
Potential Antioxidant Activity of Apigenin in the Obviating Stress-Mediated Depressive Symptoms of Experimental Mice
 , , , ,
, , , ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
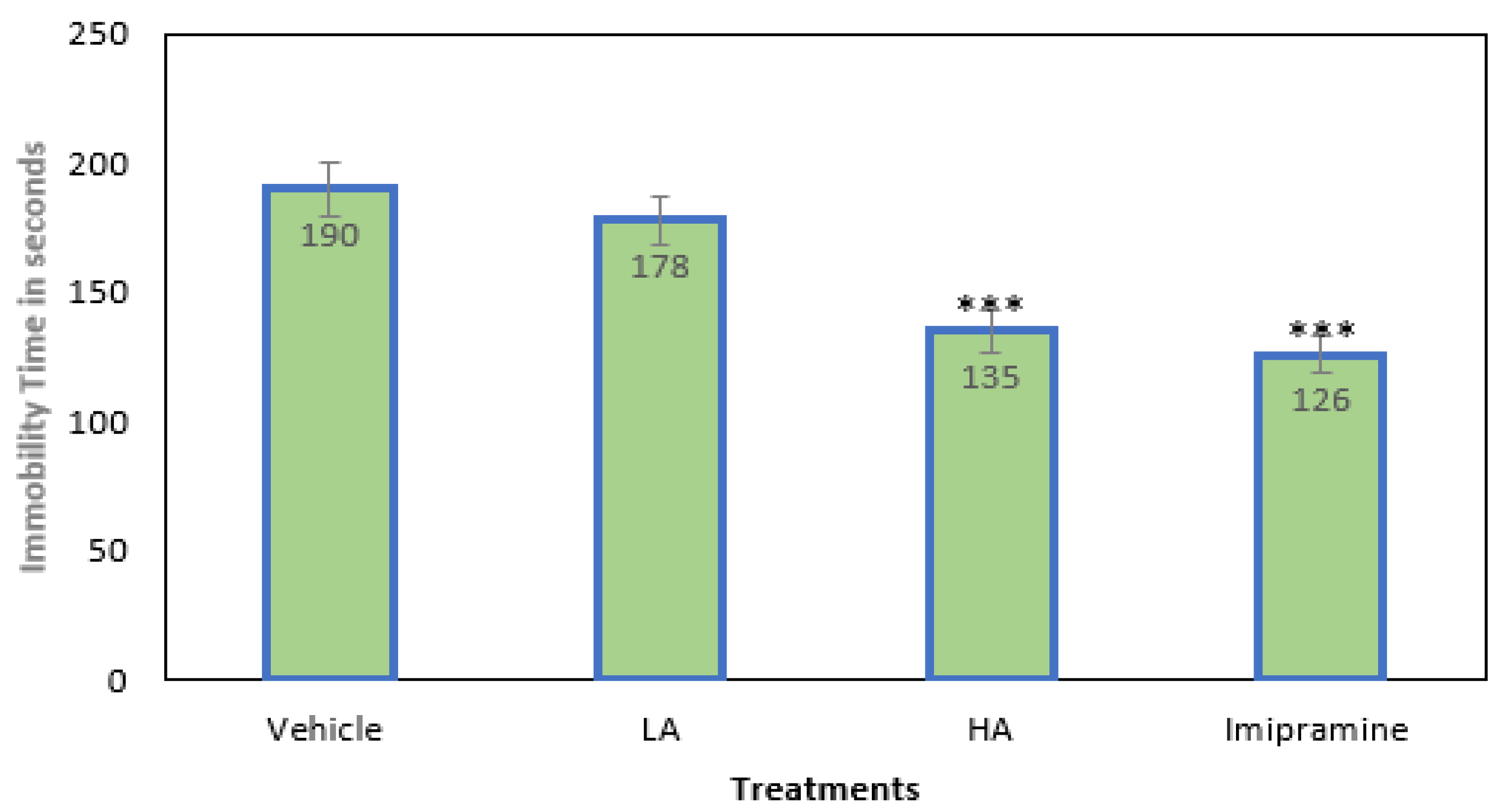
2.1. Immobility Time
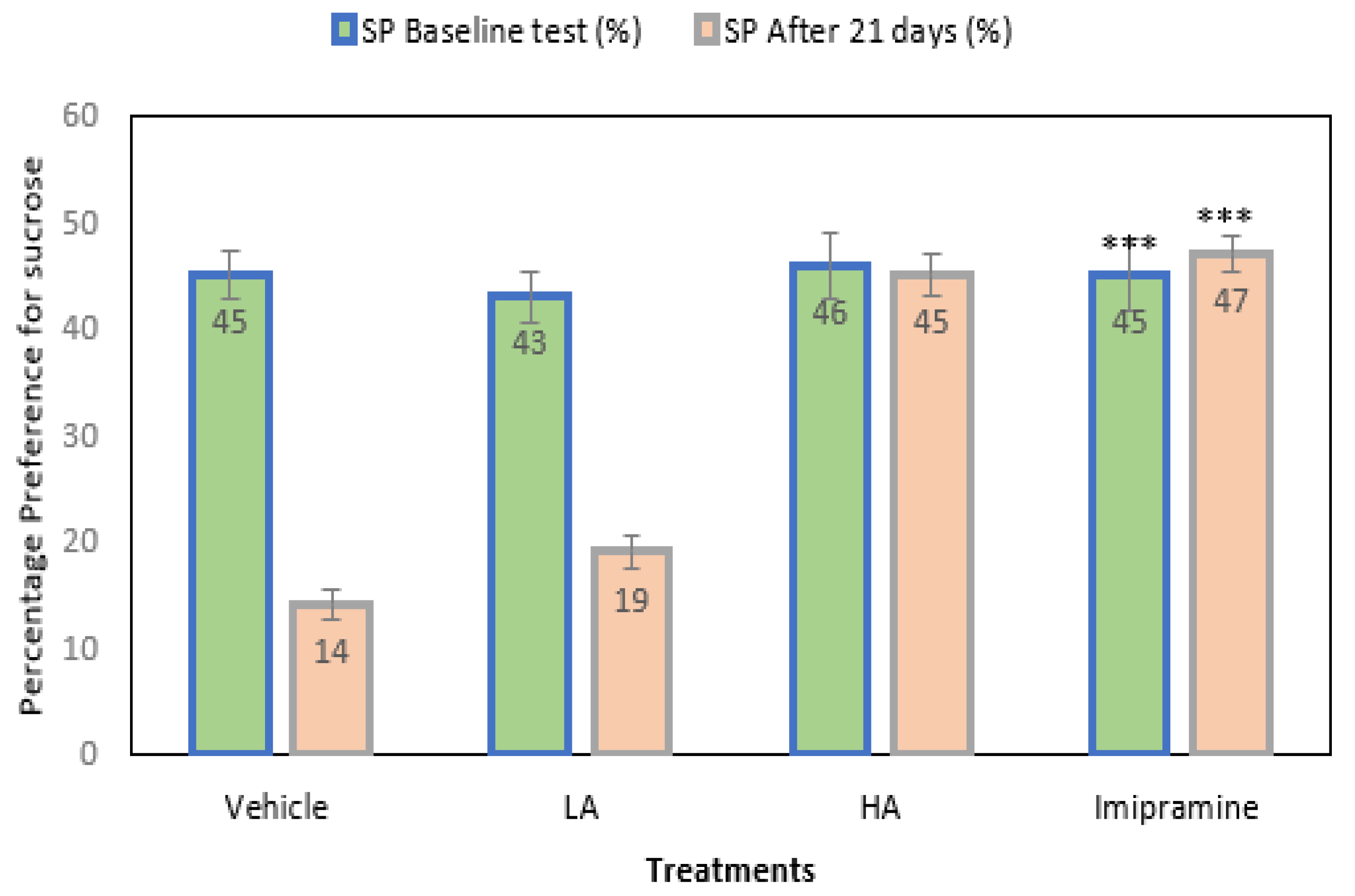
2.2. Sucrose Preference Model
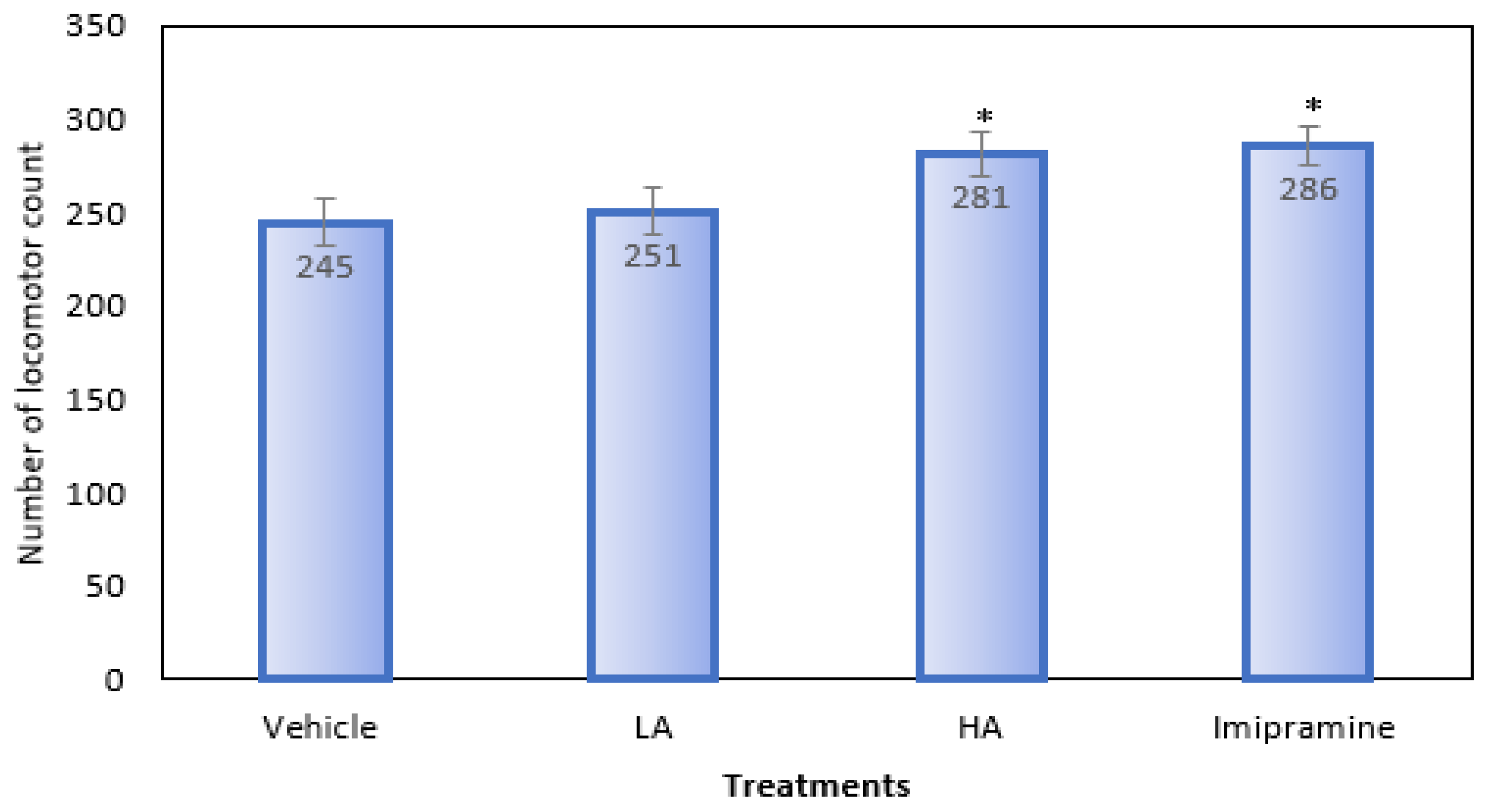
2.3. Locomotor Activity
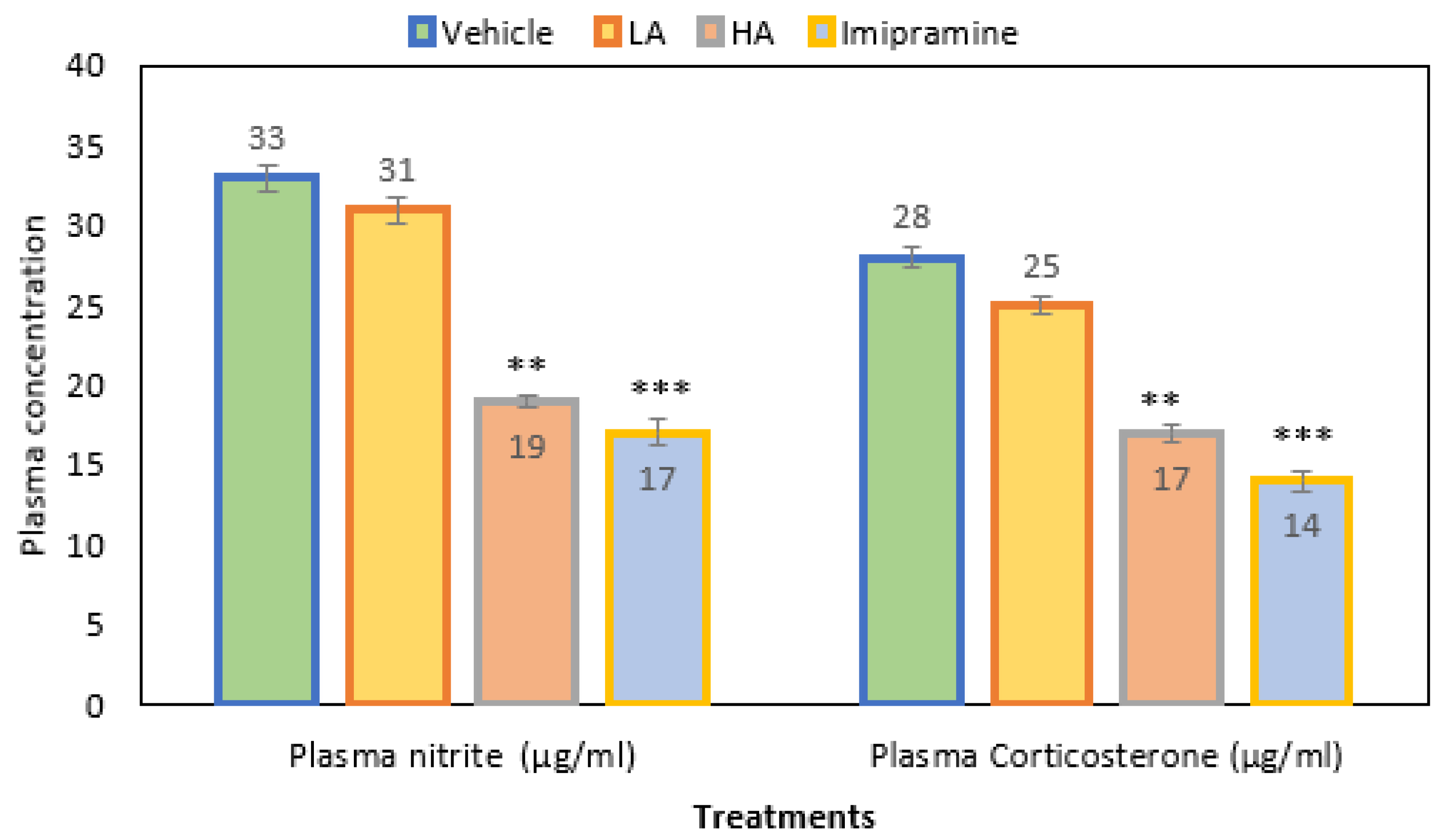
2.4. Plasma Nitrite and Corticosterone
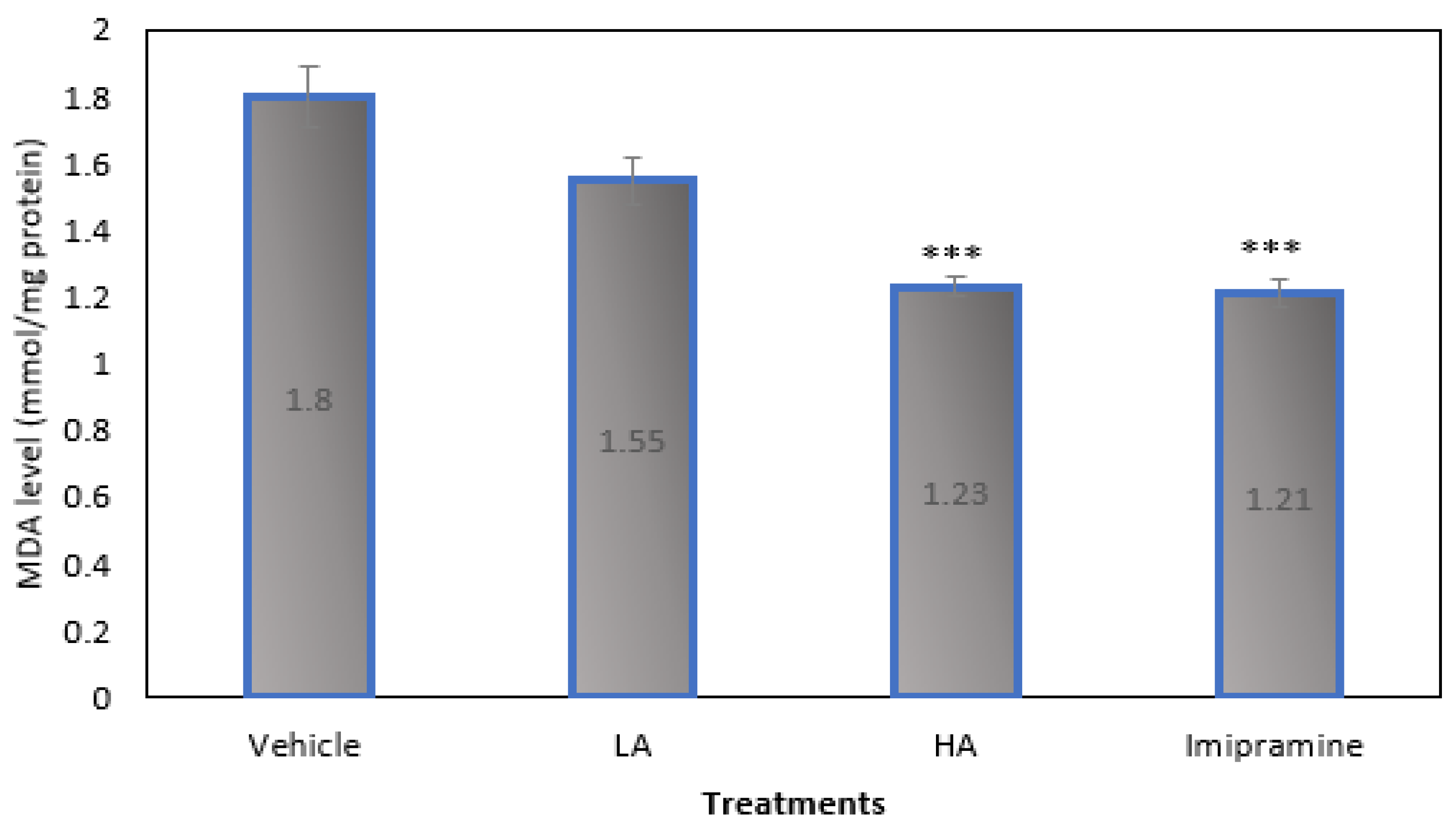
2.5. Brain Malondialdehyde (MDA) Levels
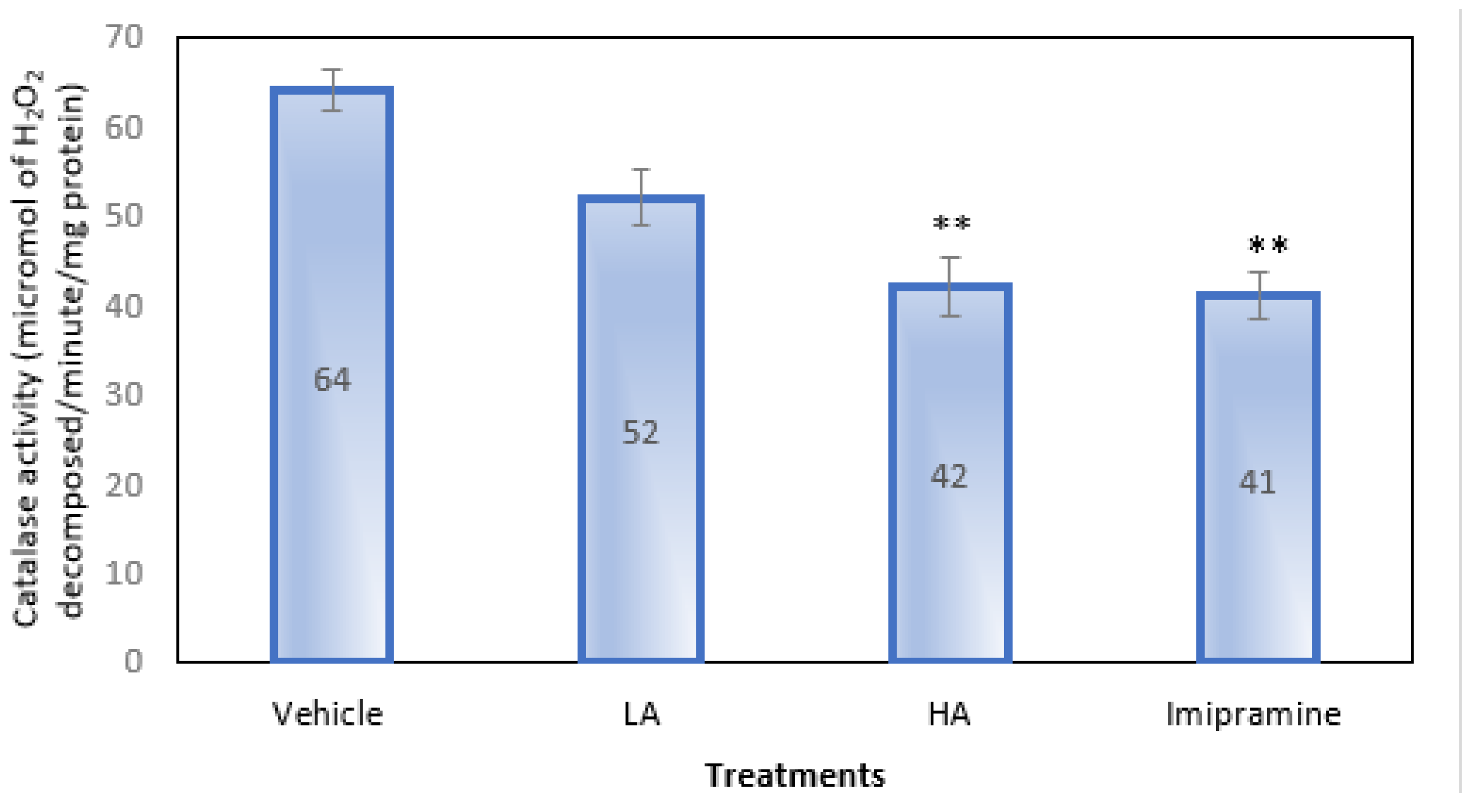
2.6. Brain Catalase Activity
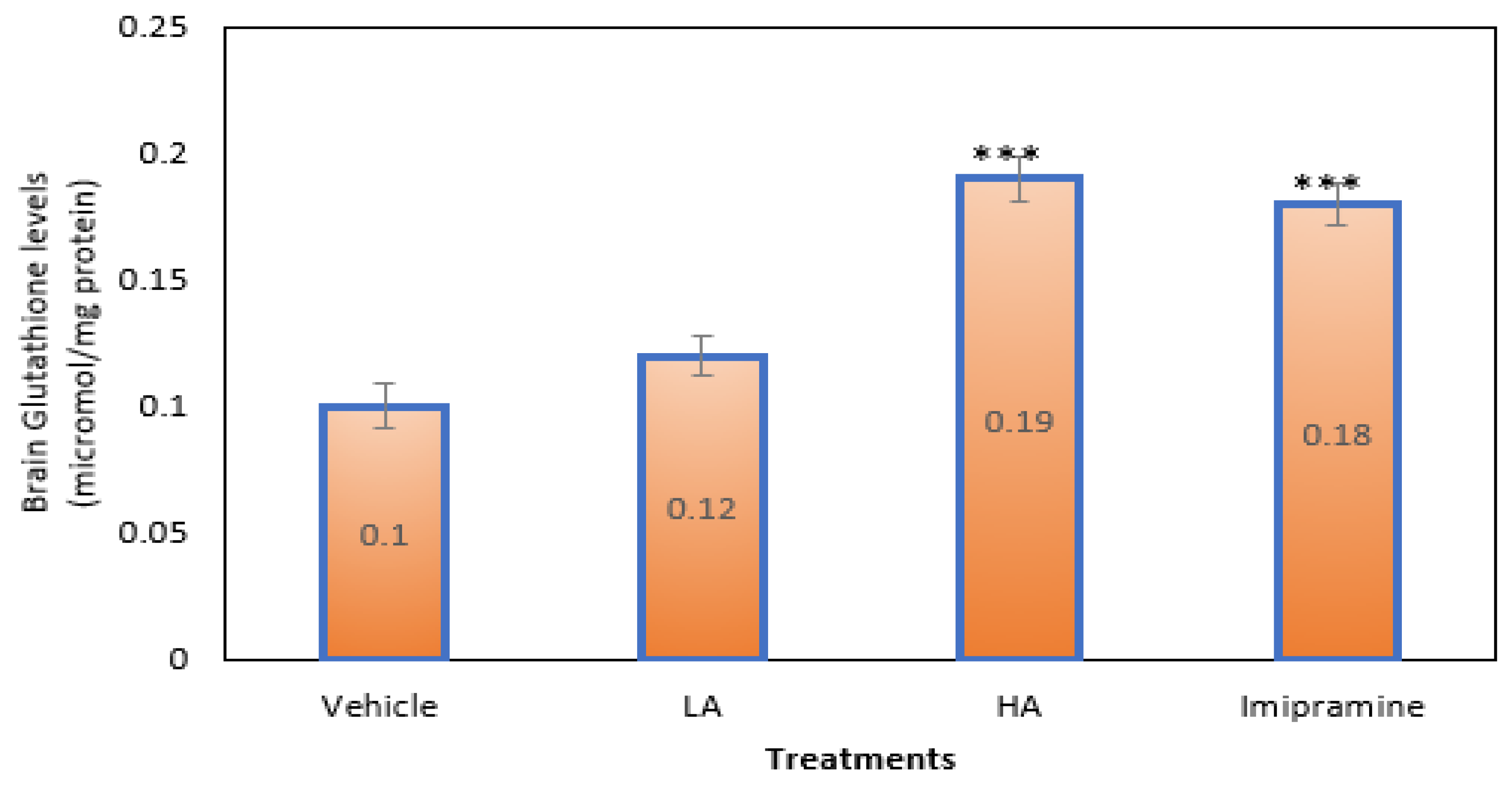
2.7. Brain Glutathione Levels
3. Discussion
4. Materials and Methods
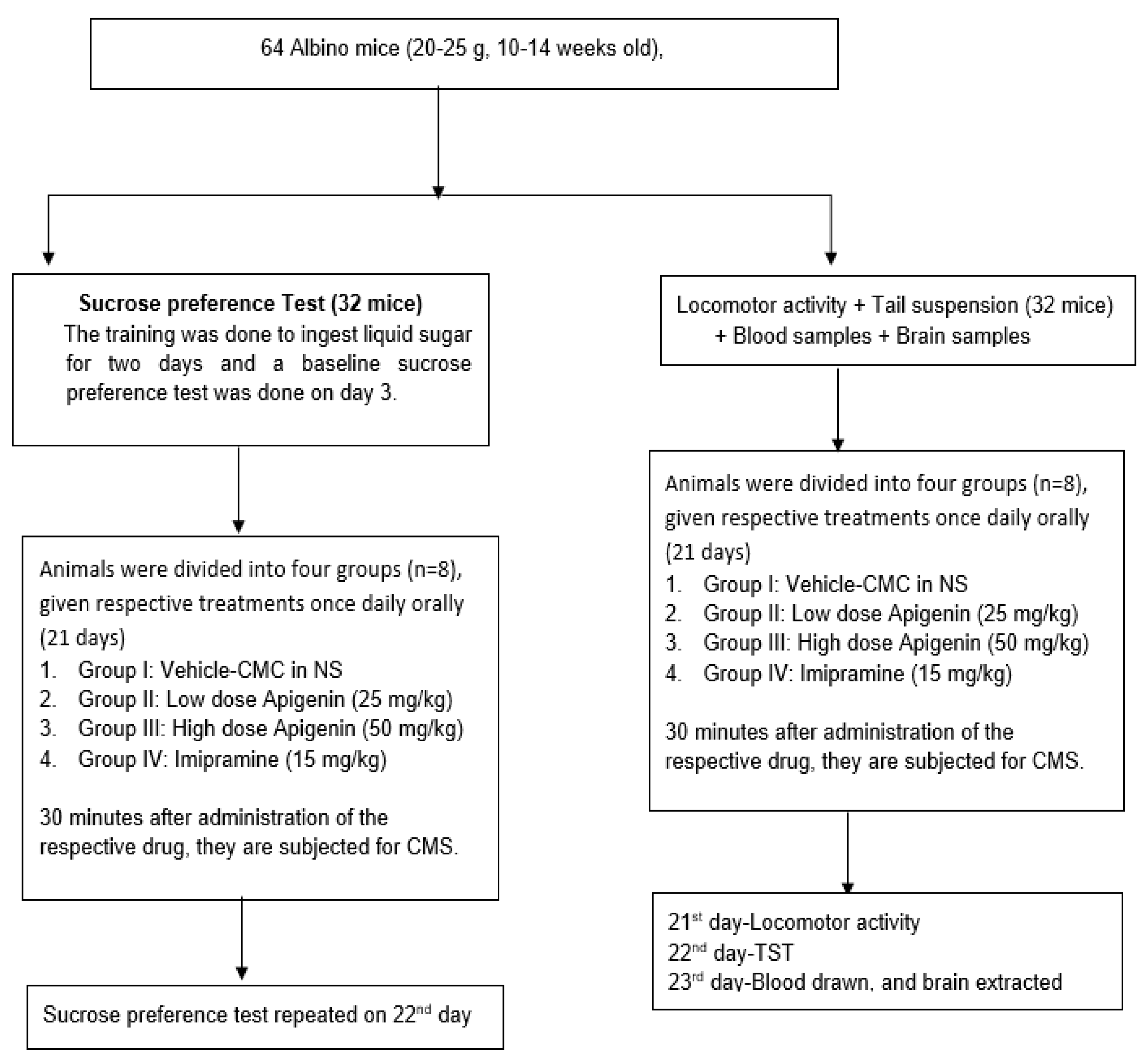
4.1. Study Samples
4.2. Chemicals and Reagents
4.3. Experimental Grouping
4.4. Chronic Mild Stress (CMS)
4.5. Tail Suspension Test
4.6. Measurement of Locomotor Activity
4.7. Sucrose Preference Test
4.8. Nitrite and Corticosterone Measurements
4.9. Reduced Glutathione and Catalase Activity
4.10. Analysis of Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Schechter, L.E.; Ring, R.H.; Beyer, C.E.; Hughes, Z.A.; Khawaja, X.; Malberg, J.E.; Rosenzweig-Lipson, S. Innovative approaches for the development of antidepressant drugs: Current and future strategies. J. Am. Soc. Exp. Neurother. 2005, 2, 590–611. [Google Scholar] [CrossRef]
- Manji, H.K.; Drevets, W.C.; Charney, D.S. The cellular neurobiology of depression. Nat. Med. 2001, 7, 541–547. [Google Scholar] [CrossRef]
- Leonard, B.; Maes, M. Mechanistic explanations how cell-mediated immune activation, inflammation and oxidative and nitrosative stress pathways and their sequels and concimatants play a role in the pathophysiology of unipolar depression. Neurosci. Biobehav. Rev. 2012, 36, 764–785. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, A.; Nomura, S.; Rinsho, N. Pathophysiology of depression. Nihon Rinsho 2007, 65, 1585–1590. [Google Scholar] [PubMed]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O & NS) pathways in major depression and their possible contribution to the (neuro) degenerative process in that illness. Prog. Neuro-Psychophar. Macol. Biol. Psychiatr. 2011, 35, 676–692. [Google Scholar]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories 486 and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Young, E.A.; Haskett, R.F.; Murphy-Weihberg, V.; Watson, S.J.; Akil, H. Loss of gluco-corticoid fast feed back in depression. Arch. Gen. Psychiatr. 1991, 48, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Wróbel, A.; Serefko, A.; Wla´z, P.; Poleszak, E. The effect of imipramine, ketamine, and zinc in the mouse model of depression. Metab. Brain Dis. 2015, 30, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress: And its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef]
- Madrigal, J.L.; Olivenza, R.; Moro, M.A.; Lizasoain, I.; Lorenzo, P.; Rodrigo, J. Glutathione depletion, lipid peroxidation and mitochondrial dysfunction are induced by chronic stress in rat brain. Neuropsychopharmacology 2001, 24, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, H.; Tatsumi, A.; Yamamoto, K.; Kobayashi, F.; Shimoi, K.; Kinae, N. Possible connections among job stress, depressive symptoms, lipid modulation and antioxidants. J. Affect. Disord. 2006, 91, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food sources, bioavailability, metabolism, and bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The therapeutic potential of apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [Green Version]
- Avallone, R.; Zanoli, P.; Puia, G.; Kleinschnitz, M.; Schreier, P.; Baraldi, M. Pharmacological profile of apigenin, a flavonoid isolated from Matricaria chamomilla. Biochem. Pharmacol. 2000, 59, 1387–1394. [Google Scholar] [CrossRef]
- Zanoli, P.; Avallone, R.; Baraldi, M. Behavioral characterisation of the flavonoids apigenin and chrysin. Fitoterapia 2000, 71, S117–S123. [Google Scholar] [CrossRef]
- Asdaq, S.M.B.; Mannasaheb, B.A.; Alanazi, A.; Almusharraf, B.; Alanazi, N.; Saad, K.; Alanazi, S.; Abduallah, K.; Alrashid, S.; Alanazi, F.; et al. Evaluation of skeletal muscle relaxant activity of apigenin in animal experimental models. Int. J. Pharmacol. 2021, 17, 400–407. [Google Scholar]
- Nakazawa, T.; Yasuda, T.; Ueda, J.; Ohsawa, K. Antidepressant-like effects of apigenin and 2, 4, 5-trimethoxycinnamic acid from Perilla frutescens in the forced swimming test. Biol. Pharm. Bull. 2003, 26, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Al-Yamani, M.J.; Asdaq, S.M.B.; Alamri, A.S.; Alsanie, W.F.; Alhomrani, M.; Alsalman, A.J.; Al Hawaj, M.A.; Alanazi, A.A.; Alanzi, K.D.; Imran, M. The role of serotonergic and catecholaminergic systems for possible antidepressant activity of apigenin. Saudi J. Biol. Sci. 2022, 29, 11–17. [Google Scholar] [CrossRef]
- Han, X.H.; Hong, S.S.; Hwang, J.S.; Lee, M.K.; Hwang, B.Y.; Ro, J.S. Monoamine oxidase inhibitory components from Cayratia japonica. Arch. Pharmacal Res. 2007, 30, 13–17. [Google Scholar] [CrossRef]
- Elhwuegi, A.S. Central monoamines and their role in major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Baloch, Z.; Mao, F. Natural Products as a Source for New Leads in Depression Treatment. Evid. Based Complement. Altern. Med. 2022, 2022, 9791434. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Feng, K.; Sun, X.; Xu, L.; Wu, S.; Rahmand, K.; Jia, D.; Han, T. Natural products for the treatment of stress-induced depression: Pharmacology, mechanism and traditional use. J. Ethnopharmacol. 2022, 285, 114692. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology 1997, 134, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, T.; Zhu, Y.; Zheng, C.J.; Ming, Q.L.; Rahman, K.; Qin, L.P. Antidepressant properties of bioactive fractions from the extract of Crocus sativus L. J. Nat. Med. 2010, 64, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bu, H.; Jiang, Y.; Sun, G.; Jiang, R.; Huang, X.; Duan, H.; Huang, Z.; Wu, Q. The antidepressant effects of apigenin are associated with the promotion of autophagy via the mTOR/AMPK/ULK1 pathway. Mol. Med. Rep. 2019, 20, 2867–2874. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, W.Y.; Xia, X.; Kong, L.D. Effects of icariin on hypothalamic-pituitary-adrenal axis action and cytokine levels in stressed Sprague-Dawley rats. Biol. Pharm. Bull. 2006, 29, 2399–2403. [Google Scholar] [CrossRef] [Green Version]
- Sousa, N.; Cerqueira, J.J.; Almeida, O.F. Corticosteroid receptors and neuroplasticity. Brain Res. Rev. 2008, 57, 561–570. [Google Scholar] [CrossRef]
- Swaab, D.F.; Bao, A.M.; Lucassen, P.J. The stress system in the human brain in depression and neurodegeneration. Ageing Res. Rev. 2005, 4, 141–194. [Google Scholar] [CrossRef]
- Bilici, M.; Efe, H.; Köroğlu, M.A.; Uydu, H.A.; Bekaroğlu, M.; Değer, O. Antioxidative enzyme activities and lipid peroxidation in major depression: Alterations by antidepressant treatments. J. Affect. Disord. 2001, 64, 43–51. [Google Scholar] [CrossRef]
- Weng, L.; Guo, X.; Li, Y.; Yang, X.; Han, Y. Apigenin reverses depression-like behavior induced by chronic corticosterone treatment in mice. Eur. J. Pharmacol. 2016, 774, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Kuhad, A.; Chopra, K. Neuropsychopharmacological effect of sesamol in unpredictable chronic mild stress model of depression: Behavioral and biochemical evidence. Psychopharmacology 2011, 214, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Asdaq, S.M.B.; Inamdar, M.N. Potential of Crocus sativus (saffron) and its Constituent, Crocin, as Hypolipidemic and Antioxidant in Rats. Appl. Biochem. Biotechnol. 2010, 162, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.G.; Salles, L.A.; Stein, A.C.; Betti, A.H.; Sakamoto, S.; Cassel, E.; Vargas, R.F.; von Poser, G.L.; Rates, S.M. Antidepressant-like effect of Valeriana glechomifolia Meyer (Valerianaceae) in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 36, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, D.; Joshi, P.; Gupta, A.; Chhillar, R. Possible involvement of monoaminergic neurotransmission in antidepressant-like activity of Emblica officinalis fruits in mice. CNS Neurosci. Ther. 2012, 18, 419–425. [Google Scholar] [CrossRef]
- Alsanie, W.F.; Alamri, A.S.; Abdulaziz, O.; Salih, M.M.; Alamri, A.; Asdaq, S.M.B.; Alhomrani, M.H.; Alhomrani, M. Antidepressant Effect of Crocin in Mice with Chronic Mild Stress. Molecules 2022, 27, 5462. [Google Scholar] [CrossRef]
- Karimi, G.R.; Hosseinzadeh, H.; Khaleghpanah, P. Study of antidepressant effect of aqueous and ethanolic extract of Crocus sativus in mice. Iran. J. Basic Med. Sci. 2001, 4, 11–15. [Google Scholar]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Dhingra, D.; Bansal, S. Antidepressant-like activity of plumbagin in unstressed and stressed mice. Pharmacol. Rep. 2015, 67, 1024–1032. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Bartos, J.; Pesez, M. Colorimetric and fluorimetric determination of steroids. Int. Union Pure Appl. Chem. 1979, 51, 2157–2169. [Google Scholar]
- Wills, E.D. Mechanisms of lipid peroxide formation in tissues: Role of metals and haematin proteins in the catalysis of the oxidation of unsaturated fatty acids. Biochem. Biophys. Acta 1965, 98, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Jollow, D.J.; Mitchell, J.R.; Zampaglione, N.; Gillette, J.R. Bromobenz induced liver necrosis: Protective role of glutathione and evidence for 3,4-bromobenzen- 564 oxide as the hepatotoxic metabolite. Pharmacology 1974, 11, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, A. Catalase activity. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alghamdi, A.; Almuqbil, M.; Alrofaidi, M.A.; Burzangi, A.S.; Alshamrani, A.A.; Alzahrani, A.R.; Kamal, M.; Imran, M.; Alshehri, S.; Mannasaheb, B.A.; et al. Potential Antioxidant Activity of Apigenin in the Obviating Stress-Mediated Depressive Symptoms of Experimental Mice. Molecules 2022, 27, 9055. https://doi.org/10.3390/molecules27249055
Alghamdi A, Almuqbil M, Alrofaidi MA, Burzangi AS, Alshamrani AA, Alzahrani AR, Kamal M, Imran M, Alshehri S, Mannasaheb BA, et al. Potential Antioxidant Activity of Apigenin in the Obviating Stress-Mediated Depressive Symptoms of Experimental Mice. Molecules. 2022; 27(24):9055. https://doi.org/10.3390/molecules27249055
Chicago/Turabian StyleAlghamdi, Adel, Mansour Almuqbil, Mohammad A. Alrofaidi, Abdulhadi S. Burzangi, Ali A. Alshamrani, Abdullah R. Alzahrani, Mehnaz Kamal, Mohd. Imran, Sultan Alshehri, Basheerahmed Abdulaziz Mannasaheb, and et al. 2022. "Potential Antioxidant Activity of Apigenin in the Obviating Stress-Mediated Depressive Symptoms of Experimental Mice" Molecules 27, no. 24: 9055. https://doi.org/10.3390/molecules27249055