
The Protective Effects of Sour Orange (Citrus aurantium L.) Polymethoxyflavones on Mice Irradiation-Induced Intestinal Injury


Abstract
:1. Introduction
2. Results
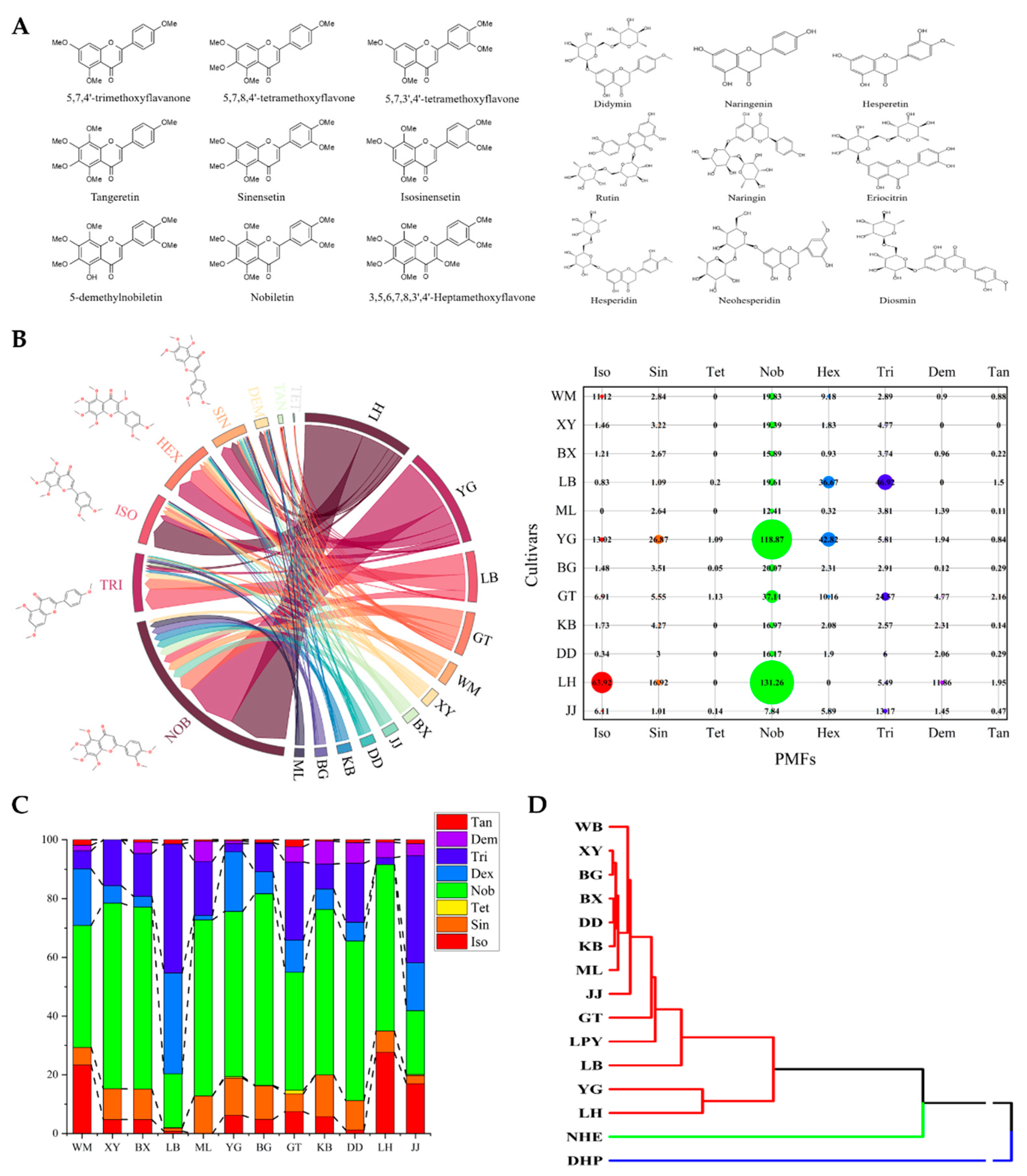
2.1. Extraction and Detection of Polymethoxyflavones in Sour Oranges
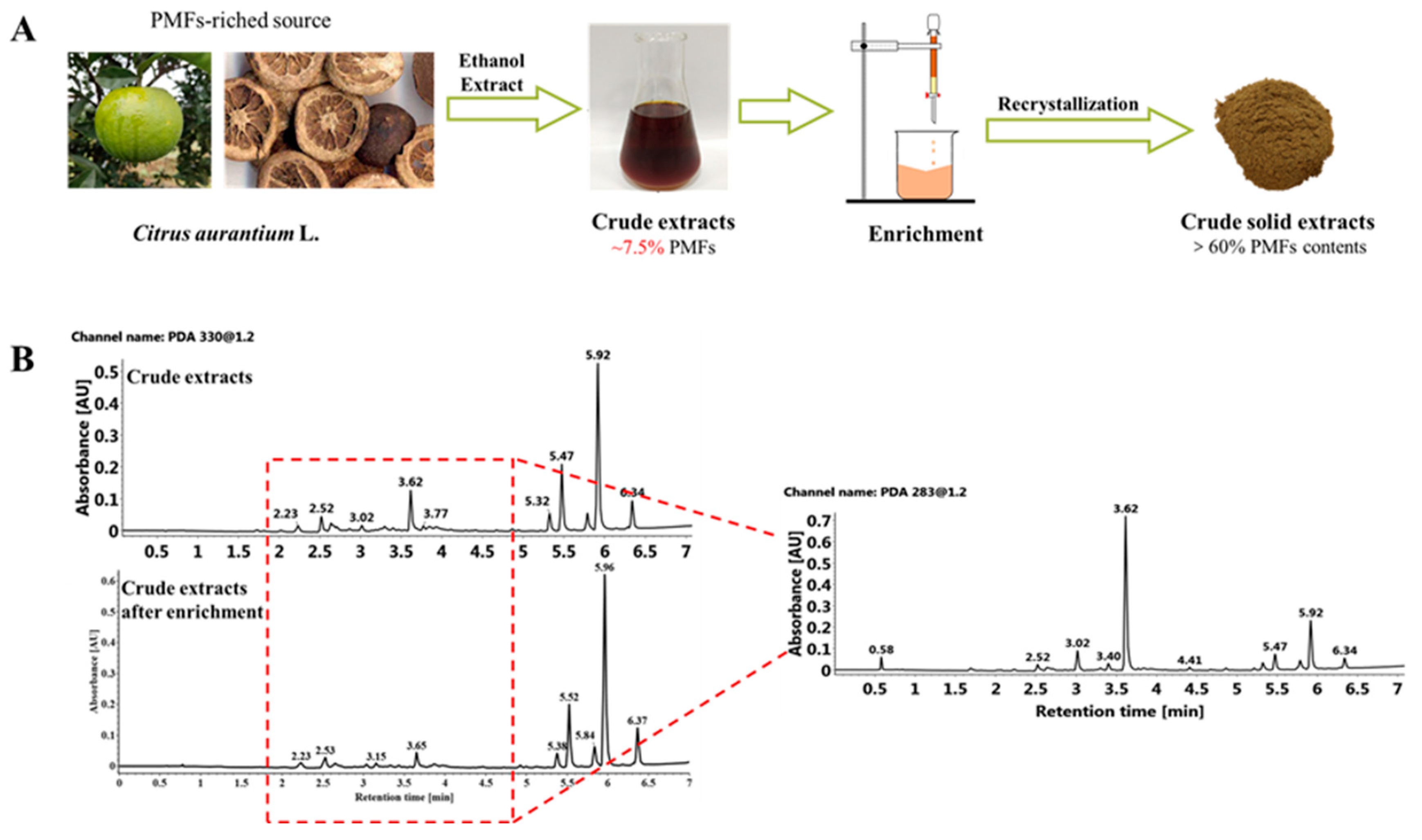
2.2. Purification of Sour Orange PMFs
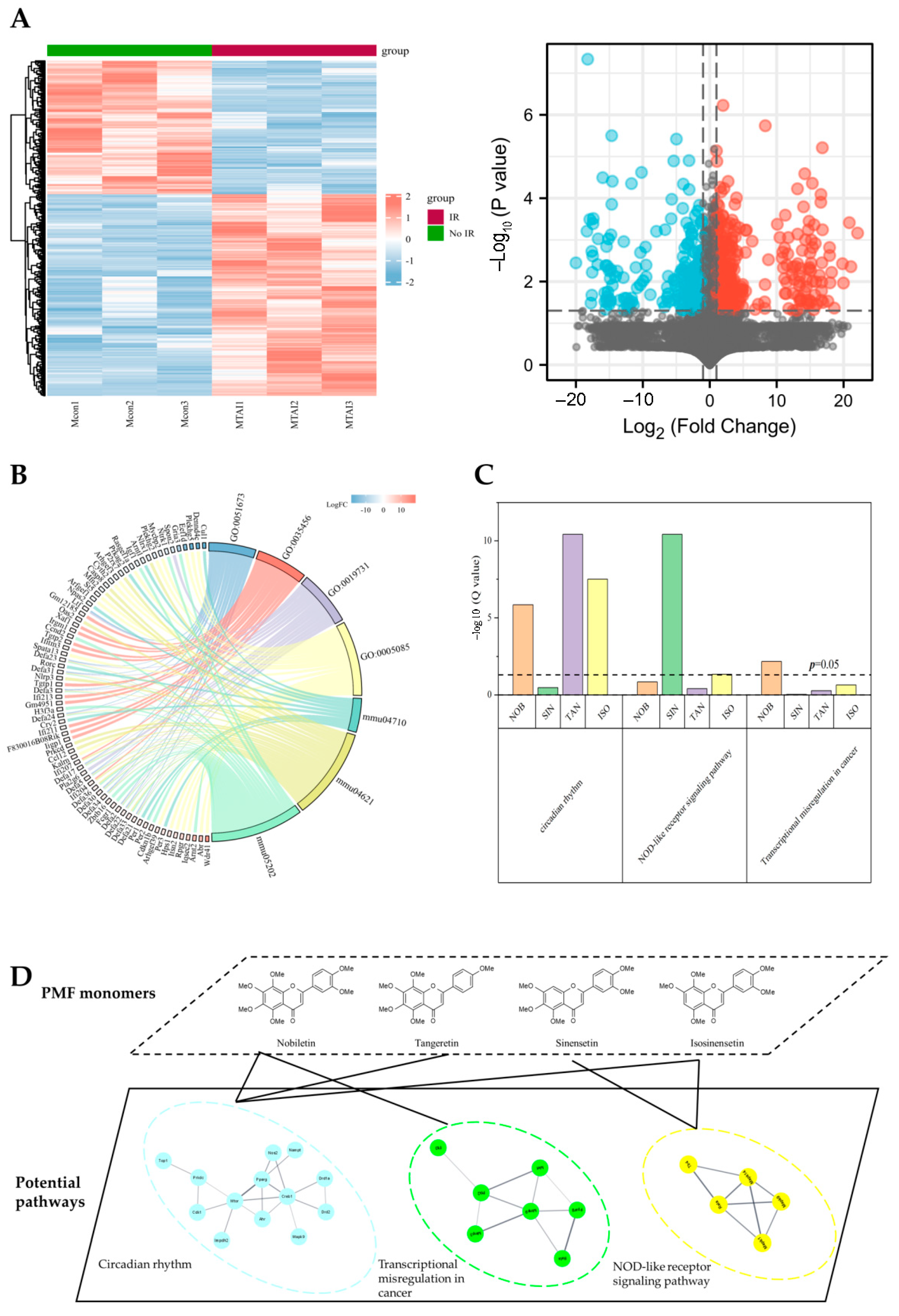
2.3. Potential Protective Effects of Sour Orange PMFs on Mice ARS
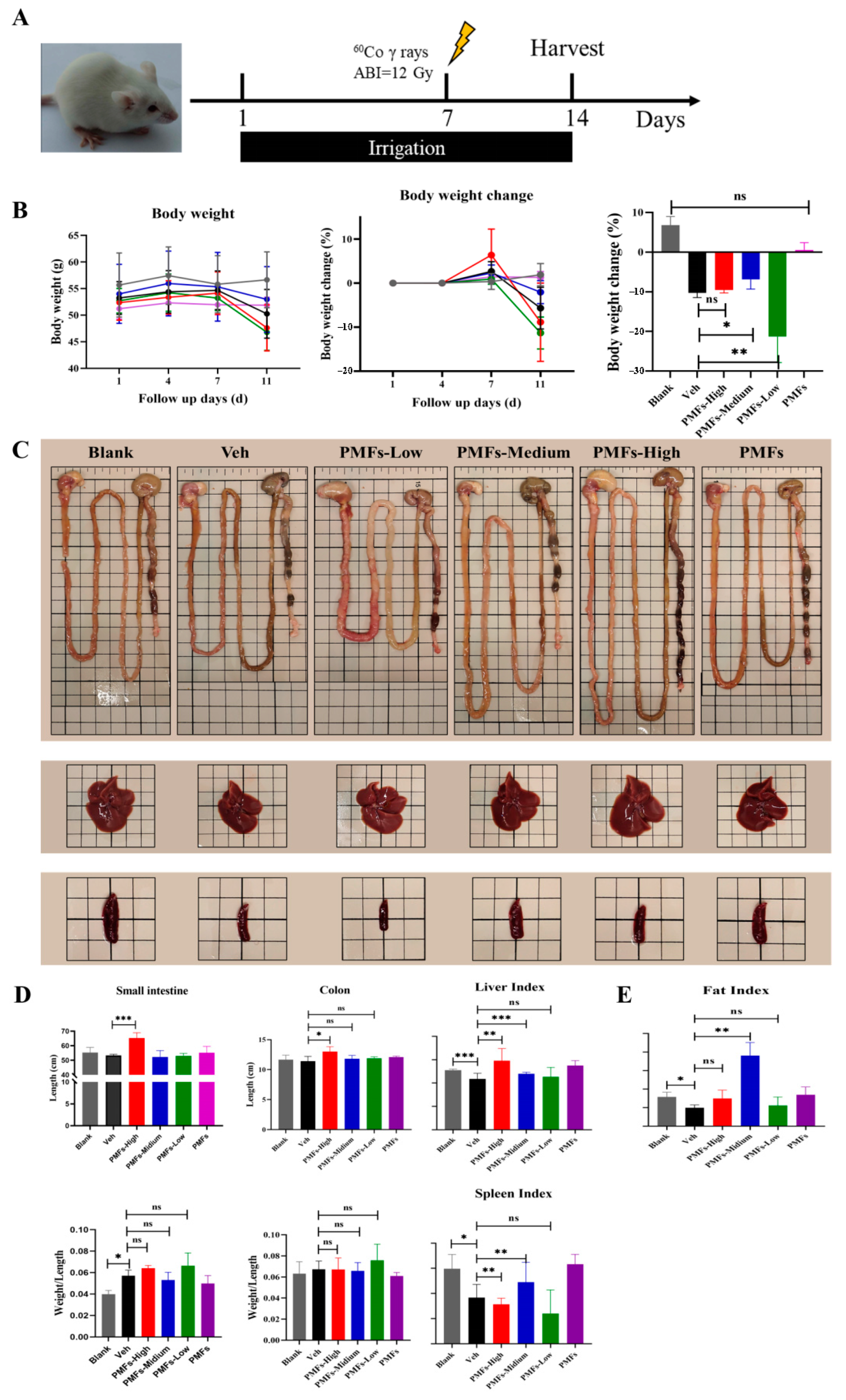
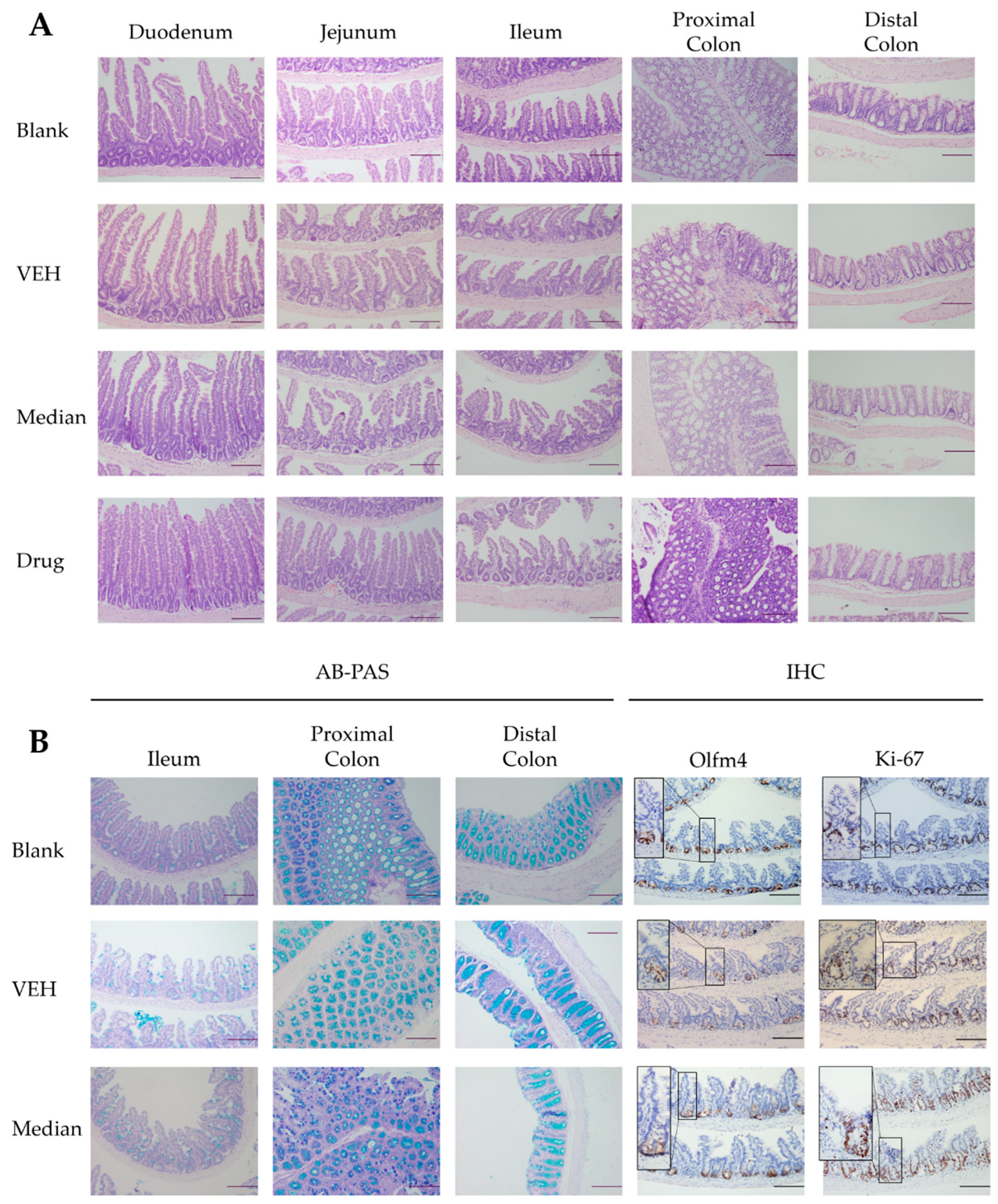
2.4. The Protective Effects of PMFs in Radiation-Induced Intestinal Injury Mice Model
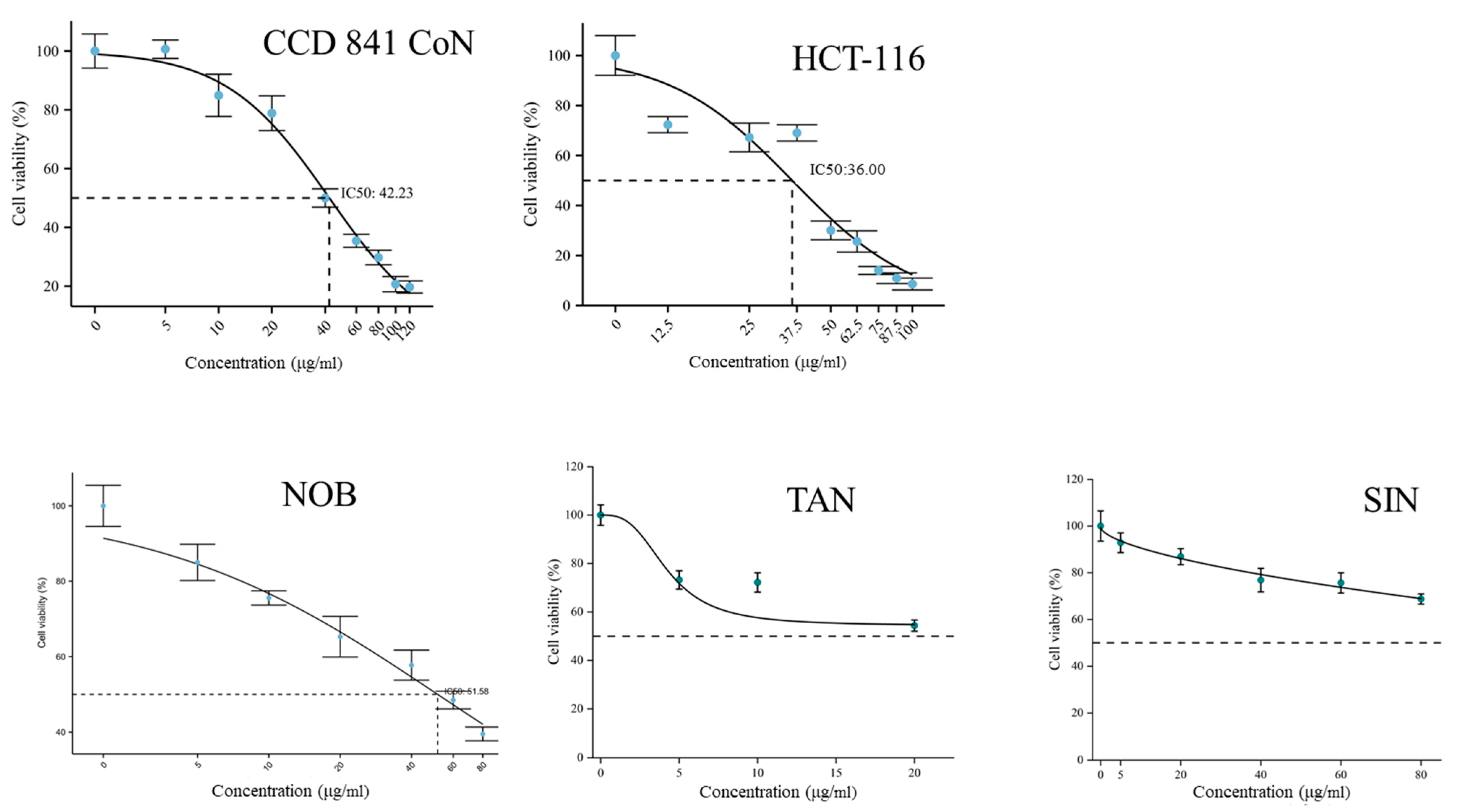
2.5. In Vitro Cytotoxicity Test of PMFs
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. PMF Extraction and Detection
4.3. Purification of PMFs in Selected Sour Orange Cultivar
4.4. Bioinformatics and Network Pharmacology Analysis of Mice ARS
4.5. In Vivo Experiments
4.6. In Vitro Experiments
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Parikh, K.; Antanaviciute, A.; Fawkner-Corbett, D.; Jagielowicz, M.; Aulicino, A.; Lagerholm, C.; Davis, S.; Kinchen, J.; Chen, H.H.; Alham, N.K.; et al. Colonic epithelial cell diversity in health and inflammatory bowel disease. Nature 2019, 567, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Bonis, V.; Rossell, C.; Gehart, H. The Intestinal Epithelium—Fluid Fate and Rigid Structure From Crypt Bottom to Villus Tip. Front. Cell Dev. Biol. 2021, 9, 661931. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, K.; Hayakawa, Y.; Koike, K. Plasticity of Intestinal Epithelium: Stem Cell Niches and Regulatory Signals. Int. J. Mol. Sci. 2021, 22, 357. [Google Scholar] [CrossRef] [PubMed]
- Flores-Cisneros, L.; Castro-Eguiluz, D.; Reyes-Barreter, D.Y.; Jaimes, E.; Cano-Blanco, C.; Avendano-Perez, C.; Carbajal-Lopez, B.; Sanchez-Lopez, M.; Mota-Garcia, A.; Gallardo-Rincon, D.; et al. Effects of dietary components during and after concomitant chemoradiotherapy, radiotherapy, or sequential chemoradiotherapy to the abdominopelvic area. Rev. Investig. Clin. Transl. Investig. 2018, 70, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Sapozhnikov, R.Y.; Khalimov, Y.S.; Matveev, S.Y.; Gaiduk, S.V.; Legeza, V.I.; Drachev, I.S. Modern condition and prospects of development of means and methods of early pathogenetic therapy of acute radiation. Bull. Russ. Mil. Med. Acad. 2019, 3, 201–205. [Google Scholar]
- Xiong, Y.-J.; Deng, Z.-B.; Liu, J.-N.; Qiu, J.-J.; Guo, L.; Feng, P.-P.; Sui, J.-R.; Chen, D.-P.; Guo, H.-S. Enhancement of epithelial cell autophagy induced by sinensetin alleviates epithelial barrier dysfunction in colitis. Pharmacol. Res. 2019, 148, 104461. [Google Scholar] [CrossRef]
- Gao, Y.-L.; Shao, L.-H.; Dong, L.-H.; Chang, P.-Y. Gut commensal bacteria, Paneth cells and their relations to radiation enteropathy. World J. Stem Cells 2020, 12, 188–202. [Google Scholar] [CrossRef]
- Hollingsworth, B.A.; Cassatt, D.R.; DiCarlo, A.L.; Rios, C.I.; Satyamitra, M.M.; Winters, T.A.; Taliaferro, L.P. Acute Radiation Syndrome and the Microbiome: Impact and Review. Front. Pharmacol. 2021, 12, 643283. [Google Scholar] [CrossRef]
- Ghosh, S.; Whitley, C.S.; Haribabu, B.; Jala, V.R. Regulation of Intestinal Barrier Function by Microbial Metabolites. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1463–1482. [Google Scholar] [CrossRef]
- Maksoud, S.; Abdel-Massih, R.M.; Rajha, H.N.; Louka, N.; Chemat, F.; Barba, F.J.; Debs, E. Citrus aurantium L. Active constituents, biological effects and extraction methods. An updated review. Molecules 2021, 26, 5832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, S.; Ho, C.-T.; Huang, Q. Citrus polymethoxyflavones as regulators of metabolic homoeostasis: Recent advances for possible mechanisms. Trends Food Sci. Technol. 2021, 110, 743–753. [Google Scholar] [CrossRef]
- Nelson, E.K. The occurrence of a pentamethyl flavonol m tangerine peel. J. Am. Chem. Soc. 1934, 56, 1392–1393. [Google Scholar] [CrossRef]
- Zeng, S.-L.; Li, S.-Z.; Xiao, P.-T.; Cai, Y.-Y.; Chu, C.; Chen, B.-Z.; Li, P.; Li, J.; Liu, E.-H. Citrus polymethoxyflavones attenuate metabolic syndrome by regulating gut microbiome and amino acid metabolism. Sci. Adv. 2020, 6, eaax6208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Li, D.; Xu, W.; Zhang, H.; Wang, H.; Perdew, G.H. β-Naphthoflavone Activation of the Ah Receptor Alleviates Irradiation-Induced Intestinal Injury in Mice. Antioxidants 2020, 9, 1264. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; He, S.; Hu, Y.; Yang, Y.; Jiao, B.; Fang, Q.; Zhou, Z. Fruit flavonoid variation between and within four cultivated Citrus species evaluated by UPLC-PDA system. Sci. Hortic. 2017, 224, 93–101. [Google Scholar] [CrossRef]
- McGough, C.; Baldwin, C.; Norman, A.; Frost, G.; Blake, P.; Tait, D.; Khoo, V.; Harrington, K.; Andreyev, H.J.N. Is supplementation with elemental diet feasible in patients undergoing pelvic radiotherapy? Clin. Nutr. 2006, 25, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Park, S.; Shim, S.; Myung, J.K. Abstract 2930: Baicalein attenuates radiation-induced enteritis by improving endothelial dysfunction. Cancer Res. 2019, 79, 2930. [Google Scholar] [CrossRef]
- Gao, Z.; Gao, W.; Zeng, S.-L.; Li, P.; Liu, E.-H. Chemical structures, bioactivities and molecular mechanisms of citrus polymethoxyflavones. J. Funct. Foods 2018, 40, 498–509. [Google Scholar] [CrossRef]
- Peng, Z.; Zhang, H.; Li, W.; Yuan, Z.; Xie, Z.; Zhang, H.; Cheng, Y.; Chen, J.; Xu, J. Comparative profiling and natural variation of polymethoxylated flavones in various citrus germplasms. Food Chem. 2021, 354, 129499. [Google Scholar] [CrossRef]
- Scora, R.W. On the History and Origin of Citrus. Bull. Torrey Bot. Club 1975, 102, 369–375. [Google Scholar] [CrossRef]
- Barrett, H.C.; Rhodes, A.M. A Numerical Taxonomic Study of Affinity Relationships in Cultivated Citrus and Its Close Relatives. Syst. Bot. 1976, 1, 105. [Google Scholar] [CrossRef]
- Li, S. Exploring traditional chinese medicine by a novel therapeutic concept of network target. Chin. J. Integr. Med. 2016, 22, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Lai, X.; Wang, X.; Yao, X.; Wang, W.; Li, S. Network pharmacology to explore the anti-inflammatory mechanism of Xuebijing in the treatment of sepsis. Phytomedicine 2021, 85, 153543. [Google Scholar] [CrossRef] [PubMed]
- In, J.G.; Foulke-Abel, J.; Estes, M.K.; Zachos, N.C.; Kovbasnjuk, O.; Donowitz, M. Human mini-guts: New insights into intestinal physiology and host-pathogen interactions. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.R.; Howard, L.A.; Harrison, D.J.; Winton, D.J. p53, mutation frequency and apoptosis in the murine small intestine. Oncogene 1997, 14, 2015–2018. [Google Scholar] [CrossRef]
- Soltanmohammadi, N.; Wang, S.; Schumacher, B. Somatic PMK-1/p38 signaling links environmental stress to germ cell apoptosis and heritable euploidy. Nat. Commun. 2022, 13, 701. [Google Scholar] [CrossRef]
- Rodrigues, P.; Macaya, I.; Bazzocco, S.; Mazzolini, R.; Andretta, E.; Dopeso, H.; Mateo-Lozano, S.; Bilić, J.; Cartón-García, F.; Nieto, R.; et al. RHOA inactivation enhances Wnt signalling and promotes colorectal cancer. Nat. Commun. 2014, 5, 5458. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Microbiol. 2018, 16, 457–470. [Google Scholar] [CrossRef]
- Ray, K. Intercrypt goblet cells—The key to colonic mucus barrier function. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 455. [Google Scholar] [CrossRef]
- Nyström, E.E.L.; Martinez-Abad, B.; Arike, L.; Birchenough, G.M.H.; Nonnecke, E.B.; Castillo, P.A.; Svensson, F.; Bevins, C.L.; Hansson, G.C.; Johansson, M.E.V. An intercrypt subpopulation of goblet cells is essential for colonic mucus barrier function. Science 2021, 372, eabb1590. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Hsieh, J.J.; Liu, C.Y.; Appel, K.L.; Waddell, A.; Almohazey, D.; Katada, K.; Bernard, J.K.; Bucar, E.B.; Gadeock, S.; et al. Sprouty2 limits intestinal tuft and goblet cell numbers through GSK3β-mediated restriction of epithelial IL-33. Nat. Commun. 2021, 12, 836. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Luqman, S.; Meena, A. Prospects of tangeretin as a modulator of cancer targets/pathways. Pharmacol. Res. 2020, 161, 105202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xin, Y.; Feng, K.; Yin, B.; Kan, Q.; Xiao, J.; Cao, Y.; Ho, C.T.; Huang, Q. Comparative analyses of bioavailability, biotransformation, and excretion of nobiletin in lean and obese rats. J. Agric. Food Chem. 2020, 68, 10709–10718. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhang, Z.; Zhai, Y.; Yan, X.; Zhou, W.; Liu, H.; Guan, L.; Peng, L. Apigenin Alleviates Obesity-Associated Metabolic Syndrome by Regulating the Composition of the Gut Microbiome. Front. Microbiol. 2022, 12, 805827. [Google Scholar] [CrossRef] [PubMed]
- Segers, C.; Mysara, M.; Claesen, J.; Baatout, S.; Leys, N.; Lebeer, S.; Verslegers, M.; Mastroleo, F. Intestinal mucositis precedes dysbiosis in a mouse model for pelvic irradiation. ISME Commun. 2021, 1, 24. [Google Scholar] [CrossRef]
- Hauer-Jensen, M.; Denham, J.W.; Andreyev, H.J.N. Radiation enteropathy-pathogenesis, treatment and prevention. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Nemoto, K.; Ohizumi, Y. An evaluation of the genotoxicity and subchronic toxicity of the peel extract of Ponkan cultivar ‘Ohta ponkan’ (Citrus reticulata Blanco) that is rich in nobiletin and tangeretin with anti-dementia activity. Regul. Toxicol. Pharmacol. 2020, 114, 104670. [Google Scholar] [CrossRef] [PubMed]
- Ting, Y.; Chiou, Y.; Jiang, Y.; Pan, M.; Lin, Z.; Huang, Q. Safety evaluation of tangeretin and the effect of using emulsion-based delivery system: Oral acute and 28-day sub-acute toxicity study using mice. Food Res. Int. 2015, 74, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, X.J.; Pan, Y.; Zhou, Z. Identification of the chemical compositions of Ponkan peel by ultra performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. Anal. Methods 2016, 8, 893–903. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, Z.; Zhou, Z. Simultaneous separation and purification of five polymethoxylated flavones from “dahongpao” tangerine (Citrus tangerina tanaka) using macroporous adsorptive resins combined with prep-HPLC. Molecules 2018, 23, 2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germplasm Code 1 | Germplasm Name 2 | Place of Origin | Abbreviation |
|---|---|---|---|
| LA0002 | Jiangjin suancheng | China | JJ |
| LA0003 | Daidai | China | DD |
| LA0004 | Hangbulaite suancheng | U.S.A. | KB |
| LA0008 | Goutoucheng | China | GT |
| LA0012 | Banggan | China | BG |
| LA0015 | Yaogan-1 | China | YG |
| LA0018 | Moluogesuancheng | Morocco | ML |
| LA0022 | Lubidukesi suancheng | Mexico | LB |
| LA0038 | Baxi suancheng | Brazil | BX |
| LA0048 | Xiaoye suancheng | U.S.A. | XY |
| LA0055 | Lianhe suancheng | China | LH |
| LA0057 | Wanmucheng | Japan | WM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Li, Z.; Wang, F.; Zhou, Z. The Protective Effects of Sour Orange (Citrus aurantium L.) Polymethoxyflavones on Mice Irradiation-Induced Intestinal Injury. Molecules 2022, 27, 1934. https://doi.org/10.3390/molecules27061934
Jiang Z, Li Z, Wang F, Zhou Z. The Protective Effects of Sour Orange (Citrus aurantium L.) Polymethoxyflavones on Mice Irradiation-Induced Intestinal Injury. Molecules. 2022; 27(6):1934. https://doi.org/10.3390/molecules27061934
Chicago/Turabian StyleJiang, Zixiao, Zhenqing Li, Fengchao Wang, and Zhiqin Zhou. 2022. "The Protective Effects of Sour Orange (Citrus aurantium L.) Polymethoxyflavones on Mice Irradiation-Induced Intestinal Injury" Molecules 27, no. 6: 1934. https://doi.org/10.3390/molecules27061934