
The In Vitro Anti-Cancer Activities and Mechanisms of Action of 9-Methoxycanthin-6-one from Eurycoma longifolia in Selected Cancer Cell Lines

Abstract
:1. Introduction
2. Results
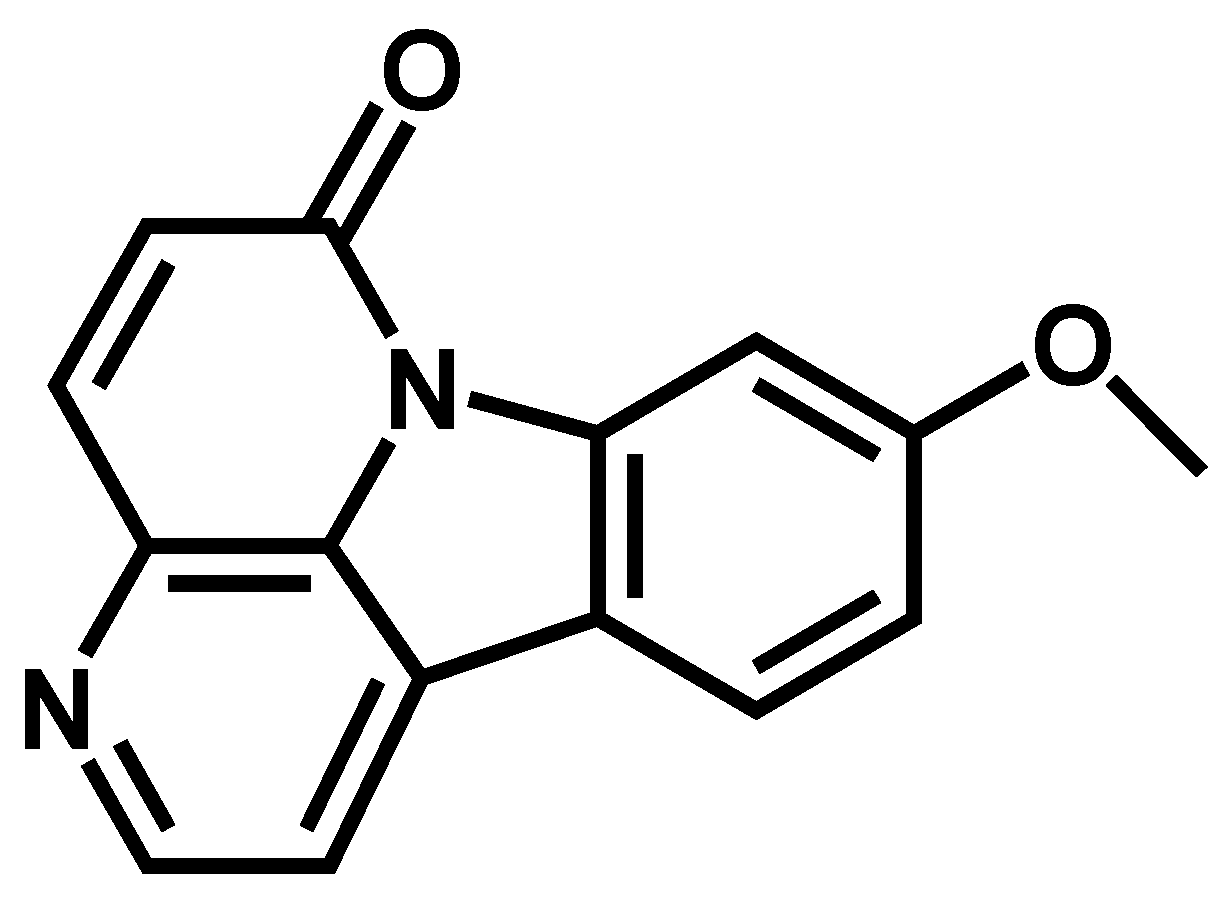
2.1. Identification of 9-Methoxycanthin-6-one
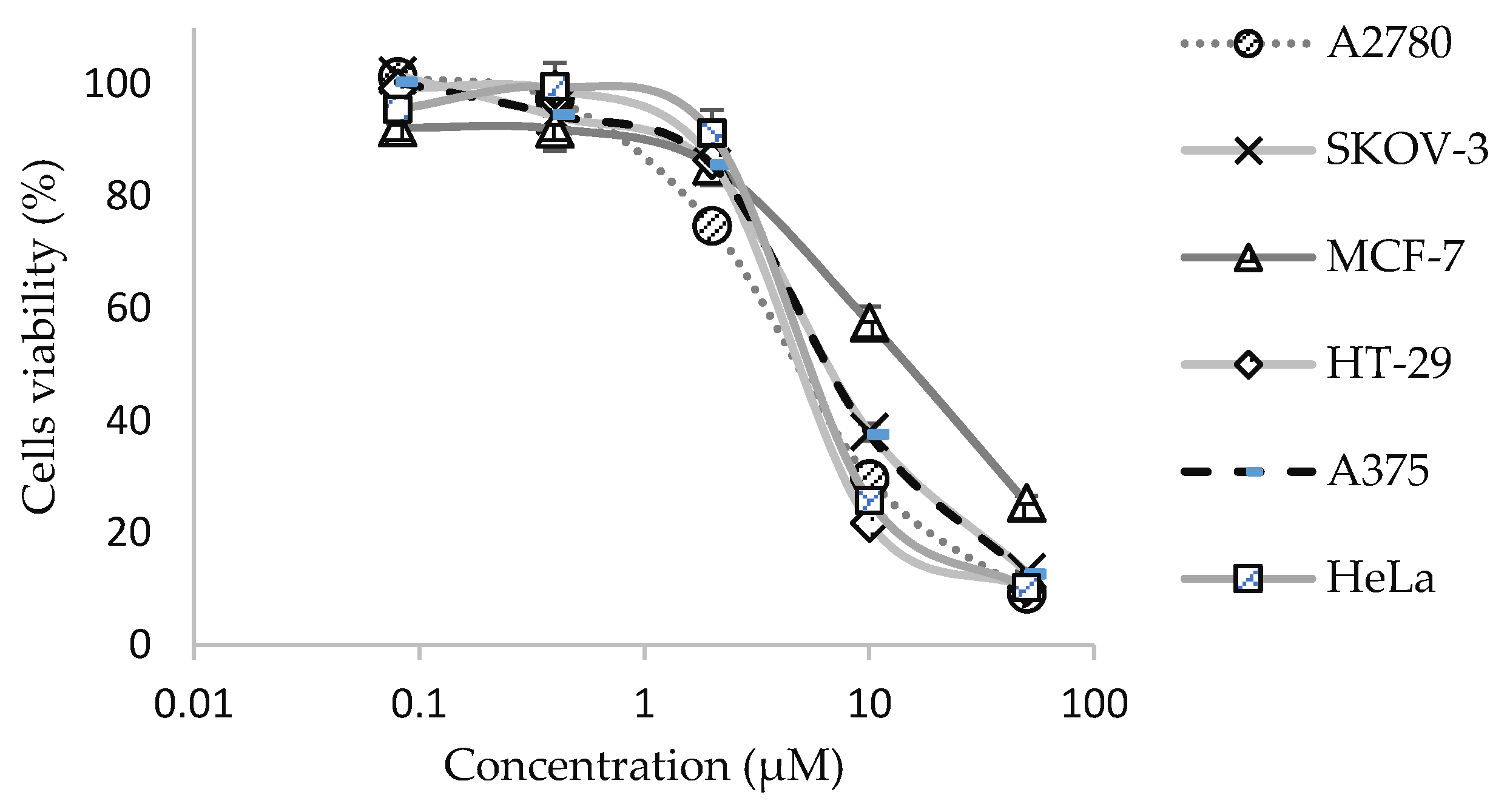
2.2. In Vitro Anti-Cancer Activities of 9-Methoxycanthine-6-one on Selected Cancer Cell Lines
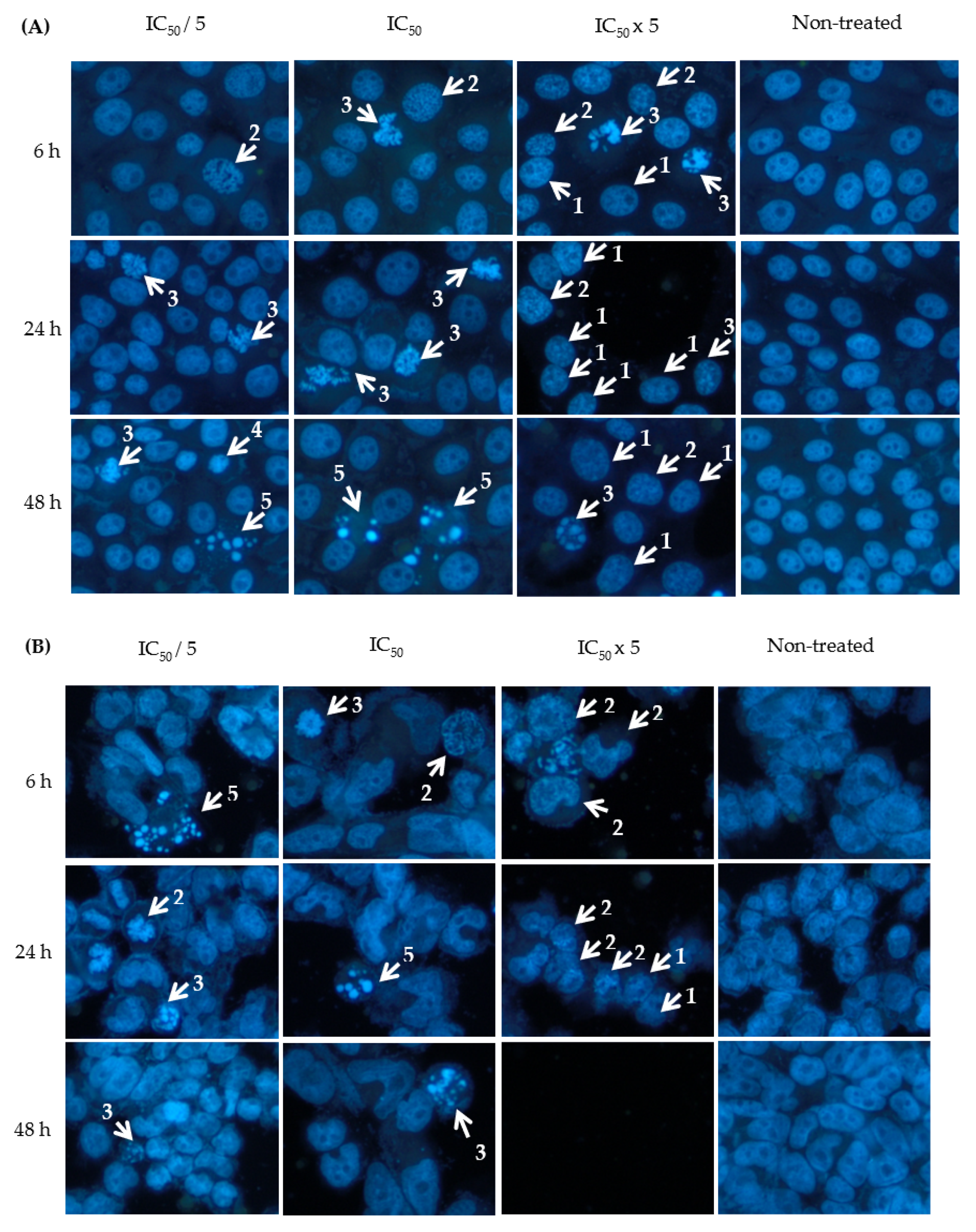
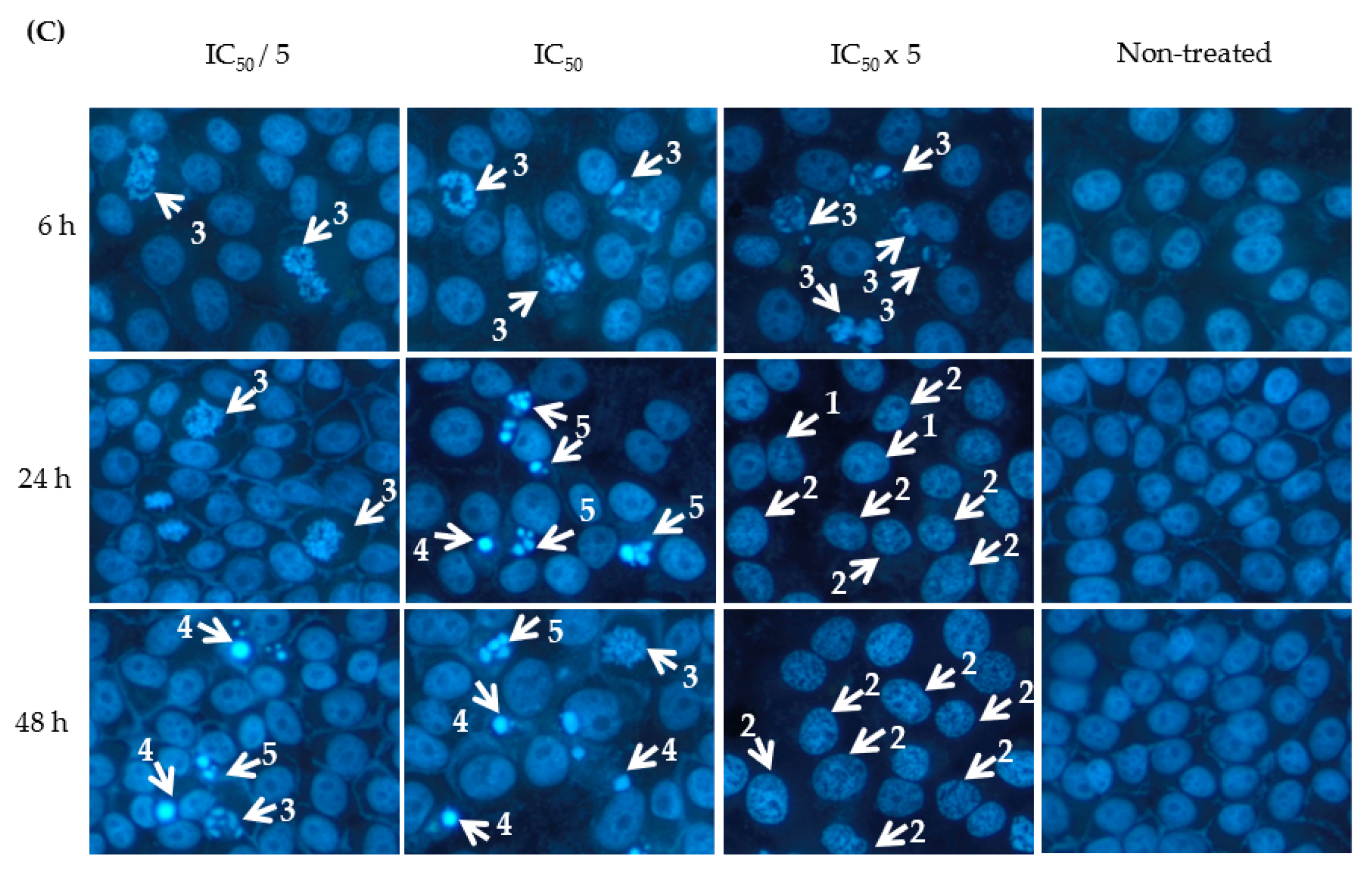
2.3. Effect of 9-Methoxycanthin-6-one on Apoptosis
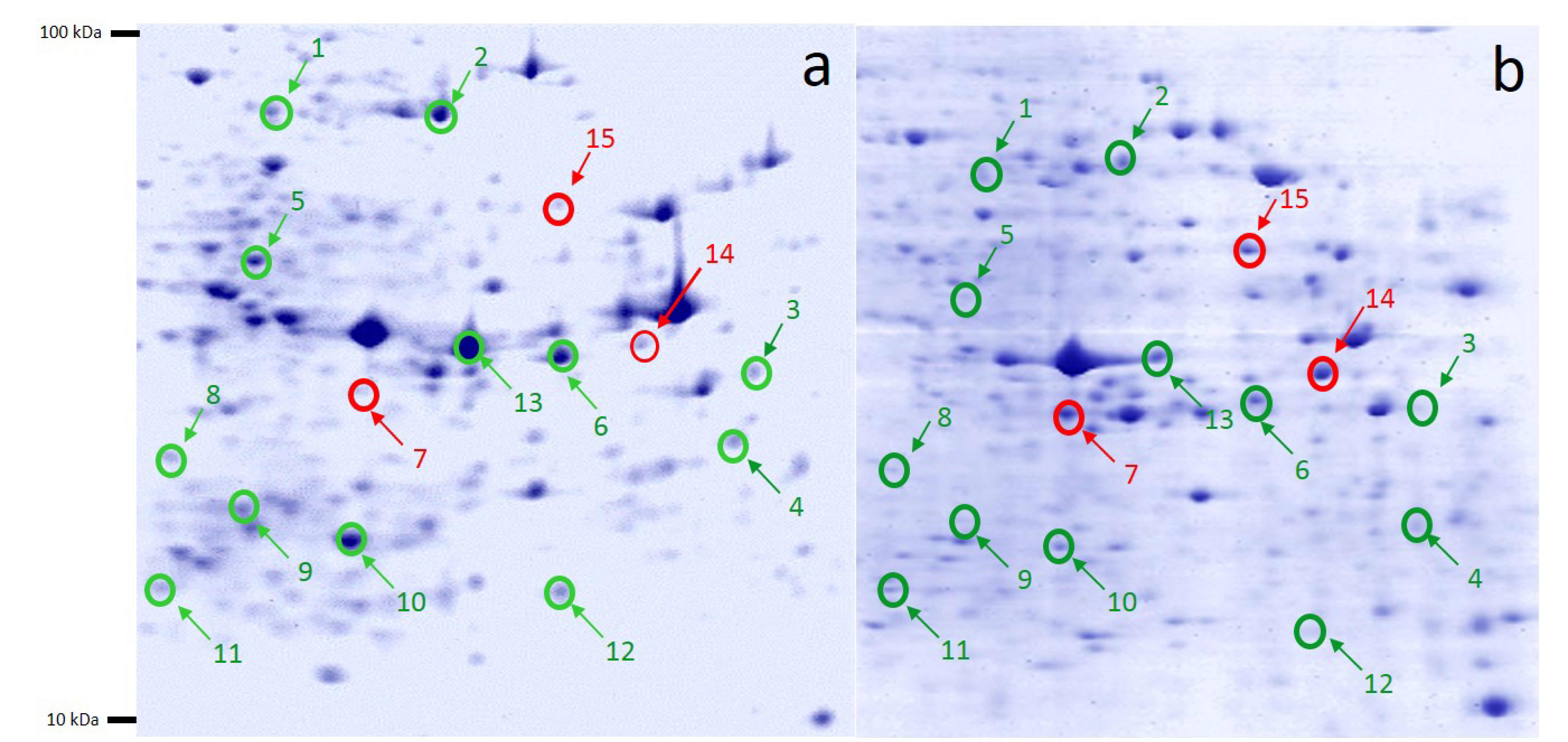
2.4. 2-DE Analysis of 9-Methoxycanthin-6-one-treated and Non-Treated A2780 Ovarian Cancer Cells
2.5. Protein Identifications
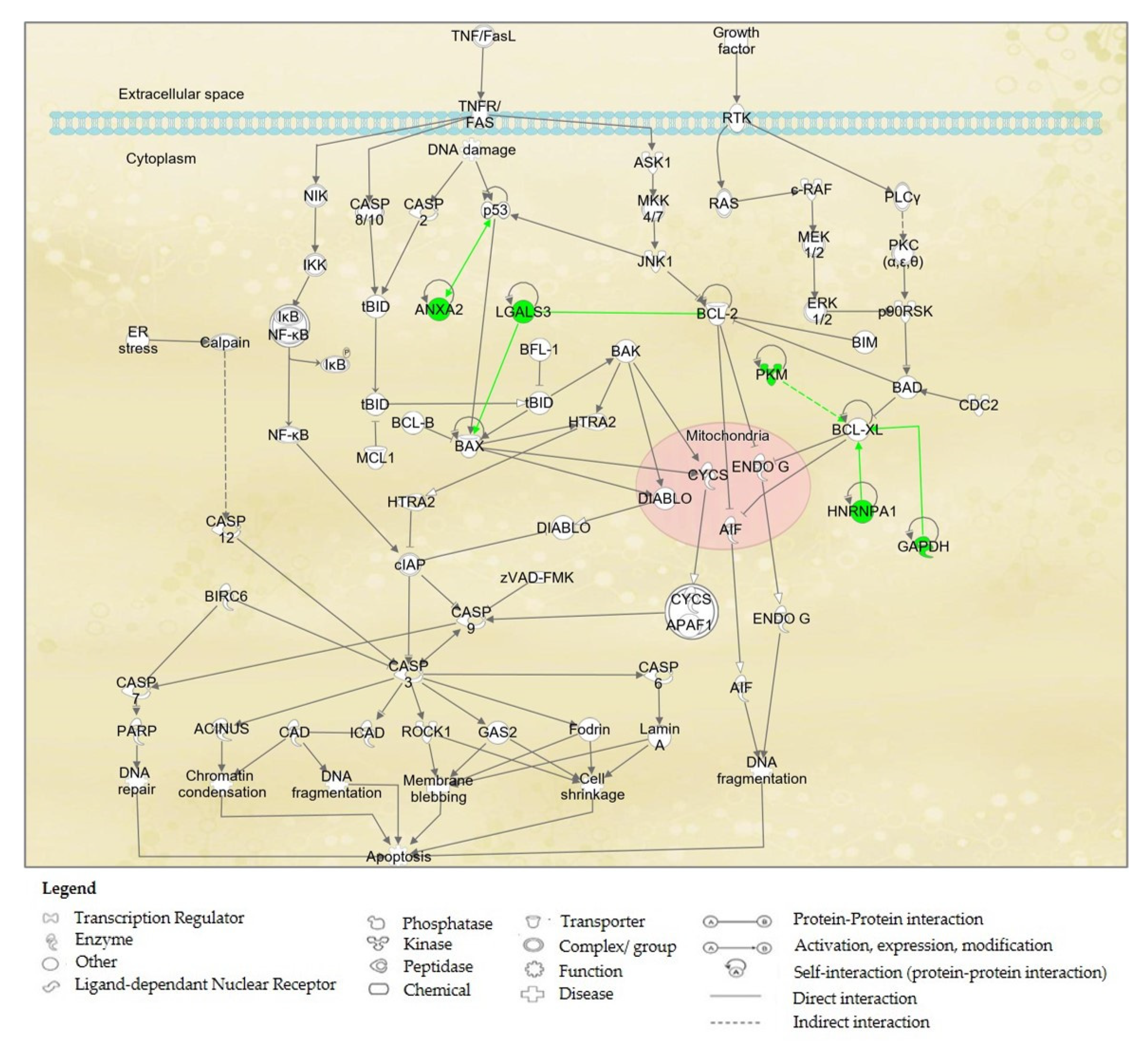
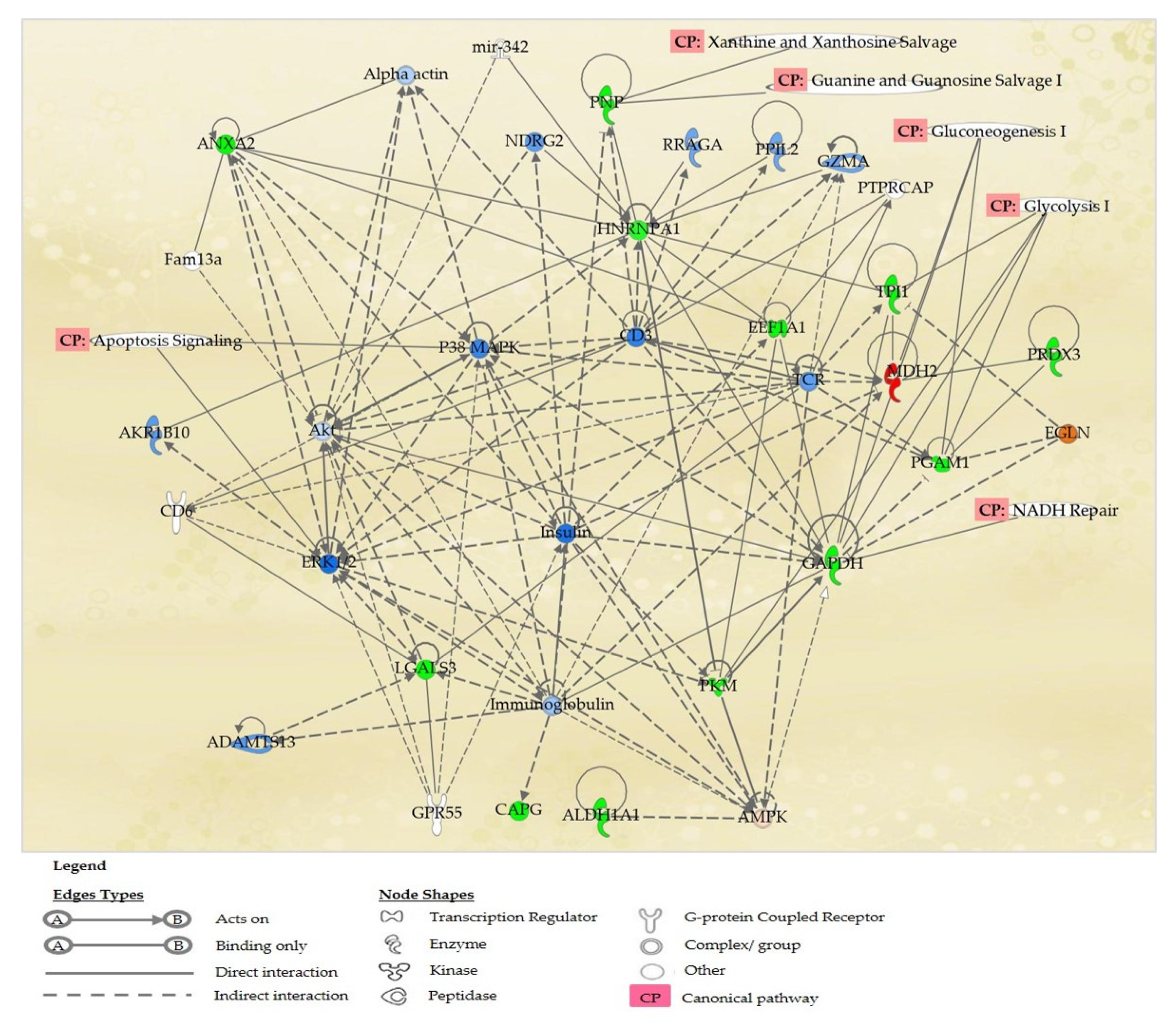
2.6. Proteins’ Interaction Analysis
3. Discussion
4. Materials and Methods
4.1. Production of E. longifolia Hairy Roots Culture in the Bioreactor
4.2. Isolation of 9-Methoxycanthin-6-one from E. longifolia Hairy Root Culture
4.3. Cell Culture and Treatments
4.4. Cells Viability Assay
4.5. Apoptotic Hoechst 33342 Assay
4.6. Two-Dimensional Gel Electrophoresis (2-DE)
4.7. Image and Data Analysis
4.8. In-Gel Tryptic Digestion
4.9. Mass Spectrometry and Database Search
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Pilleron, S.; Sarfati, D.; Janssen-Heijnen, M.; Vignat, J.; Ferlay, J.; Bray, F.; Soerjomataram, I. Global cancer incidence in older adults, 2012 and 2035: A population-based study. Int. J. Cancer 2019, 144, 49–58. [Google Scholar] [CrossRef]
- Rehman, S.U.; Choe, K.; Yoo, H.H. Review on a traditional herbal medicine, Eurycoma longifolia Jack (Tongkat Ali): Its traditional uses, chemistry, evidence-based pharmacology and toxicology. Molecules 2016, 21, 331. [Google Scholar] [CrossRef] [Green Version]
- Tamizhazhagan, V.; Pugazhendy, K. Social and economic burden of cancer on 2020—Mini review. J. Biol. Med. Sci. 2017, 1, 2. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Call, J.A.; Eckhardt, S.G.; Camidge, D.R. Targeted manipulation of apoptosis in cancer treatment. Lancet Oncol. 2008, 9, 1002–1011. [Google Scholar] [CrossRef]
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula; Ministry of Agriculture and Co-operations: Kuala Lumpur, Malaysia, 1966; Volume 1(A–H).
- Thoi, L.V.; Suong, N.N. Constituents of Eurycoma longifolia. J. Org. Chem. 1970, 35, 1104–1109. [Google Scholar] [CrossRef]
- Perry, L.M.; Metzger, J. Medicinal Plants of East and Southeast Asia: Attributed Properties and Uses; M.I.T. Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Bhat, R.; Karim, A.A. Tongkat Ali (Eurycoma longifolia Jack): A review on its ethnobotany and pharmacological importance. Fitoterapia 2010, 81, 669–679. [Google Scholar] [CrossRef]
- Kuo, P.C.; Damu, A.G.; Lee, K.H.; Wu, T.S. Cytotoxic and antimalarial constituents from the roots of Eurycoma longifolia. Bioorg. Med. Chem. 2004, 12, 537–544. [Google Scholar] [CrossRef]
- Ulbricht, C.; Conquer, J.; Flanagan, K.; Isaac, R.; Rusie, E.; Windsor, R.C. An evidence-based systematic review of Tongkat Ali (Eurycoma longifolia) by the natural standard research collaboration. J. Diet. Suppl. 2013, 10, 54–83. [Google Scholar] [CrossRef]
- Guo, Z.; Vangapandu, S.; Sindelar, R.W.; Walker, L.A.; Sindelar, R.D. Biologically active quassinoids and their chemistry: Potential leads for drug design. Curr. Med. Chem. 2005, 12, 173–190. [Google Scholar] [CrossRef]
- Kardono, L.B.S.; Angerhofer, C.K.; Tsauri, S.; Padmawinata, K.; Pezzuto, J.M.; Douglas Kinghorn, A. Cytotoxic and antimalarial constituents of the roots of Eurycoma longifolia. J. Nat. Prod. 1991, 54, 1360–1367. [Google Scholar] [CrossRef]
- Itokawa, H.; Kishi, E.; Morita, H.; Takeya, K. Cytotoxic quassinoids and tirucallane-type triterpenes from the woods of Eurycoma longifolia. Chem. Pharm. Bull. 1992, 40, 1053–1055. [Google Scholar] [CrossRef] [Green Version]
- Miyake, K.; Li, F.; Tezuka, Y.; Awale, S.; Kadota, S. Cytotoxic activity of quassinoids from Eurycoma longifolia. Nat. Prod. Commun. 2010, 5, 1009–1012. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.C.; Shi, L.S.; Damu, A.G.; Su, C.R.; Huang, C.H.; Ke, C.H.; Wu, J.-B.; Lin, A.J.; Bastow, K.F.; Lee, K.H.; et al. Cytotoxic and antimalarial β-carboline alkaloids from the roots of Eurycoma longifolia. J. Nat. Prod. 2003, 66, 1324–1327. [Google Scholar] [CrossRef]
- Nurhanan, M.Y.; Hawariah, L.P.A.; Ilham, A.M.; Shukri, M.A.M. Cytotoxic effects of the root extracts of Eurycoma longifolia Jack. Phyther. Res. 2005, 19, 994–996. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Thu, H.E.; Hussain, Z.; Mohamed, I.N.; Shuid, A.N. Eurycoma longifolia, a potential phytomedicine for the treatment of cancer: Evidence of p53-mediated apoptosis in cancerous cells. Curr. Drug Targets 2017, 19, 1109–1126. [Google Scholar] [CrossRef]
- Mitsunaga, K.; Koike, K.; Tanaka, T.; Ohkawa, Y.; Kobayashi, Y.; Sawaguchi, T.; Ohmoto, T. Canthin-6-one alkaloids from Eurycoma longifolia. Phytochemistry 1994, 35, 799–802. [Google Scholar] [CrossRef]
- Jiang, M.-X.; Zhou, Y.-J. Canthin-6-one alkaloids from Picrasma quassioides and their cytotoxic activity. J. Asian Nat. Prod. Res. 2008, 10, 1009–1012. [Google Scholar] [CrossRef]
- Choo, C.Y.; Chan, K.L. High performance liquid chromatography analysis of canthinone alkaloids from Eurycoma longifolia. Planta Med. 2002, 68, 382–384. [Google Scholar] [CrossRef]
- Mahmood, M.; Normi, R.; Sybramaniam, S. Distribution of 9-methoxycanthin-6-one from the intact plant parts and callus cultures of “Eurycoma longifolia” (Tongkat Ali). Aust. J. Crop Sci. 2011, 5, 1565–1569. [Google Scholar]
- Ling, A.P.K.; Phua, G.A.T.; Tee, C.S.; Hussein, S. Optimization of protoplast isolation protocols from callus of Eurycoma longifolia. J. Med. Plants Res. 2010, 4, 1778–1785. [Google Scholar] [CrossRef]
- Nazirah, A.; Nor-Hasnida, H.; Ismanizan, I.; Norlia, B.; Abdul-Rashih, A.; Muhammad-Fuad, Y.; Mohd-Saifullah, A.W. Production of 9-methoxycanthin-6-one in elicited Eurycoma longifolia. J. Trop. For. Sci. 2018, 30, 606–614. [Google Scholar]
- Roja, G.; Rao, P.S. Anticancer compounds from tissue cultures of medicinal plants. J. Herbs Spices Med. Plants 2008, 7, 71–102. [Google Scholar] [CrossRef]
- Lorence, A.; Medina-Bolivar, F.; Nessler, C.L. Camptothecin and 10-hydroxycamptothecin from Camptotheca acuminata hairy roots. Plant Cell Rep. 2004, 22, 437–441. [Google Scholar] [CrossRef]
- Fett-Neto, A.G.; Dicosmo, F.; Reynolds, W.F.; Sakata, K. Cell culture of taxus as a source of the antineoplastic drug taxol and related taxanes. Bio/Technology 1992, 10, 1572–1575. [Google Scholar] [CrossRef]
- Boik, J.C.; Newman, R.A. Structure-activity models of oral clearance, cytotoxicity, and LD50: A screen for promising anticancer compounds. BMC Pharmacol. 2008, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.J.; Bao, J.L.; Chen, X.P.; Huang, M.; Wang, Y.T. Alkaloids isolated from natural herbs as the anticancer agents. Evid.-Based Complement. Altern. Med. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Crowley, L.C.; Marfell, B.J.; Waterhouse, N.J. Analyzing cell death by nuclear staining with Hoechst 33342. Cold Spring Harb. Protoc. 2016, 9, pdb-rot087205. [Google Scholar] [CrossRef]
- Sanz, G.; Singh, M.; Peuget, S.; Selivanova, G. Inhibition of p53 inhibitors: Progress, challenges and perspectives. J. Mol. Cell Biol. 2019, 11, 586–599. [Google Scholar] [CrossRef] [Green Version]
- Pogmore, J.P.; Uehling, D.; Andrews, D.W. Pharmacological targeting of executioner proteins: Controlling life and death. J. Med. Chem. 2021, 64, 5276–5290. [Google Scholar] [CrossRef]
- Gutschner, T.; Diederichs, S. The hallmarks of cancer: A long non-coding RNA point of view. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, M.V.; Høgdall, C.K.; Umsen, K.M.J.; Høgdall, E.V.S. Annexin A2 and cancer: A systematic review. Int. J. Oncol. 2018, 52, 5–18. [Google Scholar] [CrossRef]
- Xu, X.H.; Pan, W.; Kang, L.H.; Feng, H.; Song, Y.Q. Association of Annexin A2 with cancer development (review). Oncol. Rep. 2015, 33, 2121–2128. [Google Scholar] [CrossRef] [Green Version]
- Blagih, J.; Buck, M.D.; Vousden, K.H. P53, Cancer and the immune response. J. Cell Sci. 2020, 133, jcs237453. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Wang, S.; Liu, J. Overexpression of tumor protein p53-regulated apoptosis-inducing protein 1 regulates proliferation and apoptosis of breast cancer cells through the PI3K/Akt pathway. J. Breast Cancer 2019, 22, 172–184. [Google Scholar] [CrossRef]
- Huang, Y.; Jin, Y.; Yan, C.H.; Yu, Y.; Bai, J.; Chen, F.; Zhao, Y.Z.; Fu, S.B. Involvement of Annexin A2 in p53 induced apoptosis in lung cancer. Mol. Cell. Biochem. 2008, 309, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, P.P. Galectin 3 as a guardian of the tumor microenvironment. Biochim. Biophys. Acta-Mol. Cell Res. 2016, 1863, 427–437. [Google Scholar] [CrossRef]
- Jia, W.; Kidoya, H.; Yamakawa, D.; Naito, H.; Takakura, N. Galectin-3 accelerates M2 macrophage infiltration and angiogenesis in tumors. Am. J. Pathol. 2013, 182, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Luna-Vargas, M.P.A.; Chipuk, J.E. Physiological and pharmacological control of BAK, BAX, and beyond. Trends Cell Biol. 2016, 26, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.F.; Fairlie, W.D. The Structural Biology of Bcl-x L. Int. J. Mol. Sci. 2019, 20, 1–18. [Google Scholar]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Lewis, E.M.; Wilkinson, A.S.; Davis, N.Y.; Horita, D.A.; Wilkinson, J.C. Nondegradative ubiquitination of apoptosis inducing factor (AIF) by x-linked inhibitor of apoptosis at a residue critical for AIF-mediated chromatin degradation. Biochemistry 2011, 50, 11084–11096. [Google Scholar] [CrossRef] [Green Version]
- Antone, P.D. Energy metabolism in cancer cells: How to explain the Warburg and Crabtree effects? Med. Hypotheses 2012, 79, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome C-mediated apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef]
- Moreno-Sánchez, R.; Marín-Hernández, A.; Saavedra, E.; Pardo, J.P.; Ralph, S.J.; Rodríguez-Enríquez, S. Who controls the ATP supply in cancer cells? Biochemistry lessons to understand cancer energy metabolism. Int. J. Biochem. Cell Biol. 2014, 50, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dong, C. Gluconeogenesis in cancer: Function and regulation of PEPCK, FBPase, and G6Pase. Trends Cancer 2019, 5, 30–45. [Google Scholar] [CrossRef]
- Grasmann, G.; Smolle, E.; Olschewski, H.; Leithner, K. Gluconeogenesis in cancer cells—Repurposing of a starvation-induced metabolic pathway? Biochim. Biophys. Acta-Rev. Cancer 2019, 1872, 24–36. [Google Scholar] [CrossRef]
- Miyamoto, T.; Amrein, H. Gluconeogenesis: An ancient biochemical pathway with a new twist. Fly 2017, 11, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Akram, M. Mini-review on glycolysis and cancer. J. Cancer Educ. 2013, 28, 454–457. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; Mcmahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Nurhanan, M.; Nor Azah, M.; Zunoliza, A.; Siti Humeriah, A.; Siti Syarifah, M.; Nor Hayati, A. In vitro anticancer activity and high-performance liquid chromatography profiles of Aquilaria subintegra fruit and seed extracts. J. Trop. For. Sci. 2017, 29, 208–214. [Google Scholar]
- Mahbub, A.A.; Le Maitre, C.L.; Haywood-Small, S.L.; McDougall, G.J.; Cross, N.A.; Jordan-Mahy, N. Differential effects of polyphenols on proliferation and apoptosis in human myeloid and lymphoid leukemia cell lines. Anti-Cancer Agents 2013, 13, 1601–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bringans, S.; Eriksen, S.; Kendrick, T.; Gopalakrishnakone, P.; Livk, A.; Lock, R.; Lipscombe, R. Proteomic analysis of the venom of Heterometrus longimanus (Asian black scorpion). Proteomics 2008, 8, 1081–1096. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | 1H δ ppm | 13C δ ppm |
|---|---|---|
| 1 | 8.23 (1H, d) | 115.660 |
| 2 | 8.78 (1H, d) | 143.810 |
| 3 | - | - |
| 4 | 7.98 (1H, d) | 138.053 |
| 5 | 6.99 (1H, d) | 129.685 |
| 6 | - | 159.376 |
| 7 | - | - |
| 8 | 7.11 (1H, d) | 101.448 |
| 9 | - | 163.411 |
| 10 | 7.91 (1H, d) | 114.756 |
| 11 | 8.12 (1H, d) | 123.949 |
| 12 | - | 116.775 |
| 13 | - | 142.084 |
| 14 | - | 129.691 |
| 15 | - | 131.150 |
| 16 | - | 132.362 |
| OCH3 | 4.04 (3H, s) | 56.200 |
| Compound | A2780 | SKOV-3 | MCF-7 | HT-29 | A375 | HeLa | H9c2 |
|---|---|---|---|---|---|---|---|
| EL50 | 4.04 ± 0.36 | 5.80 ± 0.40 | 15.09 ± 0.99 | 3.79 ± 0.069 | 5.71 ± 0.20 | 4.30 ± 0.27 | 37.34 ± 0.91 |
| Cis | 1.77 ± 0.018 | 2.27 ± 0.10 | 3.58 ± 0.14 | 1.38 ± 0.037 | 2.42 ± 0.19 | 1.54 ± 0.12 | 25.27 ± 0.77 |
| Tax | 0.018 ± 0.00011 | 0.38 ± 0.012 | >0.5 | 0.35 ± 0.0040 | 0.37 ± 0.0055 | 0.31 ± 0.0078 | >0.5 |
| Cell Line | 9-Methoxycanthin-6-one (µM) | Apoptotic Index (%) | ||
|---|---|---|---|---|
| 6 h | 24 h | 48 h | ||
| HT-29 | Non-treated | 1.76 ± 0.36 | 1.68 ± 0.38 | 1.81 ± 0.32 |
| IC50/5 | 3.36 ± 0.76 b | 3.88 ± 1.04 b | 4.14 ± 0.25 c | |
| IC50 | 4.23 ± 0.35 c | 4.56 ± 0.31 c | 5.31 ± 0.76 d | |
| IC50x5 | 100.00 ± 0.00 d | 100.00 ± 0.00 d | 100.00 ± 0.00 d | |
| A2780 | Non-treated | 2.26 ± 0.39 | 2.62 ± 0.61 | 2.71 ± 0.11 |
| IC50/5 | 3.85 ± 0.08 c | 4.13 ± 0.68 | 4.27 ± 0.58 a | |
| IC50 | 4.49 ± 0.35 d | 5.34 ± 1.45 a | 7.04 ± 1.14 c,w | |
| IC50x5 | 100.00 ± 0.00 d | 100.00 ± 0.00 d | No cells were detected | |
| SKOV-3 | Non-treated | 2.05 ± 0.56 | 2.07 ± 0.54 | 1.65 ± 0.35 |
| IC50/5 | 2.10 ± 0.19 | 2.33 ± 0.64 | 2.42 ± 0.16 a | |
| IC50 | 3.38 ± 0.69 a | 4.86 ± 0.09 d,w | 11.20 ± 0.39 d,z | |
| IC50x5 | 6.30 ± 0.22 d | 100.00 ± 0.00 d,z | 100.00 ± 0.00 d,z | |
| No | Protein Name a | Gene Name | Accession No b | pIc | mW (Da) c | Sequence Coverage (%) | Mascot Score | Protein Abundance |
|---|---|---|---|---|---|---|---|---|
| 1 | ALDH1A1 | Aldehyde dehydrogenase 1 family member A1 | P00352 | 6.3 | 54,827 | 30% | 233 | −1.52 |
| 2 | PKM | Pyruvate kinase isozymes M1/M2 | P14618 | 7.96 | 57,900 | 54% | 413 | −2.11 |
| 3 | EEF1A1 | Eukaryotic translation elongation factor 1 alpha 1 | P68104 | 9.10 | 50,109 | 33% | 153 | −4.29 |
| 4 | LGALS3 | Galectin-3 | P17931 | 8.57 | 26,136 | 27% | 235 | −6.23 |
| 5 | CAPG | Capping actin protein | P40121 | 5.82 | 38,474 | 29% | 167 | −2.35 |
| 6 | ANXA2 | Annexin A2 | P07355 | 7.57 | 38,580 | 43% | 431 | −5.72 |
| 7 | NVL | Nuclear valosin-containing protein-like | O15381 | 6.11 | 94,991 | 28% | 64 | +8.85 |
| 8 | PNP | Purine nucleoside phosphorylase | P00491 | 6.45 | 32,097 | 42% | 93 | −3.78 |
| 9 | PGAM1 | Phosphoglycerate mutase 1 | P18669 | 6.67 | 28,786 | 57% | 583 | −2.94 |
| 10 | TPI1 | Triosephosphate isomerase 1 | P60174 | 5.65 | 30,772 | 59% | 556 | −2.87 |
| 11 | PRDX3 | Thioredoxin-dependent peroxide reductase | P30048 | 7.67 | 27,675 | 25% | 128 | −2.80 |
| 12 | HNRNPA1 | Heterogeneous nuclear ribonucleoprotein A1 | P09651 | 9.17 | 38,723 | 45% | 195 | −14.65 |
| 13 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | P04406 | 8.57 | 36,030 | 22% | 239 | −4.29 |
| 14 | MDH2 | Malate dehydrogenase | P40926 | 8.92 | 35,481 | 39% | 278 | +4.00 |
| 15 | ACAA2 | Acetyl-CoA acyltransferase 2 | P42765 | 8.32 | 41,898 | 28% | 128 | +4.8 |
| No | Canonical Pathway | p-Value | Proteins Involved |
|---|---|---|---|
| 1 | Glycolysis I | 9.38 × 10−9 | GAPDH, PGAM1, PKM, TPI1 |
| 2 | Gluconeogenesis I | 2.62 × 10−6 | GAPDH, PGAM1, MDH2, |
| 3 | NADH Repair | 4.69 × 10−3 | GAPDH |
| 4 | Xanthine and Xanthosine Salvage | 5.27 × 10−3 | PNP |
| 5 | Guanine and Guanosine Salvage I | 5.86 × 10−3 | PNP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yunos, N.M.; Amin, N.D.M.; Jauri, M.H.; Ling, S.K.; Hassan, N.H.; Sallehudin, N.J. The In Vitro Anti-Cancer Activities and Mechanisms of Action of 9-Methoxycanthin-6-one from Eurycoma longifolia in Selected Cancer Cell Lines. Molecules 2022, 27, 585. https://doi.org/10.3390/molecules27030585
Yunos NM, Amin NDM, Jauri MH, Ling SK, Hassan NH, Sallehudin NJ. The In Vitro Anti-Cancer Activities and Mechanisms of Action of 9-Methoxycanthin-6-one from Eurycoma longifolia in Selected Cancer Cell Lines. Molecules. 2022; 27(3):585. https://doi.org/10.3390/molecules27030585
Chicago/Turabian StyleYunos, Nurhanan Murni, Nor Datiakma Mat Amin, Muhammad Haffiz Jauri, Sui Kiong Ling, Nor Hasnida Hassan, and Nor Jannah Sallehudin. 2022. "The In Vitro Anti-Cancer Activities and Mechanisms of Action of 9-Methoxycanthin-6-one from Eurycoma longifolia in Selected Cancer Cell Lines" Molecules 27, no. 3: 585. https://doi.org/10.3390/molecules27030585