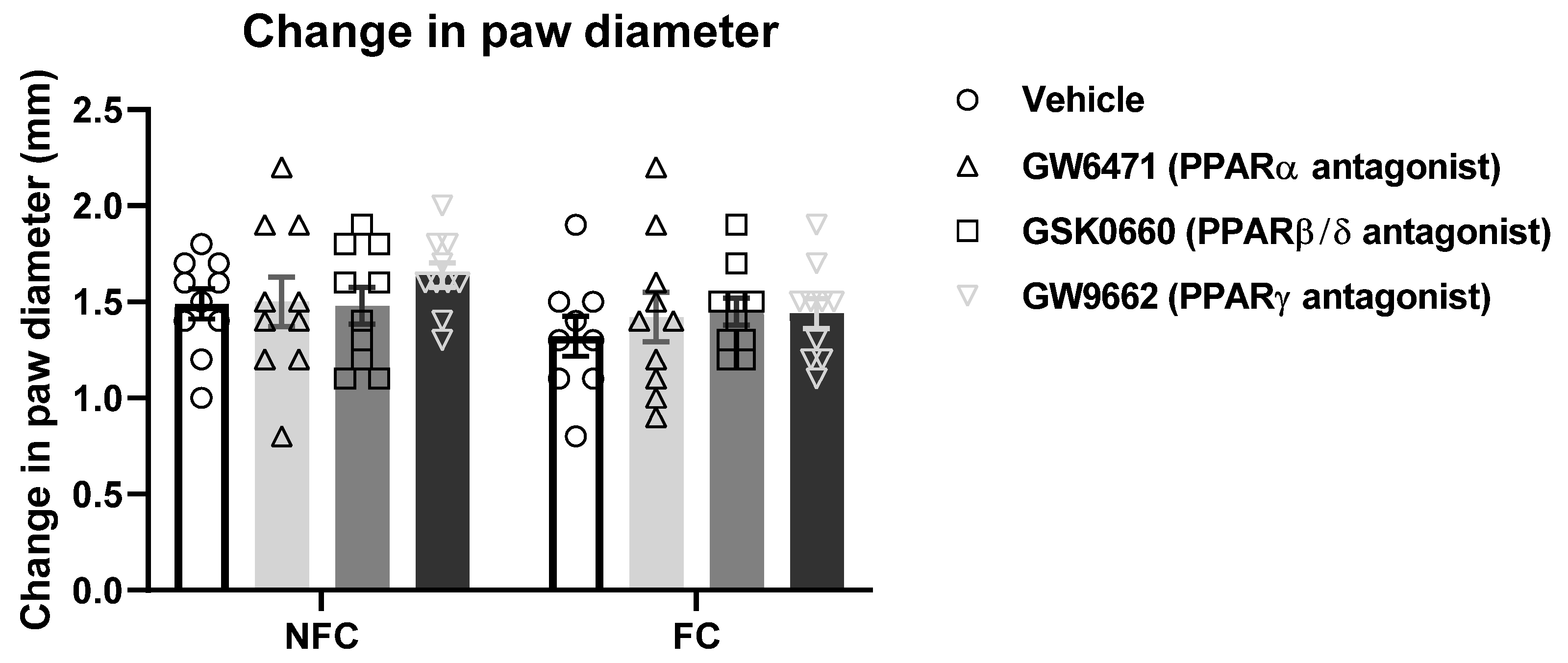
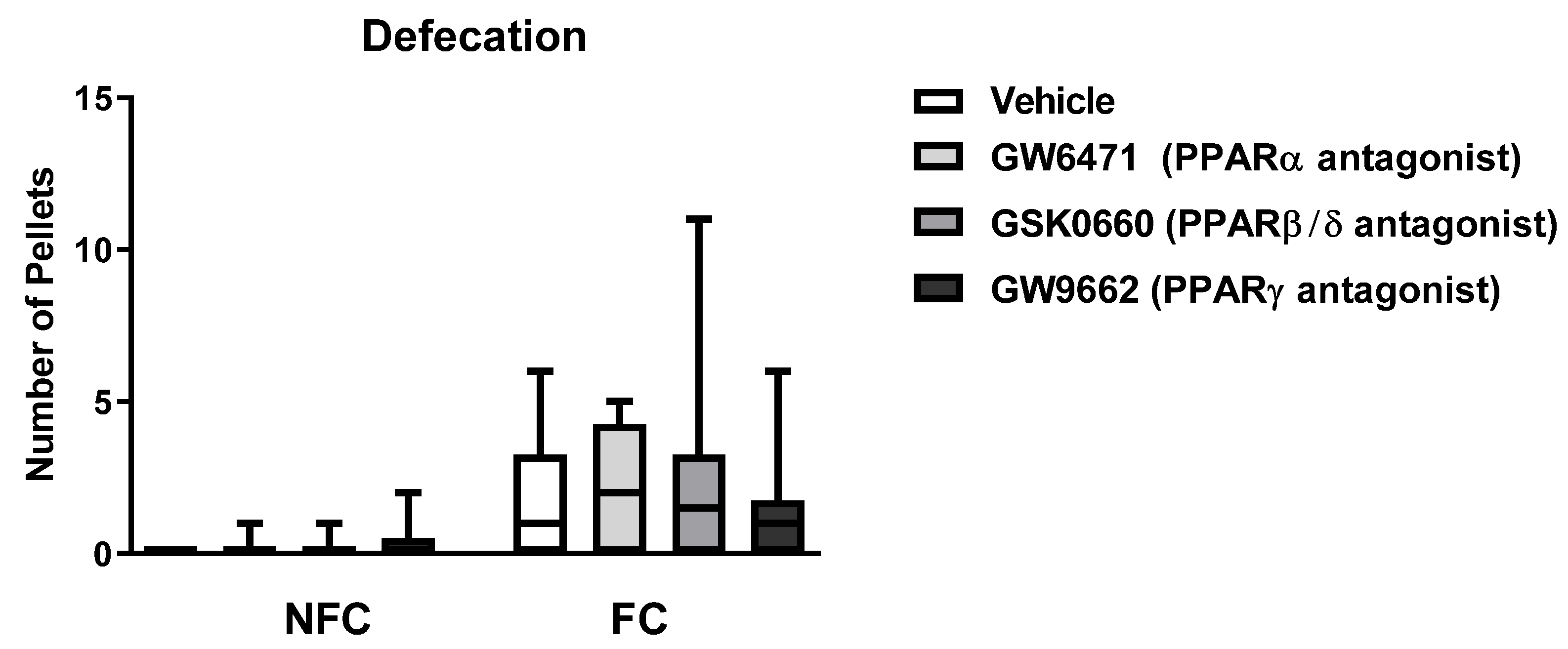
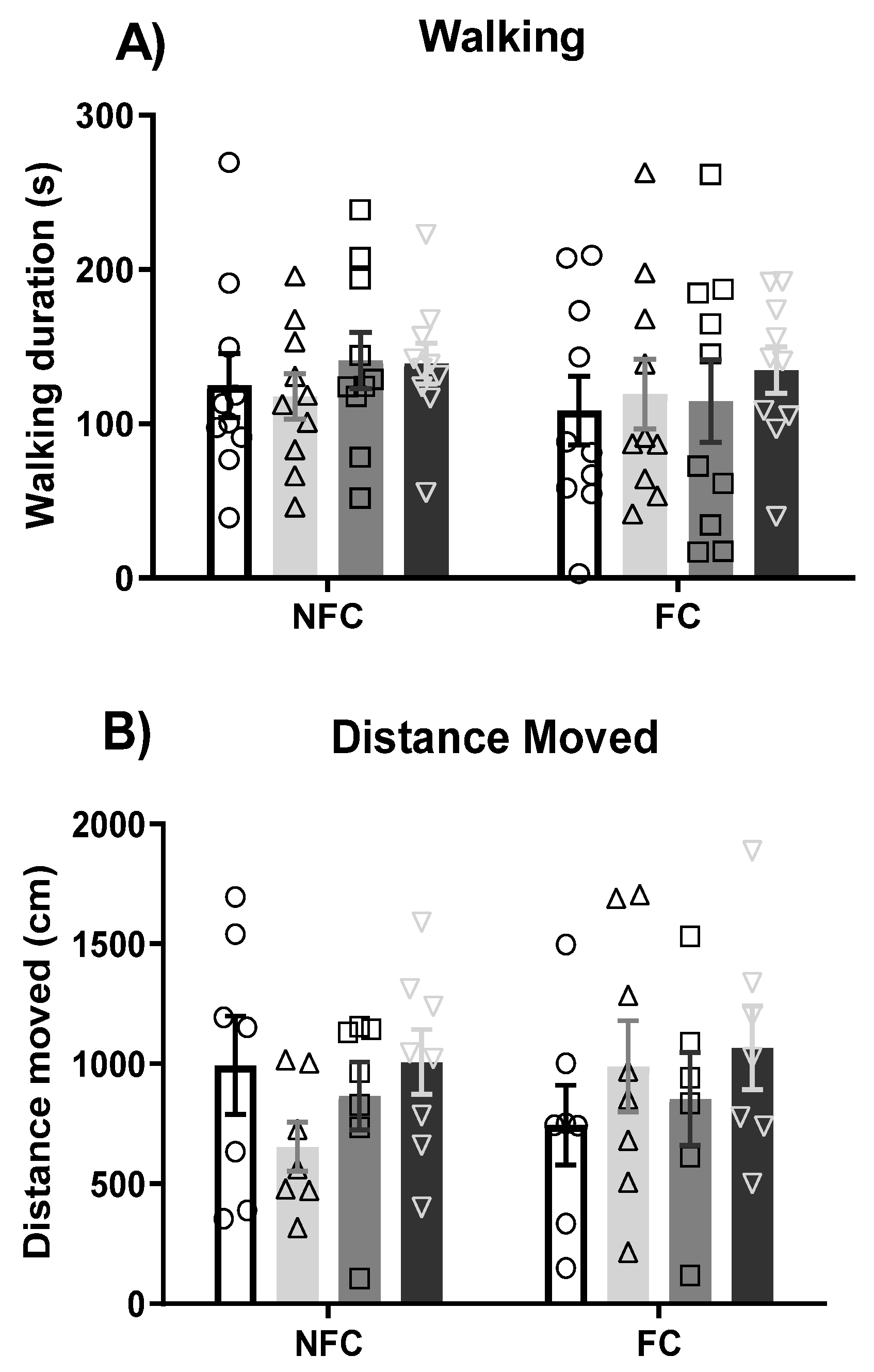
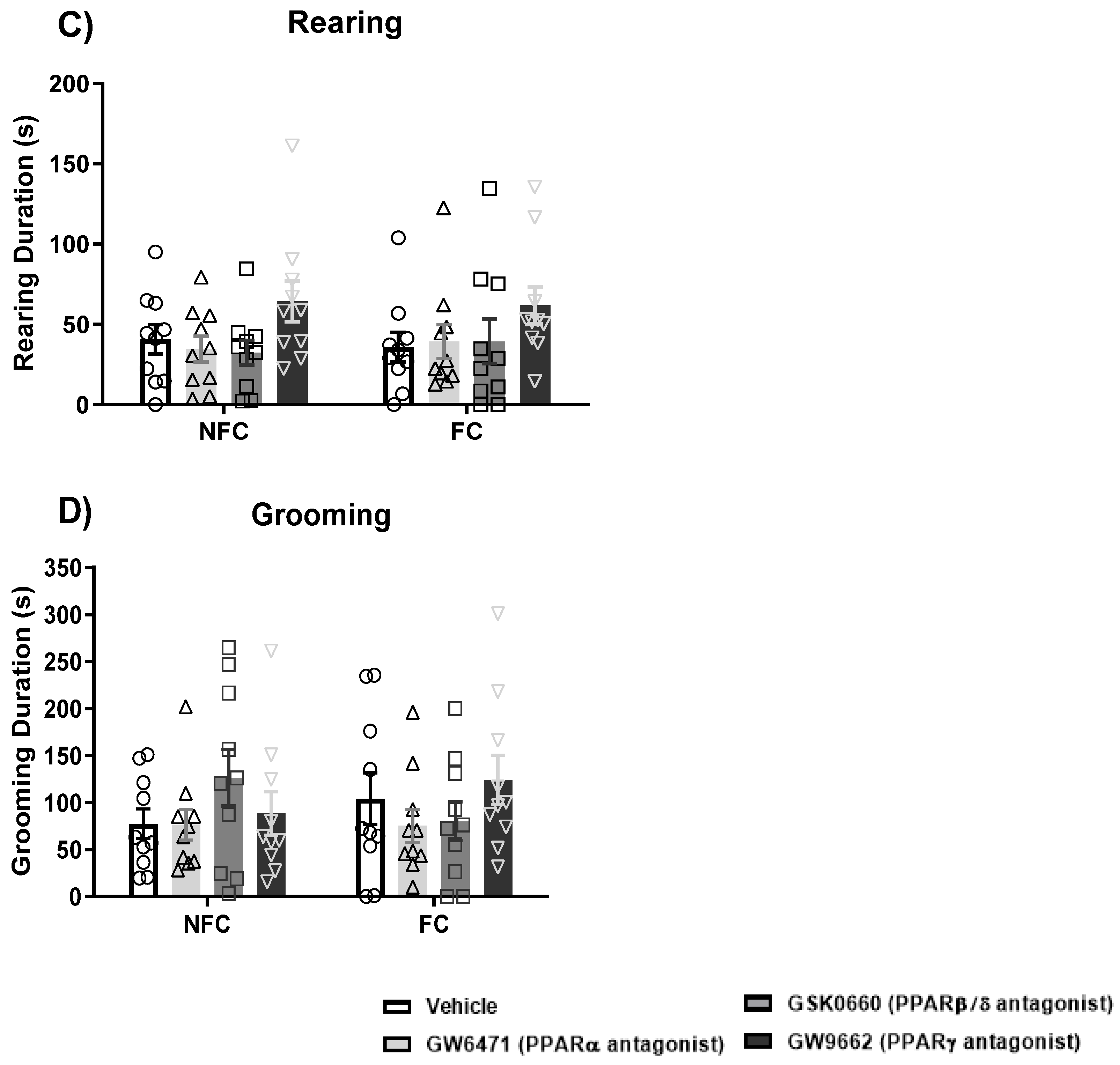
2.2.3. Intra-BLA Administration of PPAR Antagonists Does Not Affect General/Motor Behaviour
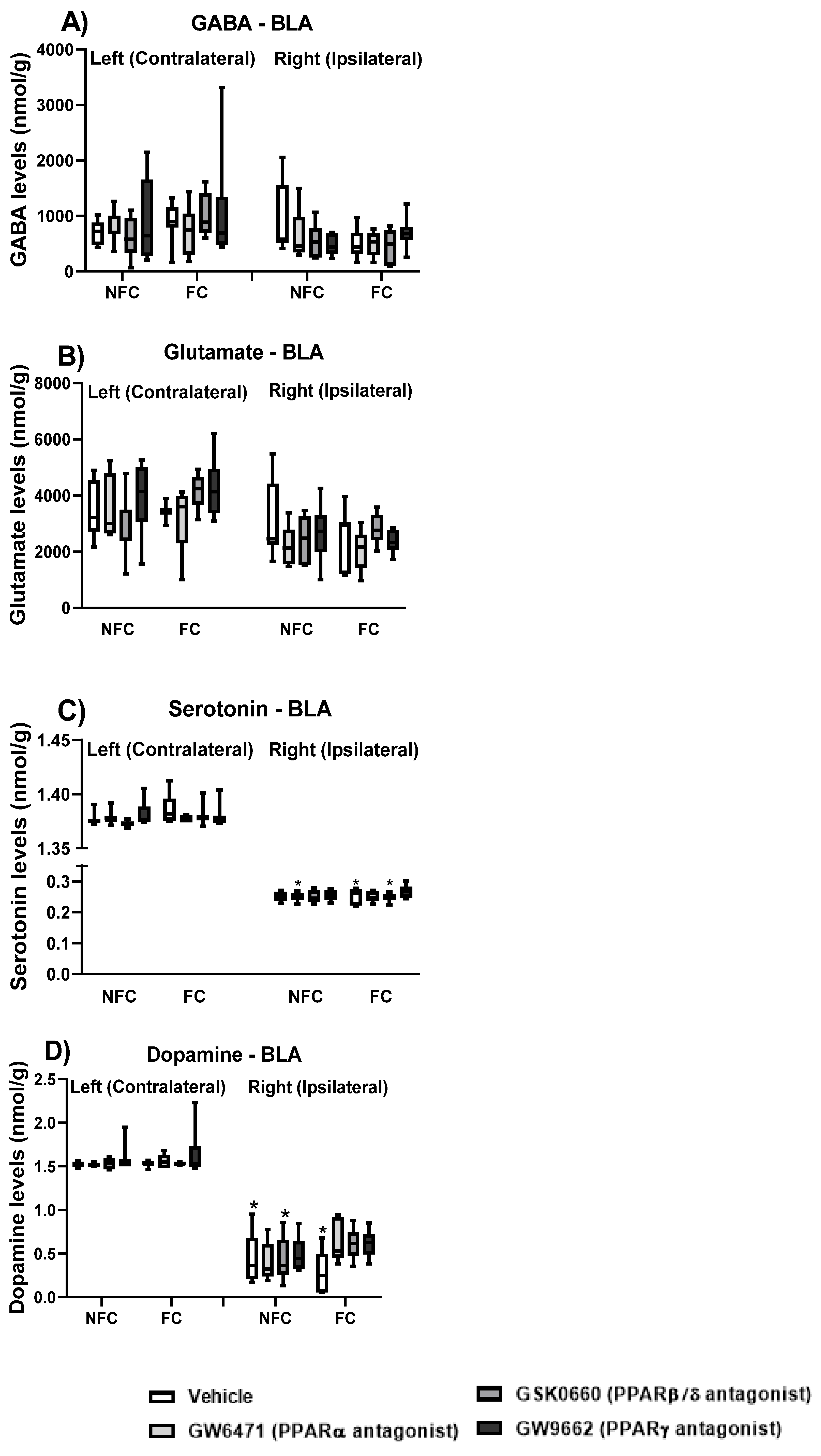
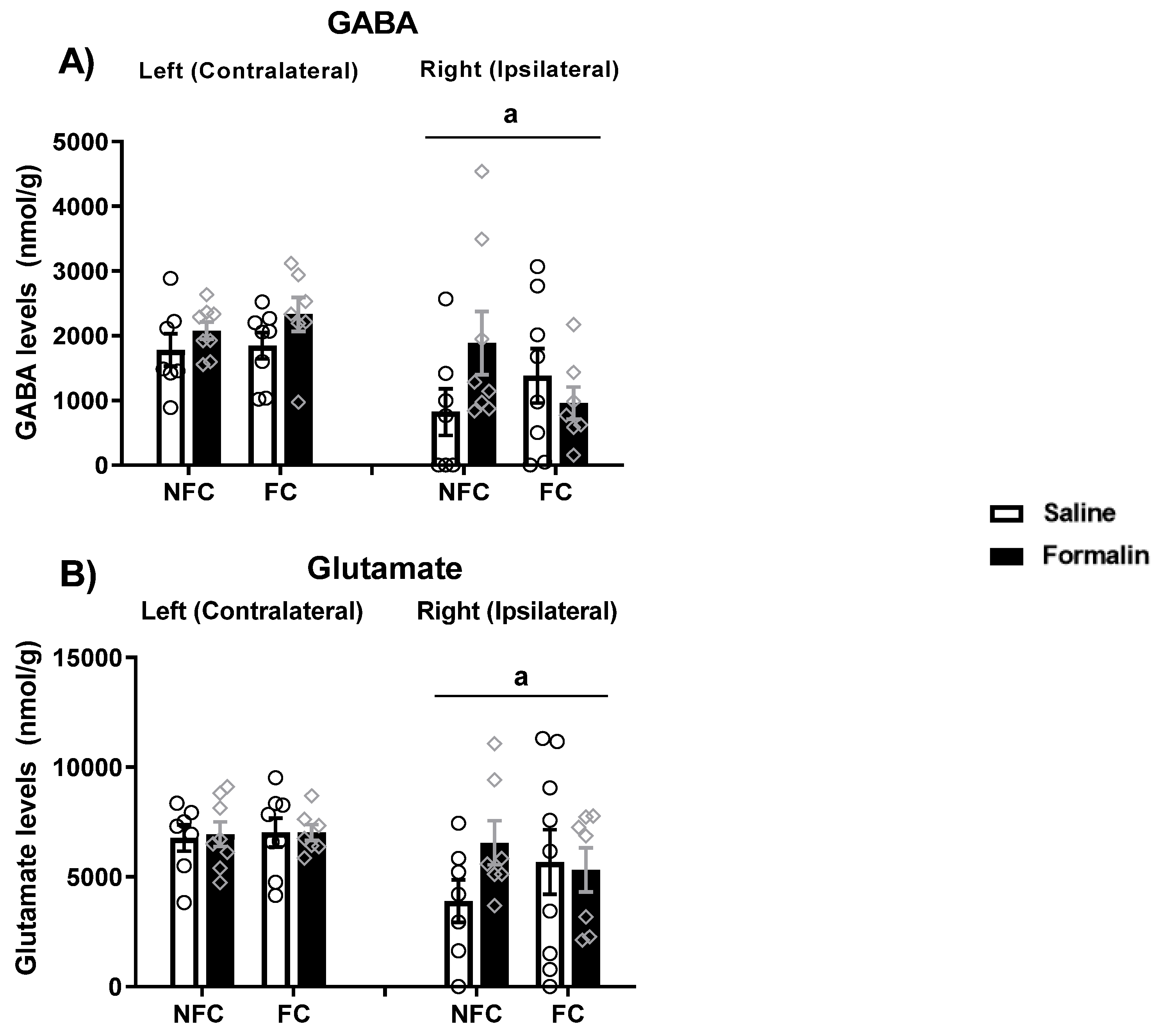
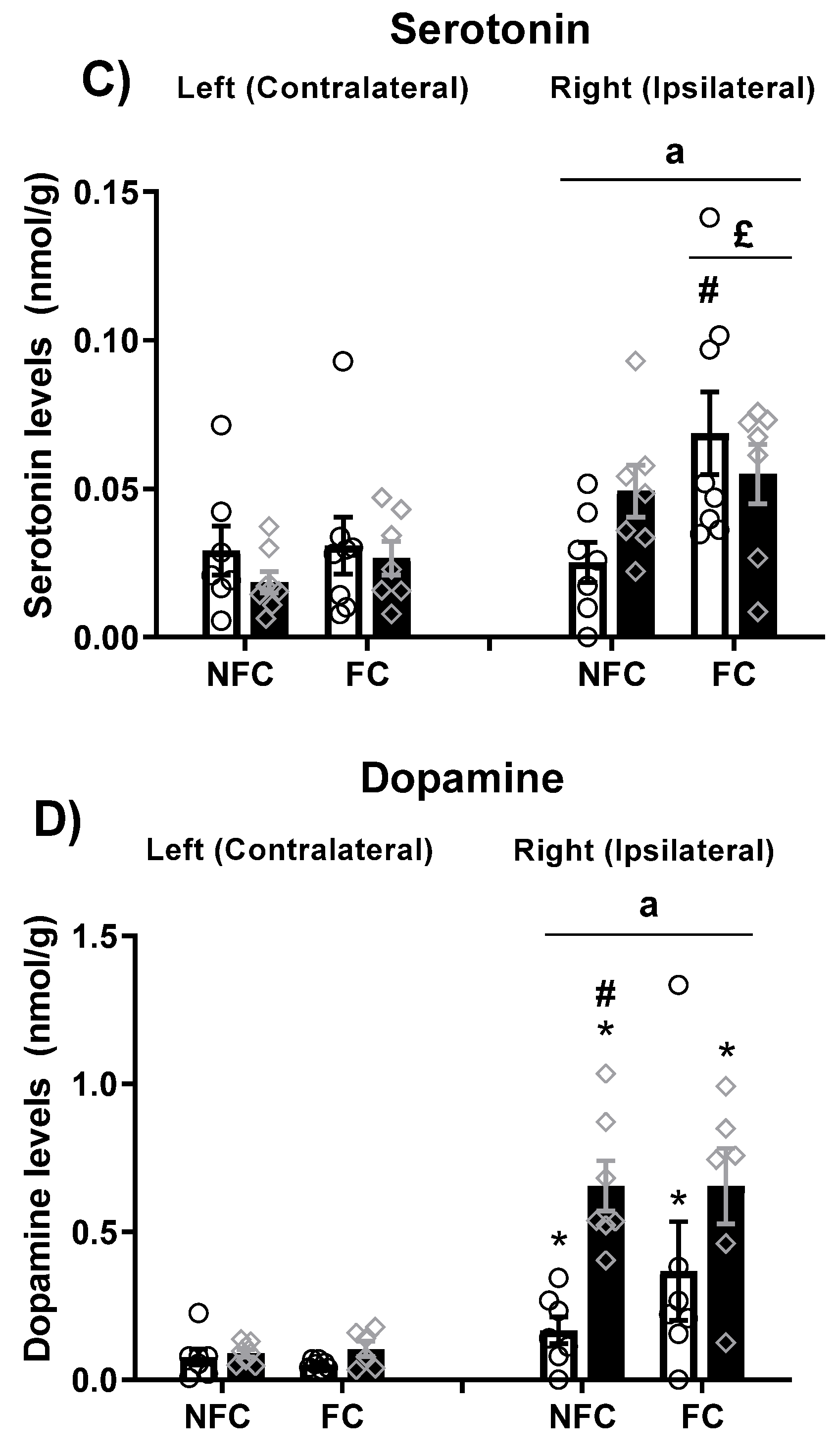
2.2.4. Effect of Fear Conditioning and PPAR Antagonist Administration on Neurotransmitter Levels in the Basolateral Amygdala (BLA)
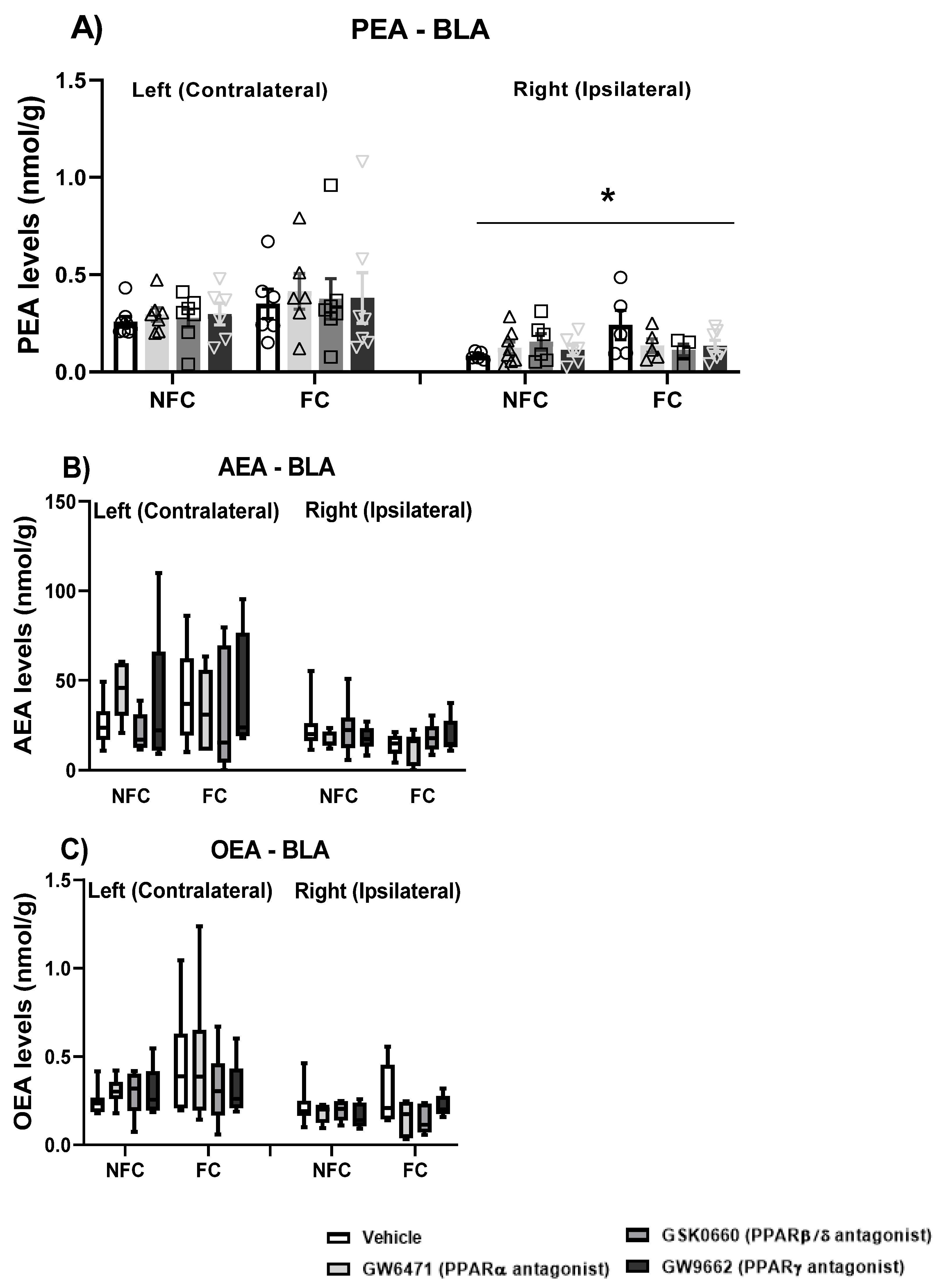
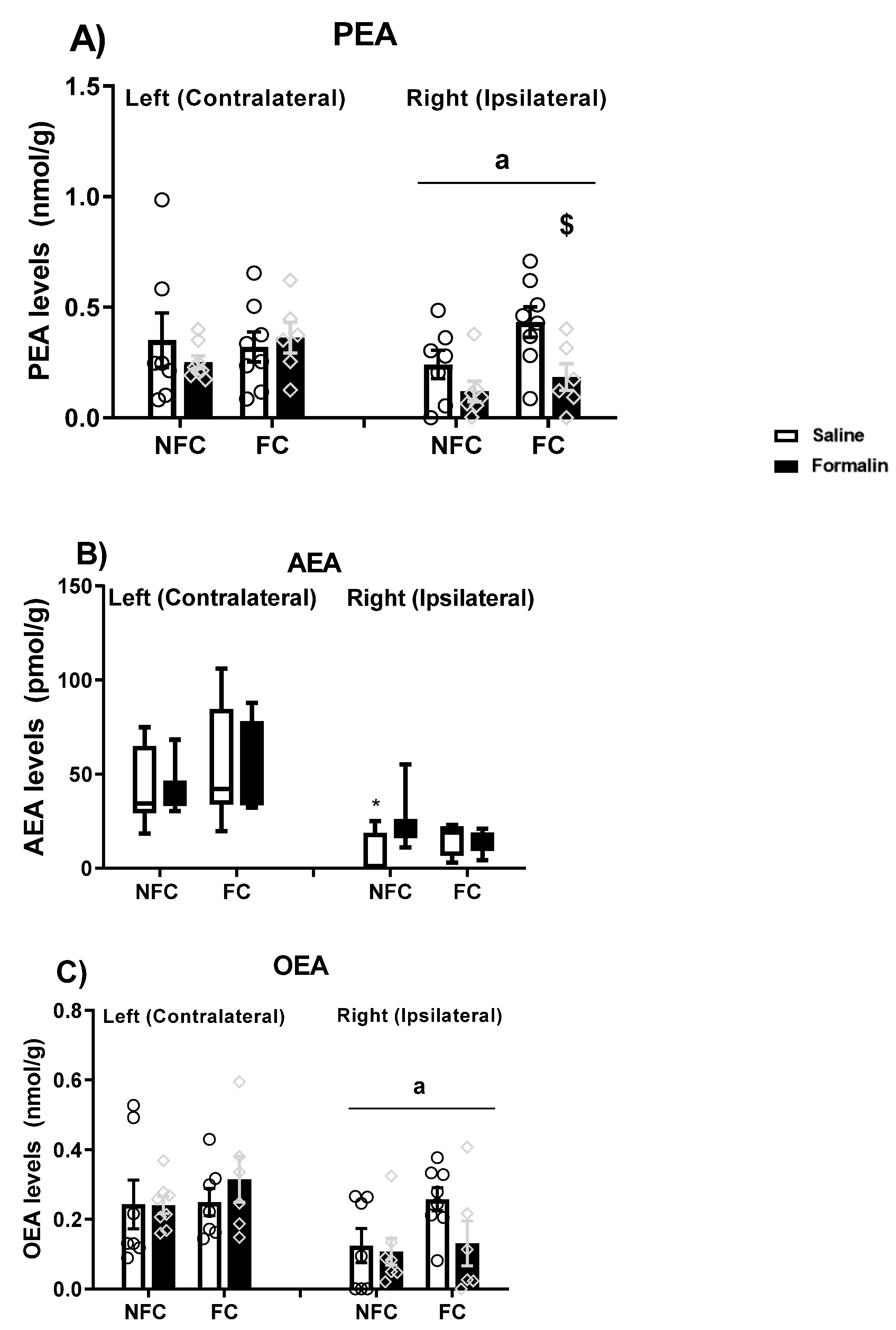
2.2.5. Effect of Fear Conditioning and PPAR Antagonist Administration on Endocannabinoid and NAE Levels in the Basolateral Amygdala (BLA)
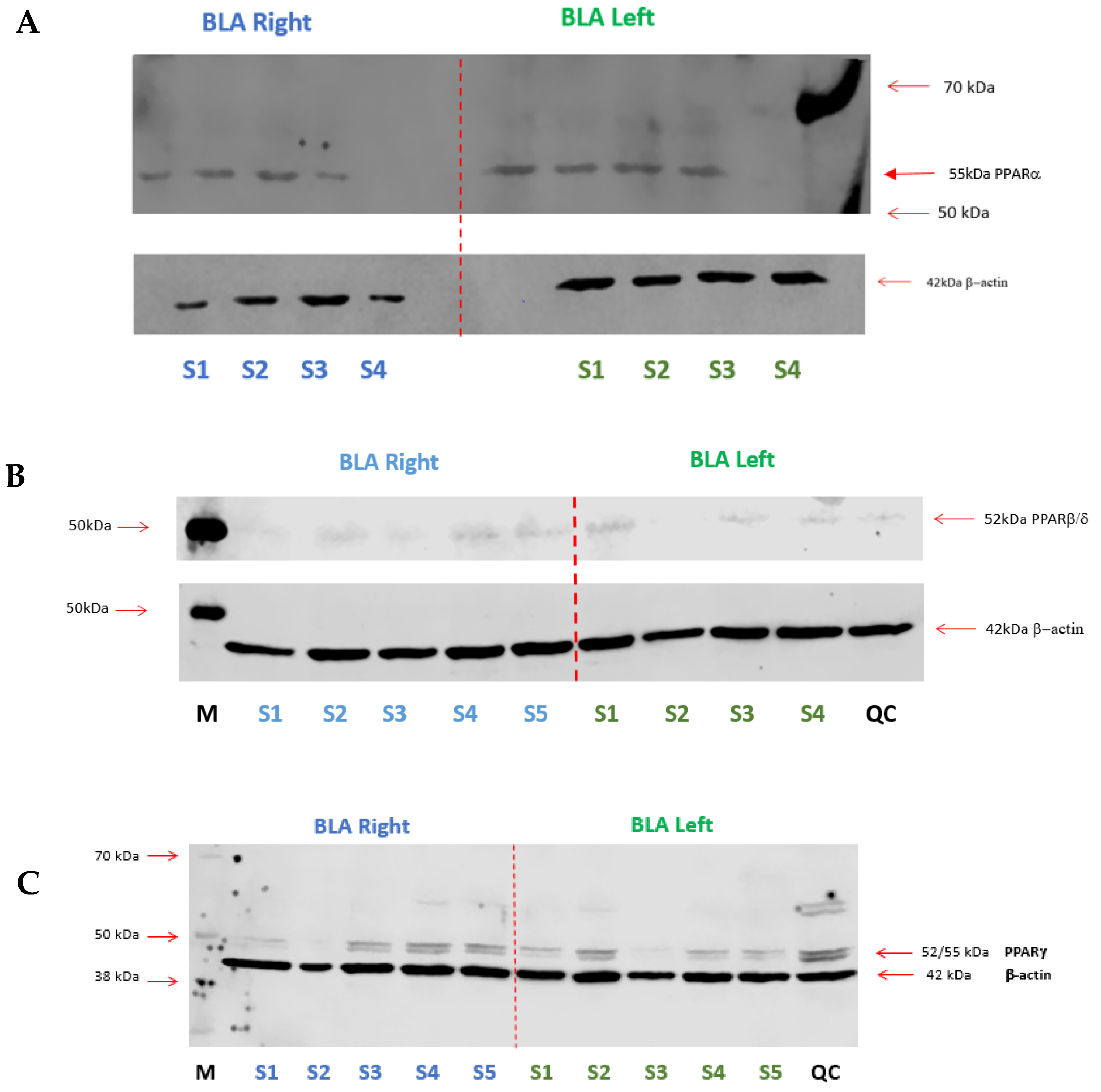
2.3. Expression of PPARs in the BLA
2.3.1. Western Blotting
2.3.2. RT-qPCR
2.4. Effects of Intraplantar Administration of Formalin on Levels of Neurotransmitters, Endocannabinoids, and NAEs in NFC and FC Rats
2.4.1. Effects of Intraplantar (i.pl.) Administration of Formalin on Levels of Neurotransmitters in NFC and FC Rats
2.4.2. Effects of Intraplantar Administration of Formalin on Levels of Endocannabinoids and NAEs in NFC and FC Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cannula Implantation
4.3. Drugs
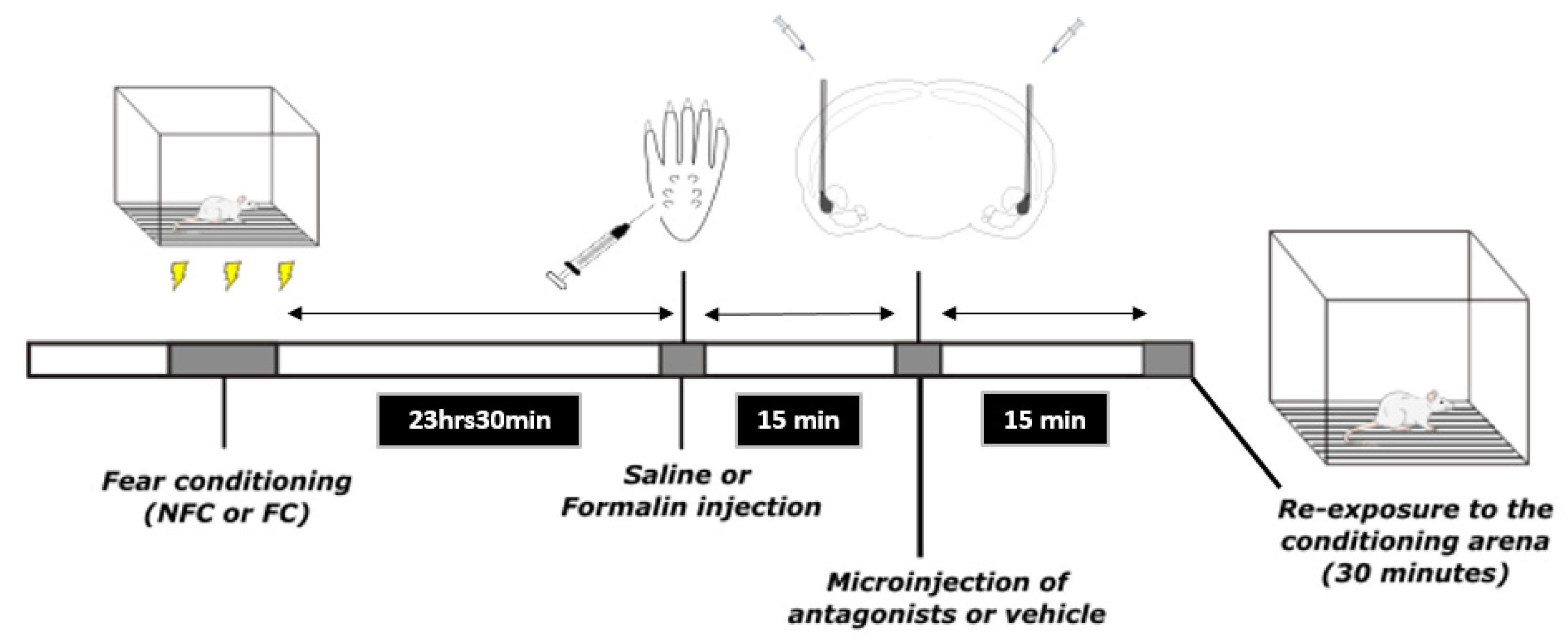
4.4. Experimental Procedure
4.5. Behavioural Analysis
4.6. Histological Verification of Intracerebral Injection Sites
4.7. Cryo-Sectioning and Tissue Microdissection
4.8. Measurement of Endocannabinoids, NAEs, and Neurotransmitters in Discrete Brain Regions Using Liquid Chromatography—Tandem Mass Spectrometry (LC-MS/MS)
4.9. Verification of PPAR Expression in the BLA
4.9.1. Verification of PPAR Expression in the BLA by Western Blotting
4.9.2. Verification of PPARβ/δ Expression in the BLA by RT-qPCR
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Issemann, I.; Green, S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature 1990, 347, 645–650. [Google Scholar] [CrossRef]
- Marion-Letellier, R.; Savoye, G.; Ghosh, S. Fatty acids, eicosanoids and PPAR gamma. Eur. J. Pharmacol. 2016, 785, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Waku, T.; Shiraki, T.; Oyama, T.; Maebara, K.; Nakamori, R.; Morikawa, K. The nuclear receptor PPARγ individually responds to serotonin- and fatty acid-metabolites. EMBO J. 2010, 29, 3395–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouaboula, M.; Hilairet, S.; Marchand, J.; Fajas, L.; Le Fur, G.; Casellas, P. Anandamide induced PPARγ transcriptional activation and 3T3-L1 preadipocyte differentiation. Eur. J. Pharmacol. 2005, 517, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, C.E.; Snider, N.T.; Thompson, J.T.; Vanden Heuvel, J.P.; Kaminski, N.E. Interleukin-2 suppression by 2-arachidonyl glycerol is mediated through peroxisome proliferator activated receptor independently of cannabinoid receptors 1 and 2. Mol. Pharmacol. 2006, 70, 101–111. [Google Scholar] [CrossRef] [PubMed]
- LoVerme, J.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The search for the palmitoylethanolamide receptor. Life Sci. 2005, 77, 1685–1698. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Gaetani, S.; Oveisi, F.; Lo Verme, J.; Serrano, A.; Rodríguez De Fonseca, F.; Rosengarth, A.; Luecke, H.; Di Giacomo, B.; Tarzia, G.; et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-a. Nature 2003, 425, 90–93. [Google Scholar] [CrossRef]
- Hong, F.; Xu, P.; Zhai, Y. The Opportunities and Challenges of Peroxisome Proliferator-Activated Receptors Ligands in Clinical Drug Discovery and Development. Int. J. Mol. Sci. 2018, 19, 2189. [Google Scholar] [CrossRef] [Green Version]
- Fruchart, J.C.; Staels, B.; Duriez, P. The role of fibric acids in atherosclerosis. Curr. Atheroscler. Rep. 2001, 3, 83–92. [Google Scholar] [CrossRef]
- Davis, M. The role of the amygdala in fear and anxiety. Annu. Rev. Neurosci. 1992, 15, 353–375. [Google Scholar] [CrossRef]
- Sah, P.; Faber, E.S.L.; Lopez De Armentia, M.; Power, J. The Amygdaloid Complex: Anatomy and Physiology. Physiol. Rev. 2003, 83, 803–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsvetkov, E.A.; Krasnoshchekova, E.I.; Vesselkin, N.P.; Kharazova, A.D. Amygdala: Neuroanatomy and Neurophysiology of Fear. J. Evol. Biochem. Physiol. 2015, 51, 406–418. [Google Scholar] [CrossRef]
- LeDoux, J. The amygdala. Curr. Biol. 2007, 17, R868–R874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, J.W.; Han, J.-S.; Kim, J.J. Selective Neurotoxic Lesions of Basolateral and Central Nuclei of the Amygdala Produce Differential Effects on Fear Conditioning. J. Neurosci. 2004, 24, 7654–7662. [Google Scholar] [CrossRef] [Green Version]
- Maren, S. Neurotoxic Basolateral Amygdala Lesions Impair Learning and Memory But Not the Performance of Conditional Fear in Rats. J. Neurosci. 1999, 19, 8696–8703. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.G.; LeDoux, J.E. Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav. Neurosci. 1992, 106, 274–285. [Google Scholar] [CrossRef]
- Sananes, C.B.; Davis, M. N-methyl-D-aspartate lesions of the lateral and basolateral nuclei of the amygdala block fear-potentiated startle and shock sensitization of startle. Behav. Neurosci. 1992, 106, 72–80. [Google Scholar] [CrossRef]
- Helmstetter, F.J. Contribution of the amygdala to learning and performance of conditional fear. Physiol. Behav. 1992, 51, 1271–1276. [Google Scholar] [CrossRef]
- Kim, J.J.; Rison, R.A.; Fanselow, M.S. Effects of amygdala, hippocampus, and periaqueductal gray lesions on short- and long-term contextual fear. Behav. Neurosci. 1993, 107, 1093–1098. [Google Scholar] [CrossRef]
- Muller, J.; Corodimas, K.P.; Fridel, Z.; LeDoux, J.E. Functional inactivation of the lateral and basal nuclei of the amygdala by muscimol infusion prevents fear conditioning to an explicit conditioned stimulus and to contextual stimuli. Behav. Neurosci. 1997, 111, 683–691. [Google Scholar] [CrossRef]
- Sacchetti, B.; Lorenzini, C.A.; Baldi, E.; Tassoni, G.; Bucherelli, C. Auditory thalamus, dorsal hippocampus, basolateral amygdala, and perirhinal cortex role in the consolidation of conditioned freezing to context and to acoustic conditioned stimulus in the rat. J. Neurosci. 1999, 19, 9570–9578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miserendino, M.J.D.; Sananes, C.B.; Melia, K.R.; Davis, M. Blocking of acquisition but not expression of conditioned fear-potentiated startle by NMDA antagonists in the amygdala. Nature 1990, 345, 716–718. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, M.S.; Kim, J.J. Acquisition of contextual Pavlovian fear conditioning is blocked by application of an NMDA receptor antagonist D,L-2-amino-5-phosphonovaleric acid to the basolateral amygdala. Behav. Neurosci. 1994, 108, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Helmstetter, F.J.; Bellgowan, P.S. Effects of muscimol applied to the basolateral amygdala on acquisition and expression of contextual fear conditioning in rats. Behav. Neurosci. 1994, 108, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Baldi, E.; Bucherelli, C. Substantia nigra, nucleus basalis magnocellularis and basolateral amygdala roles in extinction of contextual fear conditioning in the rat. Neurobiol. Learn. Mem. 2010, 94, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, H.; Radahmadi, M.; Reisi, P.; Alaei, H. Effects of electrical lesion of basolateral amygdala nucleus on rat anxiety-like behaviour under acute, sub-chronic, and chronic stresses. Clin. Exp. Pharmacol. Physiol. 2017, 44, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Degoulet, M.F.; Rostain, J.C.; David, H.N.; Abraini, J.H. Repeated administration of amphetamine induces a shift of the prefrontal cortex and basolateral amygdala motor function. Int. J. Neuropsychopharmacol. 2009, 12, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Makkar, S.R.; Zhang, S.Q.; Cranney, J.; Cranney, J. Behavioral and Neural Analysis of GABA in the Acquisition, Consolidation, Reconsolidation, and Extinction of Fear Memory. Neuropsychopharmacology 2010, 35, 1625–1652. [Google Scholar] [CrossRef]
- Walker, D.L.; Davis, M. The role of amygdala glutamate receptors in fear learning, fear-potentiated startle, and extinction. Pharmacol. Biochem. Behav. 2002, 71, 379–392. [Google Scholar] [CrossRef]
- Davis, M.; Myers, K.M. The role of glutamate and gamma-aminobutyric acid in fear extinction: Clinical implications for exposure therapy. Biol. Psychiatry 2002, 52, 998–1007. [Google Scholar] [CrossRef]
- Bauer, E.P. Serotonin in fear conditioning processes. Behav. Brain Res. 2015, 277, 68–77. [Google Scholar] [CrossRef]
- Fadok, J.P.; Darvas, M.; Dickerson, T.M.K.; Palmiter, R.D. Long-Term Memory for Pavlovian Fear Conditioning Requires Dopamine in the Nucleus Accumbens and Basolateral Amygdala. PLoS ONE 2010, 5, e12751. [Google Scholar] [CrossRef] [PubMed]
- Chhatwal, J.P.; Ressler, K.J. Modulation of fear and anxiety by the endogenous cannabinoid system. CNS Spectr. 2007, 12, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Sun, H.; Fu, Y.; Li, Z.; Pais-Vieira, M.; Galhardo, V.; Neugebauer, V. Cognitive Impairment in Pain through Amygdala-Driven Prefrontal Cortical Deactivation. J. Neurosci. 2010, 30, 5451–5464. [Google Scholar] [CrossRef] [PubMed]
- Luongo, L.; de Novellis, V.; Gatta, L.; Palazzo, E.; Vita, D.; Guida, F.; Giordano, C.; Siniscalco, D.; Marabese, I.; De Chiaro, M.; et al. Role of metabotropic glutamate receptor 1 in the basolateral amygdala-driven prefrontal cortical deactivation in inflammatory pain in the rat. Neuropharmacology 2013, 66, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.K.; Ehling, S.; Barbar, M.; Thomas, J.; Hughes, M.A.; Smith, C.E.; Pogorelov, V.M.; Aryal, D.K.; Wetsel, W.C.; Lascelles, B.D.X. Distinct neuronal populations in the basolateral and central amygdala are activated with acute pain, conditioned fear, and fear-conditioned analgesia. Neurosci. Lett. 2017, 661, 11–17. [Google Scholar] [CrossRef]
- Nakagawa, T.; Katsuya, A.; Tanimoto, S.; Yamamoto, J.; Yamauchi, Y.; Minami, M.; Satoh, M. Differential patterns of c-fos mRNA expression in the amygdaloid nuclei induced by chemical somatic and visceral noxious stimuli in rats. Neurosci. Lett. 2003, 344, 197–200. [Google Scholar] [CrossRef]
- Asmundson, G.J.; Coons, M.J.; Taylor, S.; Katz, J. PTSD and the Experience of Pain: Research and Clinical Implications of Shared Vulnerability and Mutual Maintenance Models. Can. J. Psychiatry 2002, 47, 930–937. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.G.; França, M.H.; de Paiva, M.C.A.; Andrade, L.H.; Viana, M.C. Prevalence and clinical profile of chronic pain and its association with mental disorders. Rev. Saude Publica 2017, 51, 96. [Google Scholar] [CrossRef] [Green Version]
- Warden, A.; Truitt, J.; Merriman, M.; Ponomareva, O.; Jameson, K.; Ferguson, L.B.; Mayfield, R.D.; Harris, R.A.; Berger, J.; Moller, D.E.; et al. Localization of PPAR isotypes in the adult mouse and human brain. Sci. Rep. 2016, 6, 27618. [Google Scholar] [CrossRef]
- Moreno, S.; Farioli-Vecchioli, S.; Cerù, M. Immunolocalization of peroxisome proliferator-activated receptors and retinoid × receptors in the adult rat CNS. Neuroscience 2004, 123, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Okine, B.N.; Rea, K.; Olango, W.M.; Price, J.; Herdman, S.; Madasu, M.K.; Roche, M.; Finn, D.P. A role for PPARα in the medial prefrontal cortex in formalin-evoked nociceptive responding in rats. Br. J. Pharmacol. 2014, 171, 1462–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domi, E.; Uhrig, S.; Soverchia, L.; Spanagel, R.; Hansson, A.C.; Barbier, E.; Heilig, M.; Ciccocioppo, R.; Ubaldi, M. Genetic Deletion of Neuronal PPARγ Enhances the Emotional Response to Acute Stress and Exacerbates Anxiety: An Effect Reversed by Rescue of Amygdala PPARγ Function. J. Neurosci. 2016, 36, 12611–12623. [Google Scholar] [CrossRef] [PubMed]
- Okine, B.N.; Gaspar, J.C.; Madasu, M.K.; Olango, W.M.; Harhen, B.; Roche, M.; Finn, D.P. Characterisation of peroxisome proliferator-activated receptor signalling in the midbrain periaqueductal grey of rats genetically prone to heightened stress, negative affect and hyperalgesia. Brain Res. 2017, 1657, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Okine, B.N.; Gaspar, J.C.; Finn, D.P. PPARs and pain. Br. J. Pharmacol. 2018, 176, 1421–1442. [Google Scholar] [CrossRef]
- Youssef, D.A.; El-Fayoumi, H.M.; Mahmoud, M.F. Beta-caryophyllene alleviates diet-induced neurobehavioral changes in rats: The role of CB2 and PPAR-γ receptors. Biomed. Pharmacother. 2019, 110, 145–154. [Google Scholar] [CrossRef]
- Gaspar, J.C.; Okine, B.N.; Llorente-Berzal, A.; Roche, M.; Finn, D.P. Pharmacological blockade of PPAR isoforms increases conditioned fear responding in the presence of nociceptive tone. Molecules 2020, 25, 1007. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, C.; Medalie, J.; Scherma, M.; Panlilio, L.V.; Solinas, M.; Tanda, G.; Drago, F.; Cadet, J.L.; Goldberg, S.R.; Yasar, S. Fatty acid amide hydrolase (FAAH) inhibition enhances memory acquisition through activation of PPAR-alpha nuclear receptors. Learn. Mem. 2009, 16, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Campolongo, P.; Roozendaal, B.; Trezza, V.; Cuomo, V.; Astarita, G.; Fu, J.; McGaugh, J.L.; Piomelli, D. Fat-induced satiety factor oleoylethanolamide enhances memory consolidation. Proc. Natl. Acad. Sci. USA 2009, 106, 8027–8031. [Google Scholar] [CrossRef] [Green Version]
- Ratano, P.; Palmery, M.; Trezza, V.; Campolongo, P. Cannabinoid Modulation of Memory Consolidation in Rats: Beyond the Role of Cannabinoid Receptor Subtype 1. Front. Pharmacol. 2017, 8, 200. [Google Scholar] [CrossRef] [Green Version]
- Chikahisa, S.; Chida, D.; Shiuchi, T.; Harada, S.; Shimizu, N.; Otsuka, A.; Tanioka, D.; Séi, H. Enhancement of fear learning in PPARα knockout mice. Behav. Brain Res. 2019, 359, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Mijangos-Moreno, S.; Poot-Aké, A.; Guzmán, K.; Arankowsky-Sandoval, G.; Arias-Carrión, O.; Zaldívar-Rae, J.; Sarro-Ramírez, A.; Murillo-Rodríguez, E. Sleep and neurochemical modulation by the nuclear peroxisome proliferator-activated receptor α (PPAR-α) in rat. Neurosci. Res. 2016, 105, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Mascia, P.; Pistis, M.; Justinova, Z.; Panlilio, L.V.; Luchicchi, A.; Lecca, S.; Scherma, M.; Fratta, W.; Fadda, P.; Barnes, C.; et al. Blockade of Nicotine Reward and Reinstatement by Activation of Alpha-Type Peroxisome Proliferator-Activated Receptors. Biol. Psychiatry 2011, 69, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Wang, L.P.; Shen, X.; Tsien, J.Z. Balanced dopamine is critical for pattern completion during associative memory recall. PLoS ONE 2010, 5, e15401. [Google Scholar] [CrossRef] [Green Version]
- Rea, K.; Olango, W.M.; Harhen, B.; Kerr, D.M.; Galligan, R.; Fitzgerald, S.; Moore, M.; Roche, M.; Finn, D.P. Evidence for a role of GABAergic and glutamatergic signalling in the basolateral amygdala in endocannabinoid-mediated fear-conditioned analgesia in rats. Pain 2013, 154, 576–585. [Google Scholar] [CrossRef] [Green Version]
- Morena, M.; Aukema, R.J.; Leitl, K.D.; Rashid, A.J.; Vecchiarelli, H.A.; Josselyn, S.A.; Hill, M.N. Upregulation of Anandamide Hydrolysis in the Basolateral Complex of Amygdala Reduces Fear Memory Expression and Indices of Stress and Anxiety. J. Neurosci. 2018, 7, 1275–1292. [Google Scholar] [CrossRef]
- Gaspar, J.C.; Healy, C.; Ferdousi, M.I.; Roche, M.; Finn, D.P. Pharmacological blockade of pparα exacerbates inflammatory pain-related impairment of spatial memory in rats. Biomedicines 2021, 9, 610. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Yue, J.; Hu, L.-N.; Cheng, L.-F.; Wang, X.-S.; Wang, X.-J.; Feng, B. Chronic minocycline treatment reduces the anxiety-like behaviors induced by repeated restraint stress through modulating neuroinflammation. Brain Res. Bull. 2018, 143, 19–26. [Google Scholar] [CrossRef]
- Taylor, B.K.; Dadia, N.; Yang, C.B.; Krishnan, S.; Badr, M. Peroxisome Proliferator-Activated Receptor Agonists Inhibit Inflammatory Edema and Hyperalgesia. Inflammation 2002, 26, 121–127. [Google Scholar] [CrossRef]
- Oliveira, A.C.P.; Bertollo, C.M.; Rocha, L.T.S.; Nascimento, E.B.; Costa, K.A.; Coelho, M.M. Antinociceptive and antiedematogenic activities of fenofibrate, an agonist of PPAR alpha, and pioglitazone, an agonist of PPAR gamma. Eur. J. Pharmacol. 2007, 561, 194–201. [Google Scholar] [CrossRef]
- Suardíaz, M.; Estivill-Torrús, G.; Goicoechea, C.; Bilbao, A.; Rodríguez de Fonseca, F. Analgesic properties of oleoylethanolamide (OEA) in visceral and inflammatory pain. Pain 2007, 133, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Gill, N.; Bijjem, K.R.V.; Sharma, P.L. Anti-inflammatory and anti-hyperalgesic effect of all-trans retinoic acid in carrageenan-induced paw edema in Wistar rats: Involvement of peroxisome proliferator-activated receptor-β/δ receptors. Indian J. Pharmacol. 2013, 45, 278–282. [Google Scholar] [PubMed]
- Mansouri, M.T.; Naghizadeh, B.; Ghorbanzadeh, B.; Rajabi, H.; Pashmforoush, M. Pharmacological evidence for systemic and peripheral antinociceptive activities of pioglitazone in the rat formalin test: Role of PPARγ and nitric oxide. Eur. J. Pharmacol. 2017, 805, 84–92. [Google Scholar] [CrossRef]
- Donvito, G.; Bagdas, D.; Toma, W.; Rahimpour, E.; Jackson, A.; Meade, J.A.; AlSharari, S.; Kulkarni, A.R.; Ivy Carroll, F.; Lichtman, A.H.; et al. The interaction between alpha 7 nicotinic acetylcholine receptor and nuclear peroxisome proliferator-activated receptor-α represents a new antinociceptive signaling pathway in mice. Exp. Neurol. 2017, 295, 194–201. [Google Scholar] [CrossRef]
- Roche, M.; Johnston, P.; Mhuircheartaigh, O.N.; Olango, W.M.; Mackie, K.; Finn, D.P. Effects of intra-basolateral amygdala administration of rimonabant on nociceptive behaviour and neuronal activity in the presence or absence of contextual fear. Eur. J. Pain 2010, 14, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Ford, G.K.; Kieran, S.; Dolan, K.; Harhen, B.; Finn, D.P. A role for the ventral hippocampal endocannabinoid system in fear-conditioned analgesia and fear responding in the presence of nociceptive tone in rats. Pain 2011, 152, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Rea, K.; Roche, M.; Finn, D.P. Modulation of Conditioned Fear, Fear-Conditioned Analgesia, and Brain Regional C-Fos Expression Following Administration of Muscimol into the Rat Basolateral Amygdala. J. Pain 2011, 12, 712–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, K.; Ford, G.K.; Olango, W.M.; Harhen, B.; Roche, M.; Finn, D.P. Microinjection of 2-arachidonoyl glycerol into the rat ventral hippocampus differentially modulates contextually induced fear, depending on a persistent pain state. Eur. J. Neurosci. 2014, 39, 435–443. [Google Scholar] [CrossRef]
- Butler, R.K.; Ford, G.K.; Hogan, M.; Roche, M.; Doyle, K.M.; Kelly, J.P.; Kendall, D.A.; Chapman, V.; Finn, D.P. Fear-induced suppression of nociceptive behaviour and activation of Akt signalling in the rat periaqueductal grey: Role of fatty acid amide hydrolase. J Psychopharmacol 2012, 26, 83–91. [Google Scholar] [CrossRef]
- Olango, W.M.; Roche, M.; Ford, G.K.; Harhen, B.; Finn, D.P. The endocannabinoid system in the rat dorsolateral periaqueductal grey mediates fear-conditioned analgesia and controls fear expression in the presence of nociceptive tone. Br. J. Pharmacol. 2012, 165, 2549–2560. [Google Scholar] [CrossRef] [Green Version]
- Zanoveli, J.M.; Carvalho, M.C.; Cunha, J.M.; Brandão, M.L. Extracellular serotonin level in the basolateral nucleus of the amygdala and dorsal periaqueductal gray under unconditioned and conditioned fear states: An in vivo microdialysis study. Brain Res. 2009, 1294, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Deutch, A.; Charney, D.S. A Functional Neuroanatomy of Anxiety and Fear: Implications for the Pathophysiology and Treatment of Anxiety Disorders. Crit. Rev. Neurobiol. 1996, 10, 419–446. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.; Rainnie, D.; Cassell, M. Neurotransmission in the rat amygdala related to fear and anxiety. Trends Neurosci. 1994, 17, 208–214. [Google Scholar] [CrossRef]
- Macedo, C.E.; Martinez, R.C.R.; Albrechet-Souza, L.; Molina, V.A.; Brandão, M.L. 5-HT2- and D1-mechanisms of the basolateral nucleus of the amygdala enhance conditioned fear and impair unconditioned fear. Behav. Brain Res. 2007, 177, 100–108. [Google Scholar] [CrossRef]
- Taylor, A.M.W.; Becker, S.; Schweinhardt, P.; Cahill, C. Mesolimbic dopamine signaling in acute and chronic pain: Implications for motivation, analgesia, and addiction. Pain 2016, 157, 1194–1198. [Google Scholar] [CrossRef] [Green Version]
- Benarroch, E.E. Involvement of the nucleus accumbens and dopamine system in chronic pain. Neurology 2016, 87, 1720–1726. [Google Scholar] [CrossRef]
- Ikeda, H.; Takasu, S.; Murase, K. Contribution of Anterior Cingulate Cortex and Descending Pain Inhibitory System to Analgesic Effect of Lemon Odor in Mice. Mol. Pain 2014, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wood, P.B. Role of central dopamine in pain and analgesia. Expert Rev. Neurother. 2008, 8, 781–797. [Google Scholar] [CrossRef]
- Roche, M.; O’Connor, E.; Diskin, C.; Finn, D.P. The effect of CB1 receptor antagonism in the right basolateral amygdala on conditioned fear and associated analgesia in rats. Eur. J. Neurosci. 2007, 26, 2643–2653. [Google Scholar] [CrossRef]
- Hagiwara, H.; Funabashi, T.; Akema, T.; Kimura, F. Sex-specific differences in pain response by dopamine in the bed nucleus of the stria terminalis in rats. Neuroreport 2013, 24, 181–185. [Google Scholar] [CrossRef]
- George, P.; Watson, C. The Rat Brain, in Stereotaxic Coordinates; Academic Press: San Diego, MO, USA, 1997. [Google Scholar]
- Morgenweck, J.; Abdel-aleem, O.S.; McNamara, K.C.; Donahue, R.R.; Badr, M.Z.; Taylor, B.K. Activation of peroxisome proliferator-activated receptor γ in brain inhibits inflammatory pain, dorsal horn expression of Fos, and local edema. Neuropharmacology 2010, 58, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, D.P.; Beckett, S.R.G.; Richardson, D.; Kendall, D.A.; Marsden, C.A.; Chapman, V. Evidence for differential modulation of conditioned aversion and fear-conditioned analgesia by CB1 receptors. Eur. J. Neurosci. 2004, 20, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.K.; Rea, K.; Lang, Y.; Gavin, A.M.; Finn, D.P. Endocannabinoid-mediated enhancement of fear-conditioned analgesia in rats: Opioid receptor dependency and molecular correlates. Pain 2008, 140, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Rea, K.; McGowan, F.; Corcoran, L.; Roche, M.; Finn, D.P. The prefrontal cortical endocannabinoid system modulates fear-pain interactions in a subregion-specific manner. Br. J. Pharmacol. 2019, 176, 1492–1505. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.S.; Sufka, K.J.; Coderre, T.J. Optimal scoring strategies and weights for the formalin test in rats. Pain 1997, 70, 53–58. [Google Scholar] [CrossRef]
- Ford, G.K.; Moriarty, O.; McGuire, B.E.; Finn, D.P. Investigating the effects of distracting stimuli on nociceptive behaviour and associated alterations in brain monoamines in rats. Eur. J. Pain 2008, 12, 970–979. [Google Scholar] [CrossRef]
- Kerr, D.M.; Burke, N.N.; Ford, G.K.; Connor, T.J.; Harhen, B.; Egan, L.J.; Finn, D.P.; Roche, M. Pharmacological inhibition of endocannabinoid degradation modulates the expression of inflammatory mediators in the hypothalamus following an immunological stressor. Neuroscience 2012, 204, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Thunder, M.; Moore, D.S.; McCabe, G.P. Introduction to the Practice of Statistics; WH Freeman: New York, NY, USA, 2007; Volume 79. [Google Scholar]






















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaspar, J.C.; Okine, B.N.; Dinneen, D.; Roche, M.; Finn, D.P. Effects of Intra-BLA Administration of PPAR Antagonists on Formalin-Evoked Nociceptive Behaviour, Fear-Conditioned Analgesia, and Conditioned Fear in the Presence or Absence of Nociceptive Tone in Rats. Molecules 2022, 27, 2021. https://doi.org/10.3390/molecules27062021
Gaspar JC, Okine BN, Dinneen D, Roche M, Finn DP. Effects of Intra-BLA Administration of PPAR Antagonists on Formalin-Evoked Nociceptive Behaviour, Fear-Conditioned Analgesia, and Conditioned Fear in the Presence or Absence of Nociceptive Tone in Rats. Molecules. 2022; 27(6):2021. https://doi.org/10.3390/molecules27062021
Chicago/Turabian StyleGaspar, Jessica C., Bright N. Okine, David Dinneen, Michelle Roche, and David P. Finn. 2022. "Effects of Intra-BLA Administration of PPAR Antagonists on Formalin-Evoked Nociceptive Behaviour, Fear-Conditioned Analgesia, and Conditioned Fear in the Presence or Absence of Nociceptive Tone in Rats" Molecules 27, no. 6: 2021. https://doi.org/10.3390/molecules27062021