
Laser Light Treatment Improves the Mineral Composition, Essential Oil Production and Antimicrobial Activity of Mycorrhizal Treated Pelargoniumgraveolens

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Laser Improved Mycorrhizal Colonization in P. graveolens Plants
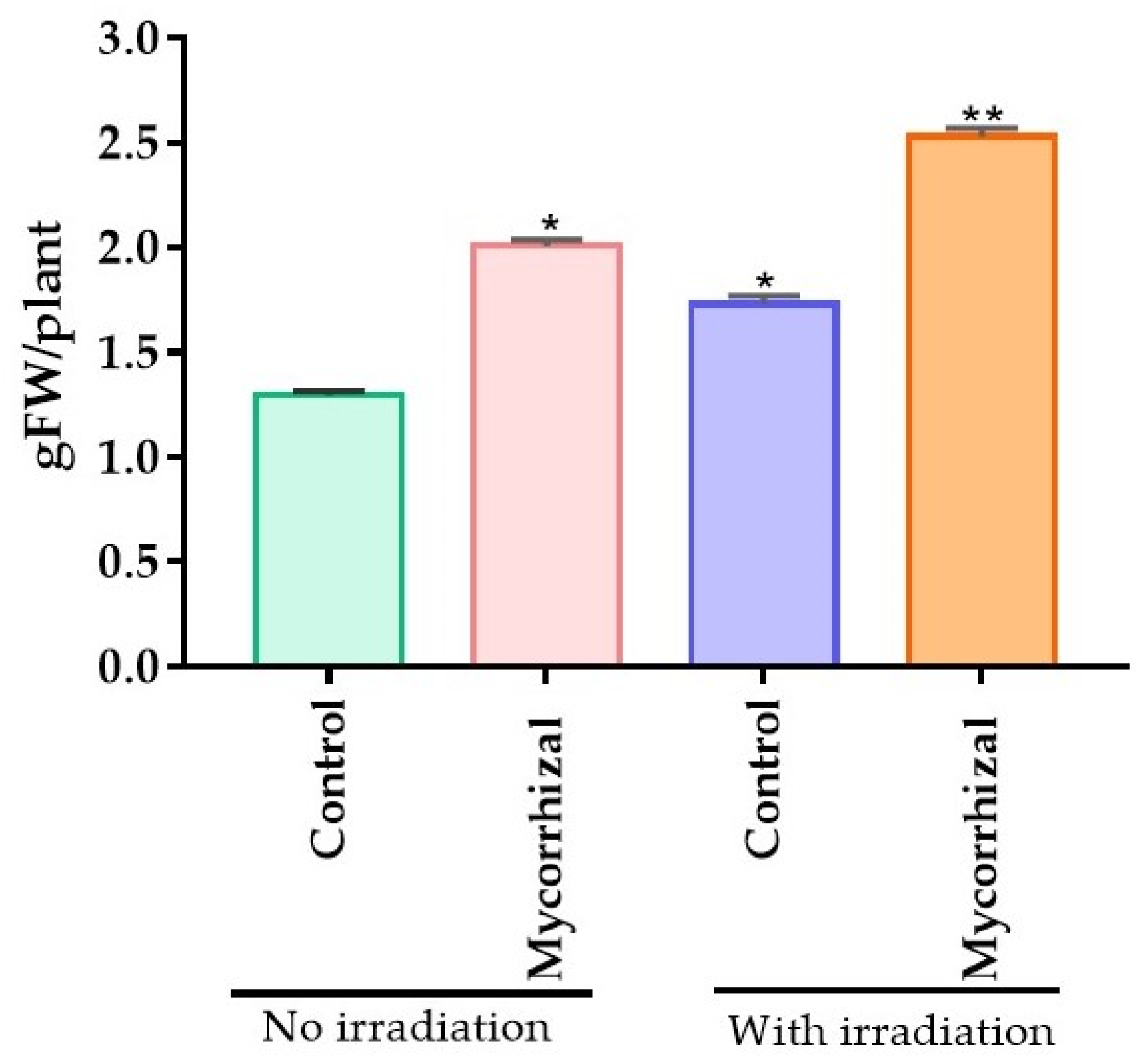
2.2. Improved Colonization in Plant-Initiated from Laser Light Treated Seeds Increased Biomass Accumulation and Biological Value of P. graveolens
2.3. Mycorrhiza Induced Mineral Uptake by P. graveolens Is Improved by Laser Light Treatment
2.4. Co-Application of Mycorrhiza and Laser Light Treatment Induced the Production of Essential Oils and Their Precursors
2.5. Laser Light Boosted the Antibacterial Properties of Mycorrhiza Inoculated P. graveolens Plant
3. Discussion
4. Materials and Methods
4.1. AMF Symbiont
4.2. Experimental Setup
4.3. AMF Growth Analysis
4.4. Elemental Analysis
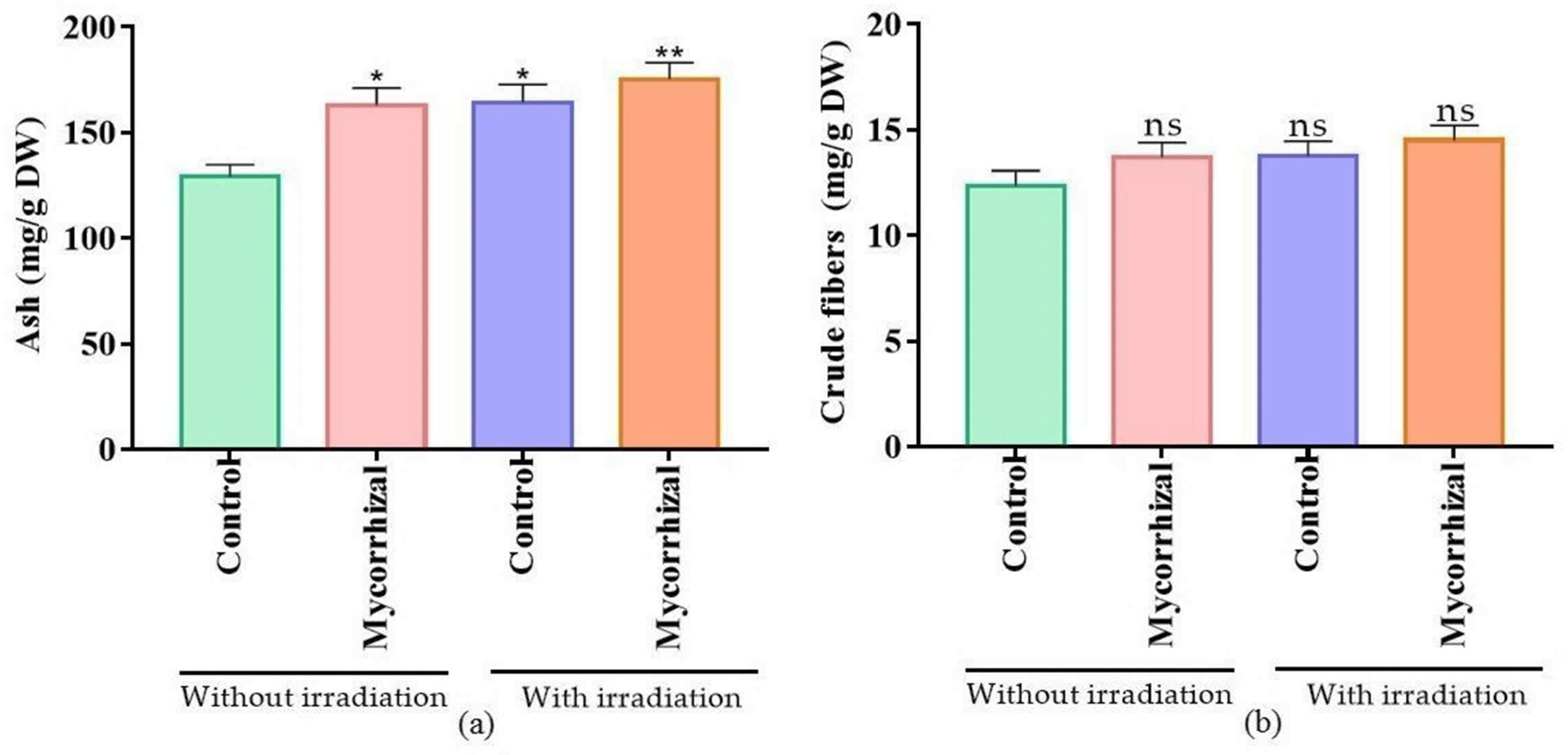
4.5. Determination of the Nutritional Value
Proximate Composition Analysis
4.6. Determination of Essential Oils
4.7. Antibacterial Activity
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Romano, B.; Lucariello, G.; Capasso, R. Topical collection: Pharmacology of medicinal plants. Biomolecules 2021, 11, 101. [Google Scholar] [CrossRef]
- Babich, O.; Sukhikh, S.; Prosekov, A.; Asyakina, L.; Ivanova, S. Medicinal plants to strengthen immunity during a pandemic. Pharmaceuticals 2020, 13, 313. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M.E.; Chávez, Á.; Ferrer, A.; Phillips, M.A. Distinct metabolic pathways drive monoterpenoid biosynthesis in a natural population of Pelargonium graveolens. J. Exp. Bot. 2020, 71, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Ennaifer, M.; Bouzaiene, T.; Chouaibi, M.; Hamdi, M. Pelargonium graveolens aqueous decoction: A new water-soluble polysaccharide and antioxidant-rich extract. Biomed. Res. Int. 2018, 2018, 2691513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsouna, A.B.; Hamdi, N. Phytochemical composition and antimicrobial activities of the essential oils and organic extracts from Pelargonium graveolens growing in Tunisia. Lipids Health Dis. 2012, 11, 167. [Google Scholar] [CrossRef] [Green Version]
- Lira, M.H.P.D.; Andrade Júnior, F.P.D.; Moraes, G.F.Q.; Macena, G.D.S.; Pereira, F.D.O.; Lima, I.O. Antimicrobial activity of geraniol: An integrative review. J. Essent. Oil Res. 2020, 32, 187–197. [Google Scholar] [CrossRef]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial activity of terpenes and terpenoids present in essential oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [Green Version]
- Sikkema, J.; de Bont, J.A.M.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [CrossRef]
- Boukhatem, M.N.; Kameli, A.; Ferhat, M.A.; Saidi, F.; Mekarnia, M. Rose geranium essential oil as a source of new and safe anti-inflammatory drugs. Libyan J. Med. 2013, 8, 22520. [Google Scholar] [CrossRef]
- Koduru, J.R.; Kailasa, S.K.; Bhamore, J.R.; Kim, K.H.; Dutta, T.; Vellingiri, K. Phytochemical-assisted synthetic approaches for silver nanoparticles antimicrobial applications: A review. Adv. Colloid Interface Sci. 2018, 256, 326–339. [Google Scholar] [CrossRef]
- Rolim, W.R.; Pelegrino, M.T.; de Araújo Lima, B.; Ferraz, L.S.; Costa, F.N.; Bernardes, J.S.; Rodigues, T.; Brocchi, M.; Seabra, A.B. Green tea extract mediated biogenic synthesis of silver nanoparticles: Characterization, cytotoxicity evaluation and antibacterial activity. Appl. Surf. Sci. 2019, 463, 66–74. [Google Scholar] [CrossRef]
- Senthilkumar, S.R.; Sivakumar, T. Green tea (Camellia sinensis) mediated synthesis of zinc oxide (ZnO) nanoparticles and studies on their antimicrobial activities. Int. J. Pharm. Pharm. Sci. 2014, 6, 461–465. [Google Scholar]
- Możdżeń, K.; Barabasz-Krasny, B.; Zandi, P. Effect of long-term of He-Ne laser light irradiation on selected physiological processes of Triticale. Plants 2020, 9, 1703. [Google Scholar] [CrossRef] [PubMed]
- Mathiassen, S.K.; Bak, T.; Christensen, S.; Kudsk, P. The effect of laser treatment as a weed control method. Biosyst. Eng. 2006, 94, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Muszyński, S.; Gładyszewska, B. Representation of He-Ne laser irradiation effect on radish seeds with selected germination indices. Int. Agrophys. 2008, 22, 151–157. [Google Scholar]
- Dziwulska, A.; Koper, R. Effect of pre-sowing laser biostimulation on germination of lucerne seeds. Acta Agrophys. 2003, 82, 33–39. [Google Scholar]
- Dziwulska, A.; Koper, R.; Wilczek, M. Estimation of effect of He-Ne laser beam on germination capacity of white clover variety of Anda cultivar. Acta Agrophys. 2004, 3, 435–441. [Google Scholar]
- Almuhayawi, M.S.; Hassan, A.H.; Abdel-Mawgoud, M.; Khamis, G.; Selim, S.; Al Jaouni, S.K.; AbdElgawad, H. Laser light as a promising approach to improve the nutritional value, antioxidant capacity and anti-inflammatory activity of flavonoid-rich buckwheat sprouts. Food Chem. 2021, 345, 128788. [Google Scholar] [CrossRef]
- Okla, M.K.; El-Tayeb, M.A.; Qahtan, A.A.; Abdel-Maksoud, M.A.; Elbadawi, Y.B.; Alaskary, M.K.; Balkhyour, M.A.; Hassan, A.H.; AbdElgawad, H. Laser light treatment of seeds for improving the biomass photosynthesis, chemical composition and biological activities of Lemongrass sprouts. Agronomy 2021, 11, 478. [Google Scholar] [CrossRef]
- Mahmood, S.; Afzal, B.; Perveen, S.; Wahid, A.; Azeem, M.; Iqbal, N. He-Ne laser seed treatment improves the nutraceutical metabolic pool of Sunflowers and provides better tolerance against water deficit. Front. Plant Sci. 2021, 12, 804. [Google Scholar] [CrossRef]
- Amiri, R.; Nikbakht, A.; Etemadi, N. Alleviation of drought stress on rose geranium [Pelargonium graveolens (L.) Herit.] in terms of antioxidant activity and secondary metabolites by mycorrhizal inoculation. Sci. Hortic. 2015, 197, 373–380. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jin, W.; Liu, A.; Zhang, S.; Liu, D.; Wang, F.; Lin, X.; He, C. Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress. Sci. Hortic. 2013, 160, 222–229. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Xia, R.X. Effects of water stress and arbuscular mycorrhizal fungi on reactive oxygen metabolism and antioxidant production by citrus (Citrus tangerine) roots. Eur. J. Soil Biol. 2006, 42, 166–172. [Google Scholar] [CrossRef]
- Ceccarelli, N.; Curadi, M.; Martelloni, L.; Sbrana, C.; Picciarelli, P.; Giovannetti, M. Mycorrhizal colonization impacts on phenolic content and antioxidant properties of artichoke leaves and flower heads two years after field transplant. Plant Soil. 2010, 335, 311–323. [Google Scholar] [CrossRef]
- Alam, M.; Khaliq, A.; Sattar, A.; Shukla, R.S.; Anwar, M.; Dharni, S. Synergistic effect of arbuscular mycorrhizal fungi and Bacillus subtilis on the biomass and essential oil yield of rose-scented geranium (Pelargonium graveolens). Arch. Agron. Soil Sci. 2011, 57, 889–898. [Google Scholar] [CrossRef]
- Friedmann, H.; Lubart, R.; Lanlicht, J.; Rochking, S. A possible explanation of laser-induced stimulation and damage of cell cultures. J. Phytochem. Photobiol. B Biol. 1991, 87, 87–91. [Google Scholar] [CrossRef]
- Dobrowolski, J.W.; Wachalewski, T.; Smyk, B.; Rózycki, E.; Barabasz, W. Experiments on the influence of laser light on some biological elements of the natural environment. Environ. Manag. Health 1997, 4, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.P.; Yue, M.; Wang, X.L. Influence of He–Ne laser irradiation on seeds thermodynamic parameters and seedlings growth of Isatis indogotica. Plant Sci. 2005, 168, 601–606. [Google Scholar] [CrossRef]
- Li, P.; Li, H.; Zong, Y.; Li, F.Y.; Han, Y.; Hao, X. Photosynthesis and metabolite responses of Isatis indigotica Fortune to elevated [CO2]. Crop J. 2017, 5, 345–353. [Google Scholar] [CrossRef]
- Bolton, M.D. Primary metabolism and plant defense-fuel for the fire. Mol. Plant-Microbe Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bago, B.; Pfeffer, P.E.; Abubaker, J.; Jun, J.; Allen, J.W.; Brouillette, J.; Douds, D.D.; Lammers, P.J.; Shachar-Hill, Y. Carbon export from arbuscular mycorrhizal roots involves the translocation of carbohydrate as well as lipid. Plant Physiol. 2003, 131, 1496–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechri, B.; Tekaya, M.; Cheheb, H.; Attia, F.; Hammami, M. Accumulation of flavonoids and phenolic compounds in olive tree roots in response to mycorrhizal colonization: A possible mechanism for regulation of defense molecules. J. Plant Physiol. 2015, 185, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Bączek, K.B.; Wiśniewska, M.; Przybył, J.L.; Kosakowska, O.; Węglarz, Z. Arbuscular mycorrhizal fungi in chamomile (Matricaria recutita L.) organic cultivation. Ind. Crops Prod. 2019, 140, 111562. [Google Scholar] [CrossRef]
- Hao, Z.; Charles, D.J.; Yu, L.; Simon, J.E. Purification and characterization of a phenylalanine ammonia-lyase from Ocimum basilicum. Phytochemistry 1996, 43, 735–739. [Google Scholar]
- Akiyama, K.; Matsuoka, H.; Hayashi, H. Isolation and identification of a phosphate deficiency-induced C-glycosyl flavonoid that stimulates arbuscular mycorrhiza formation in melon roots. Mol. Plant-Microbe Interact. 2002, 15, 334–340. [Google Scholar] [CrossRef]
- Abdallah, M.; Dubousset, L.; Meuriot, F.; Etienne, P.; Avice, J.C.; Ourry, A. Effect of mineral sulphur availability on nitrogen and sulphur uptake and remobilization during the vegetative growth of Brassica napus L. J. Exp. Bot. 2010, 61, 2635–2646. [Google Scholar] [CrossRef]
- Qin, M.; Zhang, Q.; Pan, J.; Jiang, S.; Liu, Y.; Bahadur, A.; Peng, Z.; Yang, Y.; Feng, H. Effect of arbuscular mycorrhizal fungi on soil enzyme activity is coupled with increased plant biomass. Eur. J. Soil Sci. 2020, 71, 84–92. [Google Scholar] [CrossRef]
- Al-Karaki, G.N. Effects of mycorrhizal fungi inoculation on green pepper yield and mineral uptake under irrigation with saline water. Adv. Plants Agric. Res. 2017, 6, 00231. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.D.; Zhang, D.J.; Hu, X.C.; Wu, Q.S.; Jiang, C.J.; Xia, T.J.; Gao, X.B.; Kuča, K. Mycorrhiza-induced changes in root growth and nutrient absorption of tea plants. Plant Soil Environ. 2018, 64, 283–289. [Google Scholar]
- Selvakumar, G.; Kim, K.; Hu, S.; Sa, T. Effect of salinity on plants and the role of arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria in alleviation of salt stress. In Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment; Springer: New York, NY, USA, 2014; pp. 115–144. [Google Scholar]
- Balliu, A.; Sallaku, G.; Rewald, B. AMF inoculation enhances growth and improves the nutrient uptake rates of transplanted, salt-stressed tomato seedlings. Sustainability 2015, 7, 15967–15981. [Google Scholar] [CrossRef] [Green Version]
- Symanczik, S.; Lehmann, M.F.; Wiemken, A.; Boller, T.; Courty, P.E. Effects of two contrasted arbuscular mycorrhizal fungal isolates on nutrient uptake by Sorghum bicolor under drought. Mycorrhiza 2018, 28, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.F.; Lei, H.C. Effect of daily light integral on plant growth and development . Acta Hortic. Sin. 2019, 46, 1670–1680. [Google Scholar]
- Zhai, J.; Gao, Y.; Zhang, X.W.; Han, L.J.; Bi, H.G.; Li, Q.M.; Ai, X.Z. Effects of silicon and calcium on photosynthesis, yield and quality of cucumber in solar-greenhouse. Acta Hortic. Sin. 2019, 46, 701–713. [Google Scholar]
- Etxeberria, E.; Gonzalez, P.; Fanton-Borges, A.; Brodersen, C. The use of laser light to enhance the uptake of foliar-applied substances into citrus (Citrus sinensis) leaves. Appl. Plant Sci. 2016, 4, 1500106. [Google Scholar] [CrossRef]
- Ni, Z.J.; Wang, X.; Shen, Y.; Thakur, K.; Han, J.; Zhang, J.G.; Hu, F.; Wei, Z.J. Recent updates on the chemistry, bioactivities, mode of action, and industrial applications of plant essential oils. Trends Food Sci. Technol. 2021, 110, 78–89. [Google Scholar] [CrossRef]
- Moyo, M.; Van Staden, J. Medicinal properties and conservation of Pelargonium sidoides DC. J. Ethnopharmacol. 2014, 152, 243–255. [Google Scholar] [CrossRef]
- Boukhatem, M.N.; Kameli, A.; Saidi, F. Essential oil of Algerian rose-scented geranium (Pelargonium graveolens): Chemical composition and antimicrobial activity against food spoilage pathogens. Food Control 2013, 34, 208–213. [Google Scholar] [CrossRef]
- Ahamad, J.; Uthirapathy, S. Chemical characterization and antidiabetic activity of essential oils from Pelargonium graveolens leaves. ARO—Sci. J. Koya Univ. 2021, 9, 109–113. [Google Scholar]
- Okla, M.K.; Abdel-Mawgoud, M.; Alamri, S.A.; Abbas, Z.K.; Al-Qahtani, W.H.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Hassan, A.H.; Selim, S.; Alruhaili, M.H.; et al. Developmental stages-specific response of Anise plants to laser-induced growth, nutrients accumulation, and essential oil metabolism. Plants 2021, 10, 2591. [Google Scholar] [CrossRef] [PubMed]
- Balkhyour, M.A.; Tammar, A.M.; Summan, A.S.; Hassan, A.H. Enhancing biomass and productivity of coumarins and essential oil in ajwain (Trachyspermum ammi) sprouts via laser light treatment. Ind. Crops Prod. 2021, 170, 113837. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Thomidis, T.; Lazari, D.; Panou-Filotheou, E.; Karagiannidou, C. Effect of three Greek arbuscular mycorrhizal fungi in improving the growth, nutrient concentration, and production of essential oils of oregano and mint plants. Sci. Hortic. 2011, 129, 329–334. [Google Scholar] [CrossRef]
- Rydlová, J.; Jelínková, M.; Dušek, K.; Dušková, E.; Vosátka, M.; Püschel, D. Arbuscular mycorrhiza differentially affects synthesis of essential oils in coriander and dill. Mycorrhiza 2016, 26, 123–131. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Deans, S.G.; Hart, S. Bioactivity of geranium oils from different commercial sources. J. Essent. Oil Res. 1996, 8, 281–290. [Google Scholar] [CrossRef]
- Pintore, G.; Usai, M.; Bradesi, P.; Juliano, C.; Boatto, G.; Tomi, F.; Chessa, M.; Cerri, R.; Casanova, J. Chemical composition and antimicrobial activity of Rosmarinus officinalis L. oils from Sardinia and Corsica. Flavour Fragr. J. 2002, 17, 15–19. [Google Scholar] [CrossRef]
- Laredo, J.V.; Abut, M.; Calvo-Torras, M.A. Antimicrobial activity of essences from Labiatae. Microbios 1995, 82, 171–172. [Google Scholar]
- Hethelyi, E.; Koczka, I.; Tetenyi, P. Phytochemical and antimicrobial analysis of essential oils. Herba Hung. 1989, 28, 99–115. [Google Scholar]
- Rosato, A.; Vitali, C.; De Laurentis, N.; Armenise, D.; Milillo, M.A. Antibacterial effect of some essential oils administered alone or in combination with Norfloxacin. Phytomedicine 2007, 14, 727–732. [Google Scholar] [CrossRef]
- Ghannadi, A.; Bagherinejad, M.R.; Abedi, D.; Jalali, M.; Absalan, B.; Sadeghi, N. Antibacterial activity and composition of essential oils from Pelargonium graveolens L’Her and Vitex agnus-castus L. Iran. J. Microbiol. 2012, 4, 171. [Google Scholar]
- Kordali, S.; Cakir, A.; Mavi, A.; Kilic, H.; Yildirim, A. Screening of chemical composition and antifungal and antioxidant activities of the essential oils from three Turkish Artemisia species. J. Agric. Food Chem. 2005, 53, 1408–1416. [Google Scholar] [CrossRef] [PubMed]
- Semeniuc, C.A.; Pop, C.R.; Rotar, A.M. Antibacterial activity and interactions of plant essential oil combinations against Gram-positive and Gram-negative bacteria. J. Food Drug Anal. 2017, 25, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavastin, D.E.; Bujor, A.; Tuchiluş, C.; Mircea, C.G.; Gherman, S.P.; Aprotosoaie, A.C. Studies on antioxidant, antihyperglycemic and antimicrobial effects of edible mushrooms Boletus edulis and Cantharellus cibarius. J. Plant Dev. 2016, 23, 87–95. [Google Scholar]
- Gwynne, P.J.; Gallagher, M.P. Light as a broad-spectrum antimicrobial. Front. Microbiol. 2018, 9, 119. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Andrade, G.; Mihara, K.L.; Linderman, R.G.; Bethlenfalvay, G.J. Bacteria from rhizosphere and hyphosphere soils of different arbuscular-mycorrhizal fungi. Plant Soil 1997, 192, 71–79. [Google Scholar] [CrossRef]
- Agusa, T.; Kunito, T.; Yasunaga, G.; Iwata, H.; Subramanian, A.; Ismail, A.; Tanabe, S. Concentrations of trace elements in marine fish and its risk assessment in Malaysia. Mar. Pollut. Bull. 2005, 51, 896–911. [Google Scholar] [CrossRef]
- Clark, J.M.; Switzer, R.L. Experimental Biochemistry, 2nd ed.; Freeman and Company: San Francisco, CA, USA, 1977; p. 180. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; The Association: Arlington, VA, USA, 1990. [Google Scholar]
- Hassanpour, H.; Khavari-Nejad, R.A.; Niknam, V.; Razavi, K.; Najafi, F. Effect of penconazole and drought stress on the essential oil composition and gene expression of Mentha pulegium L. (Lamiaceae) at flowering stage. Acta Physiol. Plant. 2014, 36, 1167–1175. [Google Scholar] [CrossRef]
- Rubnawaz, S.; Okla, M.K.; Akhtar, N.; Khan, I.U.; Bhatti, M.Z.; Duong, H.Q.; El-Tayeb, M.A.; Elbadawi, Y.B.; Almaary, K.S.; Moussa, I.M.; et al. Antibacterial, antihemolytic, cytotoxic, anticancer, and antileishmanial effects of Ajuga bracteosa transgenic plants. Plants 2021, 10, 1894. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Growth Parameters | No Irradiation | Laser Irradiation | ||
|---|---|---|---|---|
| Control | Mycorrhizal | Control | Mycorrhizal | |
| Colonization (% root) | 0 ± 0 a | 38.7 ± 3.1 b | 0 ± 0 a | 49.3 ± 3.7 c |
| Hyphal length (cm/g soil) | 0 ± 0 a | 13.1 ± 1.4 b | 0 ± 0 a | 19.4 ± 0.7 c |
| Number of arbuscules/cm root | 0 ± 0 a | 5.2 ± 0.2 b | 0 ± 0 a | 6.8 ± 0.3 c |
| Metabolites (mg/g DW) | No Irradiation | Laser Irradiation | ||
|---|---|---|---|---|
| Control | Mycorrhizal | Control | Mycorrhizal | |
| Total protein | 148.68 ± 8 a | 158.3 ± 9 b | 163.5 ± 11.8 b | 186.5 ± 6.5 c |
| Total sugar | 59.68 ± 2.6 a | 61.7 ± 3.6 a | 64.1 ± 3.9 a | 76.4 ± 3.1 b |
| Total phenols | 11.07 ± 1.2 a | 14.5 ± 1 b | 17.5 ± 1 a | 21.5 ± 1 c |
| Total flavonoids | 0.53 ± 0.1 a | 0.8 ± 0.1 b | 0.8 ± 0.1 b | 1.2 ± 0.1 c |
| Total alkaloid | 18.5 ± 1.6 a | 26.3 ± 2 b | 19.4 ± 2.4 a | 24.1 ± 2.9 b |
| Minerals (mg/g DW) | No Irradiation | Laser Irradiation | ||
|---|---|---|---|---|
| Control | Mycorrhizal | Control | Mycorrhizal | |
| K | 12.2 ± 0.7 a | 17.7 ± 0.5 b | 16.4 ± 0.8 b | 26.9 ± 0.7 c |
| P | 4.57 ± 0.4 a | 6.67 ± 0.3 b | 5.89 ± 0.6 a | 8.16 ± 0.3 c |
| Ca | 2.32 ± 0.3 a | 3.4 ± 0.2 ab | 2.95 ± 0.2 a | 4.3 ± 0.3 b |
| Mg | 1.81 ± 0.3 a | 2.7 ± 0.2 ab | 2 ± 0.3 a | 3.47 ± 0.3 b |
| Na | 0.39 ± 0.1 a | 0.61 ± 0.1 b | 0.53 ± 0.1 ab | 0.65 ± 0.1 b |
| Fe | 0.15 ± 0 a | 0.23 ± 0 ab | 0.21 ± 0 a | 0.27 ± 0 bc |
| Zn | 0.04 ± 0 a | 0.06 ± 0 a | 0.06 ± 0 a | 0.05 ± 0 a |
| Parameters | No Irradiation | Laser Irradiation | ||
|---|---|---|---|---|
| Control | Mycorrhizal | Control | Mycorrhizal | |
| Oil yield (%) | 12.8 ± 0.3 a | 12.7 ± 0.5 a | 13.4 ± 0.5 a | 16.8 ± 0.4 b |
| Essential oil (% dry weight) | 3.1 ± 0.1 a | 3.48 ± 0.2 a | 3.8 ± 0.2 a | 4.6 ± 0.1 a |
| Total essential oil (%) | ||||
| Beta-Citronellol | 16.4 ± 0.1 a | 17.1 ± 0.1 a | 18.3 ± 0.1 b | 20.4 ± 0.1 b |
| Citronellyl formate | 7.3 ± 0.6 a | 7.8 ± 0.8 a | 8.9 ± 0.9 a | 9.3 ± 0.7 b |
| Citronellyl propionate | 0.09 ± 0 a | 0.08 ± 0 a | 0.09 ± 0 a | 0.14 ± 0 b |
| Geraniol | 20.1 ± 0.8 a | 27.7 ± 1 b | 22.6 ± 1.1 b | 34.8 ± 0.9 c |
| Geraniol formate | 7.05 ± 0.7 a | 8.73 ± 1 a | 7.76 ± 1 a | 9.8 ± 0.8 b |
| α-pinene | 1.04 ± 0.1 a | 1.68 ± 0.1 a | 1.68 ± 0.1 a | 1.7 ± 0.1 a |
| Limonene | 0.17 ± 0 a | 0.16 ± 0 a | 0.15 ± 0 a | 0.26 ± 0 b |
| Myrcene | 0.33 ± 0 a | 0.19 ± 0 a | 0.19 ± 0 a | 0.18 ± 0 a |
| Linalool | 4.04 ± 0.1 a | 4.99 ± 0.2 b | 4.17 ± 0.2 a | 5.33 ± 0.1 b |
| Cis-β-ocimene | 0.24 ± 0.1 a | 0.27 ± 0.1 a | 0.25 ± 0.1 a | 0.31 ± 0.1 a |
| Sabinene | 0.3 ± 0 a | 0.5 ± 0 b | 0.27 ± 0 a | 0.48 ± 0 b |
| p-cymene | 0.57 ± 0 a | 0.52 ± 0 a | 0.54 ± 0 a | 0.8 ± 0 b |
| Beta-phellandrene | 0.7 ± 0.1 a | 0.75 ± 0.1 a | 0.95 ± 0.1 b | 0.91 ± 0.1 b |
| I-Menthol | 0.04 ± 0.1 a | 0.13 ± 0.2 b | 0.04 ± 0.2 a | 0.08 ± 0.1 b |
| I-Menthone | 4.55 ± 0.2 a | 5.12 ± 0.2 a | 5.36 ± 0.2 a | 5.9 ± 0.2 a |
| Nerol | 3.66 ± 0.2 a | 4.18 ± 0.3 a | 4.31 ± 0.3 a | 5.91 ± 0.3 b |
| Beta-bourbonene | 0.75 ± 0.4 a | 1.02 ± 0.5 a | 0.82 ± 0.5 a | 0.93 ± 0.4 a |
| Beta-cubebene | 0.94 ± 0.2 a | 1.11 ± 0.3 a | 1.55 ± 0.3 ab | 2.6 ± 0.2 b |
| z-citral | 0.04 ± 0 a | 0.05 ± 0 a | 0.05 ± 0 a | 0.06 ± 0 a |
| α-Terpinene | 0.04 ± 0 a | 0.07 ± 0 b | 0.04 ± 0 a | 0.085 ± 0 b |
| Trans-caryophyllene | 1.67 ± 0 a | 1.84 ± 0 a | 1.97 ± 0 a | 1.97 ± 0 a |
| Isoledene | 1.4 ± 0 a | 2.55 ± 0 b | 1.65 ± 0 b | 2.95 ± 0 b |
| Germacrene-D | 3.83 ± 0 a | 3.48 ± 0 a | 3.71 ± 0 a | 5.3 ± 0 b |
| Ledene | 3.19 ± 0 a | 3.31 ± 0 a | 3.53 ± 0 a | 3.97 ± 0 a |
| δ-cadinene | 1.83 ± 0 a | 1.52 ± 0 a | 1.61 ± 0 a | 2.7 ± 0 b |
| Neryl acetate | 1.58 ± 0 a | 2.75 ± 0.1 b | 1.64 ± 0.1 a | 2.97 ± 0 b |
| Geranyl tiglate | 1.77 ± 0.1 a | 1.98 ± 0.1 a | 3.15 ± 0.1 b | 3.37 ± 0.1 b |
| Mintsulfide | 2.5 ± 0.7 a | 4.62 ± 0.9 b | 4.46 ± 1 b | 6.12 ± 0.8 c |
| Essential oil precursors | ||||
| Phenylalanine | 3.37 ± 0.2 a | 3.43 ± 0.3 a | 3.53 ± 0.3 a | 5.41 ± 0.2 b |
| Cinnamic acid | 4.55 ± 0.4 a | 7.36 ± 0.5 b | 3.32 ± 0.6 a | 7.4 ± 0.5 b |
| Shikimic acid | 72.9 ± 2.5 a | 87.6 ± 3.4 b | 81.21 ± 3.6 a | 90.4 ± 1.4 b |
| Bacterial Specie | No Irradiation | Laser Irradiation | ||
|---|---|---|---|---|
| Control | Mycorrhizal | Control | Mycorrhizal | |
| Zones of Inhibition (mm) | ||||
| Bacillus subtilis | 13.2 ± 0.6 a | 21 ± 1.3 b | 18.1 ± 0.8 b | 25.1 ± 1 c |
| Streptococcussalivarius | 21.8 ± 0.4 a | 30 ± 1.4 b | 23.54 ± 1 a | 32.1 ± 0.9 b |
| Escherichia coli | 18.6 ± 0.6 a | 25 ± 1.8 b | 16.05 ± 0.8 a | 35.31 ± 1.2 c |
| Pseudomonas aeruginosa | 19.3 ± 1 a | 29 ± 1.1 b | 17.98 ± 0.9 a | 23.54 ± 1 a |
| Sarcina lutea | 14.7 ± 0.5 a | 23 ± 1.2 b | 22.47 ± 1.2 b | 31.03 ± 0.9 c |
| Staphylococcus aureus | 15.8 ± 1.2 a | 22 ± 1 b | 14.98 ± 1.8 a | 23.54 ± 1.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okla, M.K.; Rubnawaz, S.; Dawoud, T.M.; Al-Amri, S.; El-Tayeb, M.A.; Abdel-Maksoud, M.A.; Akhtar, N.; Zrig, A.; AbdElgayed, G.; AbdElgawad, H. Laser Light Treatment Improves the Mineral Composition, Essential Oil Production and Antimicrobial Activity of Mycorrhizal Treated Pelargoniumgraveolens. Molecules 2022, 27, 1752. https://doi.org/10.3390/molecules27061752
Okla MK, Rubnawaz S, Dawoud TM, Al-Amri S, El-Tayeb MA, Abdel-Maksoud MA, Akhtar N, Zrig A, AbdElgayed G, AbdElgawad H. Laser Light Treatment Improves the Mineral Composition, Essential Oil Production and Antimicrobial Activity of Mycorrhizal Treated Pelargoniumgraveolens. Molecules. 2022; 27(6):1752. https://doi.org/10.3390/molecules27061752
Chicago/Turabian StyleOkla, Mohammad K., Samina Rubnawaz, Turki M. Dawoud, Saud Al-Amri, Mohamed A. El-Tayeb, Mostafa A. Abdel-Maksoud, Nosheen Akhtar, Ahlem Zrig, Gehad AbdElgayed, and Hamada AbdElgawad. 2022. "Laser Light Treatment Improves the Mineral Composition, Essential Oil Production and Antimicrobial Activity of Mycorrhizal Treated Pelargoniumgraveolens" Molecules 27, no. 6: 1752. https://doi.org/10.3390/molecules27061752