
A Joint Approach of Morphological and UHPLC-HRMS Analyses to Throw Light on the Autochthonous ‘Verdole’ Chestnut for Nutraceutical Innovation of Its Waste
 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Morphological Analysis
2.2. Extraction of Chestnut Fruits
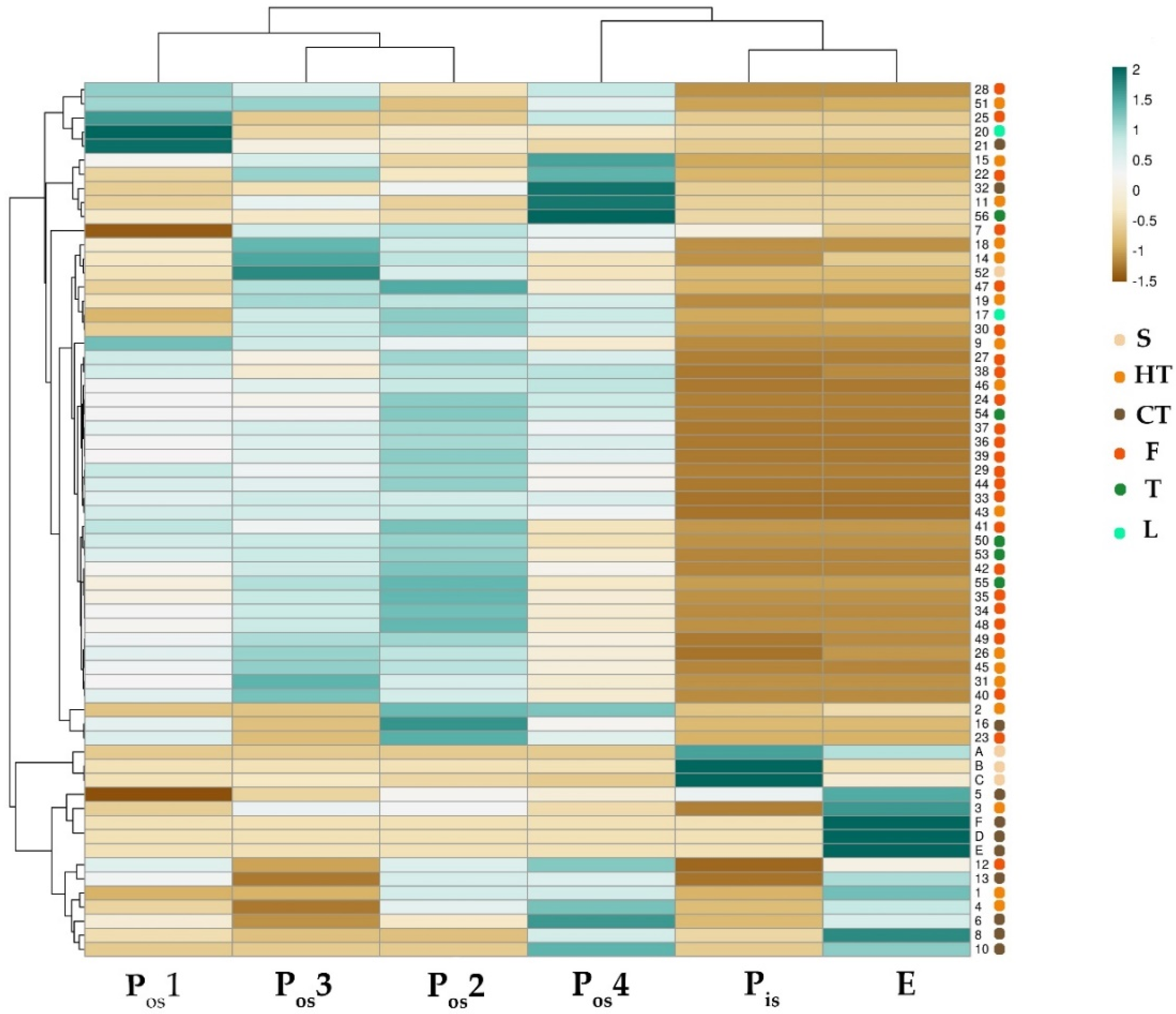
2.3. Shell Components Differ in Phenols and Flavonoids, and Otherwise Act as Antioxidants
2.4. UHPLC-HR-MS/MS Analysis
3. Materials and Methods
3.1. Chestnut Samples
3.2. Extraction of Verdole cv. Chestnut
3.3. Determination of DPPH and ABTS Radical Scavenging Capacity
3.4. Determination of Fe (III) Reducing Power
3.5. Determination of Total Phenols and Total Flavonoids
3.6. Determination of Condensed Tannins
3.7. UHPLC-ESI-QqTOF-MS/MS Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Beccaro, G.; Alma, A.; Bounous, G.; Gomes-Laranjo, J. The Chestnut Handbook Crop & Forest Management, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019; p. 378. [Google Scholar]
- Ferreira-Cardoso, J.V.; Rodrigues, L.; Gomes, E.F.; Sequeira, C.A.; Torres-Perreira, J.M.G. Lipid composition of Castanea sativa Mill. fruits of some native portuguese cultivars. Acta Hortic. 1998, 494, 133–138. [Google Scholar] [CrossRef]
- Gabrielli, A. La Civiltà Del Castagno. Monti Boschi 1994, 65, 3. [Google Scholar]
- Conedera, M.; Giudici, F.; Manetti, M.C. Distribution and economic potential of the sweet chesnut (Castanea sativa Mill.) in Europe. Ecol. Mediterr. 2004, 30, 179–193. [Google Scholar] [CrossRef]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobot. 2004, 13, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Romano, A.; Aponte, M. Chestnut as source of novel ingredients for celiac people. Encycl. Food Secur. Sustain. 2019, 1, 364–368. [Google Scholar]
- Torra, M.; Belorio, M.; Ayuso, M.; Carocho, M.; Ferreira, I.C.F.R.; Barros, L.; Gómez, M. Chickpea and Chestnut Flours as Non-Gluten Alternatives in Cookies. Foods 2021, 10, 911. [Google Scholar] [CrossRef]
- FAO. 2020. Available online: www.fao.org/statistics/en (accessed on 18 November 2022).
- Mgaya, J.; Shombe, G.B.; Masikane, S.C.; Mlowe, S.; Mubofu, E.B.; Revaprasadu, N. Cashew nut shell: A potential bio-resource for the production of bio-sourced chemicals, materials and fuels. Green Chem. 2019, 21, 1186–1201. [Google Scholar] [CrossRef]
- An, J.Y.; Wang, L.T.; Lv, M.J.; Wang, J.D.; Cai, Z.H.; Wang, Y.Q.; Zhang, S.; Yang, Q.; Fu, Y.J. An efficiency strategy for extraction and recovery of ellagic acid from waste chestnut shell and its biological activity evaluation. Microchem. J. 2020, 154, 105616. [Google Scholar] [CrossRef]
- De Vasconcelos, M.C.B.M.; Nunes, F.; Viguera, C.G.; Bennett, R.N.; Rosa, E.A.S.; Ferreira-Cardoso, J.V. Industrial processing effects on chestnut fruits (Castanea sativa Mill.) 3. Minerals, free sugars, carotenoids and antioxidant vitamins. Int. J. Food Sci. 2010, 45, 496–505. [Google Scholar] [CrossRef]
- Braga, N.; Rodrigues, F.; Beatriz, M.; Oliveira, P.P. Castanea sativa by-products: A review on added value and sustainable Application. Nat. Prod. Res. 2015, 29, 1–18. [Google Scholar] [CrossRef]
- Pinto, D.; Vieira, E.F.; Peixoto, A.F.; Freire, C.; Freitas, V.; Costa, P.; Delerue-Matos., C.; Rodriguesa., F. Optimizing the extraction of phenolic antioxidants from chestnut shells by subcritical water extraction using response surface methodology. Food Chem. 2020, 334, 127521. [Google Scholar] [CrossRef]
- Rodriguez, C.P.; Martucci, J.F.; Neira, L.M.; Arbelaiz, A.; Eceiza, A.; Ruseckaite, R.A. Functional properties and in vitro antioxidant and antibacterial effectiveness of pigskin gelatin films incorporated with hydrolysable chestnut tannin. Food Sci. Technol. 2015, 21, 221–231. [Google Scholar] [CrossRef]
- Turrini, F.; Donno, D.; Boggia, R.; Beccaro, G.L.; Zunin, P.; Leardi, R.; Pittaluga, A.M. An innovative green extraction and re-use strategy to valorize food supplement by-products: Castanea sativa bud preparations as case study. Int. Food Res. J. 2019, 115, 276–282. [Google Scholar] [CrossRef]
- Squillaci, G.; Apone, F.; Sena, L.M.; Carola, A.; Tito, A.; Bimonte, M.; de Lucia, A.; Colucci, G.; Cara, F.L.; Morana, A. Chestnut (Castanea sativa Mill.) industrial wastes as a valued bioresource for the production of active ingredients. Process Biochem. 2018, 64, 228–236. [Google Scholar] [CrossRef]
- Cacciola, N.A.; Squillaci, G.; D’Apolito, M.; Petillo, O.; Veraldi, F.; LaCara, F.; Peluso, G.; Margarucci, S.; Morana, A. Castanea sativa Mill. Shells Aqueous Extract Exhibits Anticancer Properties Inducing Cytotoxic and Pro-Apoptotic Effects. Molecules 2019, 24, 3401. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Yang, X.; Chang, X. Bioactive phenolic components and potential health effects of chestnut shell: A review. J. Food Biochem. 2021, 4, 13696. [Google Scholar] [CrossRef]
- Echegaray, N.; Gómez, B.; Barba, F.J.; Franco, D.; Estévez, M.; Carballo, J.; Marszałek, K.; Lorenzo, J.M. Chestnuts and by-products as source of natural antioxidants in meat and meat products: A review. Trends Food Sci. Technol. 2018, 82, 110–121. [Google Scholar] [CrossRef]
- Morana, A.; Squillaci, G.; Paixão, S.M.; Alves, L.; Cara, F.L.; Moura, P. Development of an energy biorefinery model for chestnut (Castanea sativa Mill.) shells. Energies 2017, 10, 1504. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, Z.; Liu, G.; Wu, Y.; Ouyang, J. Inhibitory effect of chestnut (Castanea mollissima Blume) inner skin extract on the activity of α-amylase, α-glucosidase, dipeptidyl peptidase IV and in vitro digestibility of starches. Food Chem. 2020, 324, 126847. [Google Scholar] [CrossRef]
- Ciucure, C.T.; Geana, E.-I.; Sandru, C.; Tita, O.; Botu, M. Phytochemical and Nutritional Profile Composition in Fruits of Different Sweet Chestnut (Castanea sativa Mill.) Cultivars Grown in Romania. Separations 2022, 9, 66. [Google Scholar] [CrossRef]
- Ahmed, S.; Griffin, T.S.; Kraner, D.; Schaffner, M.K.; Sharma, D.; Hazel, M.; Leitch, A.R.; Orians, C.M.; Han, W.; Stepp, J.R.; et al. Environmental Factors Variably Impact Tea Secondary Metabolites in the Context of Climate Change. Front. Plant Sci. 2019, 10, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conidi, C.; Donato, L.; Algieri, C.; Cassano, A. Valorization of chestnut processing by-products: A membrane-assisted green strategy for purifying valuable compounds from shells. J. Clean. Prod. 2022, 378, 134564. [Google Scholar] [CrossRef]
- ISTAT. Coltivazioni: Coltivazioni Legnose Fruttifere. 2022. Available online: http://dati.istat.it/Index.aspx?QueryId=33705 (accessed on 19 November 2022).
- Comegna, L.; Damiano, E.; Greco, R.; Guida, A.; Olivares, L.; Picarelli, L. Effects of the Vegetation on the Hydrological Behavior of a Loose Pyroclastic Deposit. Procedia Environ. Sci. 2013, 19, 922–931. [Google Scholar] [CrossRef] [Green Version]
- Protected Geographical Indication. Available online: http://data.europa.eu/eli/reg_impl/2018/1234/oj (accessed on 20 November 2022).
- Typical Products of Campania. Available online: http://www.agricoltura.regione.campania.it/Tipici/castagna-montella.html (accessed on 20 November 2022).
- Romani, A.; Campo, M.; Lagioia, G.; Scarnicci, M.C.; Paiano, A. A sustainable approach to the re-use of biomass: Synergy between circular agroindustry and Biorefinery Models. In Industrial Symbiosis for the Circular Economy, 1st ed.; Salomone, R., Cecchin, A., Deutz, P., Raggi, A., Cutaia, L., Eds.; Springer: Cham, Switzerland, 2020; pp. 165–179. [Google Scholar]
- UPOV (International Union for the Protection of New Varieties of Plants). Available online: https://www.upov.int/meetings/en/doc_details.jsp?meeting_id=42485&doc_id=368806 (accessed on 7 July 2022).
- Pérez, P.F.; López, J.F. Morphological and phenological description of 38 sweet chestnut cultivars (Castanea sativa Miller) in a contemporary collection. Span. J. Agric. Res. 2009, 7, 829. [Google Scholar] [CrossRef] [Green Version]
- Marinoni, D.T.; Akkak, A.; Beltramo, C.; Guaraldo, P.; Boccacci, P.; Bounous, G.; Maria, A.; Ferrara, A.; Ebone, E.; Viotto, E.; et al. Genetic and morphological characterization of chestnut (Castanea sativa Mill.) germplasm in Piedmont (north-western Italy). Tree Genet. Genomes 2013, 9, 1017–1030. [Google Scholar] [CrossRef]
- Prasad, K.; Hassan, F.; Yang, B.; Kong, K.; Ramanan, R.; Azlan, A.; Ismail, A. Response surface optimisation for the extraction of phenolic compounds and antioxidant capacities of underutilised mangifera pajang kosterm. peels. Food Chem. 2011, 128, 1121–1127. [Google Scholar] [CrossRef]
- Murakami, A.N.N.; de Amboni, R.D.M.C.; Prudêncio, E.S.; Amante, E.R.; de Zanotta, L.M.; Maraschin, M.; Petrus, J.C.C.; Teófilo, R.F. Concentration of phenolic compounds in aqueous mate (Ilex paraguariensis A. St. Hil) extract through nanofiltration. LWT Food Sci. Technol. 2011, 44, 2211–2216. [Google Scholar] [CrossRef]
- Aires, A.; Carvalho, R.; Saavedra, M.J. Valorization of solid wastes from chestnut industry processing: Extraction and optimization of polyphenols, tannins and ellagitannins and its potential for adhesives, cosmetic and pharmaceutical industry. J. Waste Manag. 2016, 48, 457–464. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, M.; Bhandari, B.; Yang, Z. Micronization and nanosizing of particles for an enhanced quality of food: A Review. Crit. Rev. Food Sci. Nutr. 2017, 58, 993–1001. [Google Scholar] [CrossRef]
- Pinto, G.; de Pascale, S.; Aponte, M.; Scaloni, A.; Addeo, F.; Caira, S. Polyphenol profiling of chestnut pericarp, integument and curing water extracts to qualify these food by-products as a source of antioxidants. Molecules 2021, 26, 2335. [Google Scholar] [CrossRef]
- Gullón, B.; Eibes, G.; Dávila, I.; Moreira, M.T.; Labidi, J.; Gullón, P. Hydrothermal treatment of chestnut shells (Castanea sativa) to produce oligosaccharides and antioxidant compounds. Carbohydr. Polym. 2018, 192, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Cerulli, A.; Napolitano, A.; Masullo, M.; Hošek, J.; Pizza, C.; Piacente, S. Chestnut Shells (Italian cultivar “Marrone di roccadaspide” pgi): Antioxidant activity and chemical investigation with in depth LC-hrms/MSN rationalization of tannins. Food Res. Int. 2020, 129, 108787. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, G.; Fontenla, E.; Santos, J.; Freire, M.; González-Álvarez, J.; Antorrena, G. Antioxidant activity and phenolic content of chestnut (Castanea sativa) shell and eucalyptus (Eucalyptus globulus) bark extracts. Ind. Crops Prod. 2008, 28, 279–285. [Google Scholar] [CrossRef]
- Lameirão, F.; Pinto, D.F.; Vieira, E.F.; Peixoto, A.; Freire, C.; Sut, S.; Dall’Acqua, S.; Costa, P.; Delerue-Matos, C.; Rodrigues, F. Green-sustainable recovery of phenolic and antioxidant compounds from industrial chestnut shells using ultrasound-assisted extraction: Optimization and evaluation of biological activities in vitro. Antioxidants 2020, 9, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, D.; Silva, A.M.; Freitas, V.; Vallverdú-Queralt, A.; Delerue-Matos, C.; Rodrigues, F. Microwave-assisted extraction as a green technology approach to recover polyphenols from Castanea sativa shells. ACS Food Sci. Technol. 2021, 1, 229–241. [Google Scholar] [CrossRef]
- Cacciola, N.A.; Cerrato, A.; Capriotti, A.L.; Cavaliere, C.; D’Apolito, M.; Montone, C.M.; Piovesana, S.; Squillaci, G.; Peluso, G.; Laganà, A. Untargeted characterization of chestnut (Castanea sativa Mill.) Shell polyphenol extract: A valued bioresource for prostate cancer cell growth inhibition. Molecules 2020, 25, 2730. [Google Scholar] [CrossRef]
- Nazzaro, M.; Barbarisi, C.; Cara, F.L.; Volpe, M. Chemical and biochemical characterisation of an IGP ecotype chestnut subjected to different treatments. Food Chem. 2011, 128, 930–936. [Google Scholar] [CrossRef]
- González López, N.; Moure, A.; Domínguez, H.; Parajó, J.C. Valorization of chestnut husks by non-isothermal hydrolysis. Ind. Crops Prod. 2012, 36, 172–176. [Google Scholar] [CrossRef]
- Vella, F.M.; Laratta, B.; Cara, F.L.; Morana, A. Recovery of bioactive molecules from chestnut (Castanea sativa Mill.) by-products through extraction by different solvents. Nat. Prod. Res. 2017, 32, 1022–1032. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Abouelenein, D.; Acquaticci, L.; Alessandroni, L.; Abd-Allah, R.H.; Borsetta, G.; Sagratini, C.; Maggi, F.; Vittori, S.; Caprioli, G. Effect of roasting, boiling, and frying processing on 29 polyphenolics and antioxidant activity in seeds and shells of sweet chestnut (Castanea sativa mill.). Plant 2021, 10, 2192. [Google Scholar] [CrossRef]
- Živković, J.; Mujić, I.; Nikolić, G.; Vidović, S.; Mujić, A. Extraction and analysis of condensed tannins in Castanea sativa Mill. J. Cent. Eur. Agric. 2009, 10, 283–290. [Google Scholar]
- Vella, F.M.; de Masi, L.; Calandrelli, R.; Morana, A.; Laratta, B. Valorization of the agro-forestry wastes from Italian chestnut cultivars for the recovery of bioactive compounds. Eur. Food Res. Technol. 2019, 245, 2679–2686. [Google Scholar] [CrossRef]
- Silva, V.; Falco, V.; Dias, M.I.; Barros, L.; Silva, A.; Capita, R.; Alonso-Calleja, C.; Amaral, J.S.; Igregias, G.; Ferreira, I.C.F.R.; et al. Evaluation of the phenolic profile of Castanea sativa Mill. by-products and their antioxidant and antimicrobial activity against Multiresistant bacteria. Antioxidants 2020, 9, 87. [Google Scholar] [CrossRef]
- Dinis, L.; Oliveira, M.M.; Almeida, J.; Costa, R.; Gomes-Laranjo, J.; Peixoto, F. Antioxidant activities of chestnut nut of Castanea sativa Mill. (cultivar ‘Judia’) as function of origin ecosystem. Food Chem. 2012, 132, 1–8. [Google Scholar] [CrossRef]
- Comandini, P.; Lerma-García, M.J.; Simó-Alfonso, E.F.; Toschi, T.G. Tannin analysis of chestnut bark samples (Castanea sativa mill.) by HPLC-Dad–MS. Food Chem. 2014, 157, 290–295. [Google Scholar] [CrossRef]
- Esposito, T.; Celano, R.; Pane, C.; Piccinelli, A.L.; Sansone, F.; Picerno, P.; Zaccardelli, M.; Acquino, R.P.; Mencherini, T. Chestnut (Castanea sativa Miller.) burs extracts and functional compounds: UHPLC-UV-hrms profiling, antioxidant activity, and inhibitory effects on phytopathogenic fungi. Molecules 2019, 24, 302. [Google Scholar] [CrossRef] [Green Version]
- Pimentel-Moral, S.; Cádiz-Gurrea, M.D.; Rodríguez-Pérez, C.; Segura-Carretero, A. Recent advances in extraction technologies of phytochemicals applied for the Revaluation of Agri-Food By-Products. In Functional and Preservative Properties of Phytochemicals, 1st ed.; Prakash, B., Ed.; Academic Press: San Diego, CA, USA, 2020; pp. 209–239. [Google Scholar]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogr. A 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Ham, J.; Kim, H.; Lim, S. Antioxidant and deodorizing activities of phenolic components in chestnut inner shell extracts. Ind. Crops Prod. 2015, 73, 99–105. [Google Scholar] [CrossRef]
- Hong, K.H. Sustainable functional finishing for cotton fabrics using chestnut shell extract. Cellulose 2021, 28, 11729–11743. [Google Scholar] [CrossRef]
- Liu, H.; Fang, Y.; Li, Y.; Ma, L.; Wang, Q.; Xiao, G.; Zou, C. Characterization of PCS-2A, a polysaccharide derived from chestnut shell, and its protective effects against H2O2-induced liver injury in hybrid grouper. Int. J. Biol. Macromol. 2021, 193, 814–822. [Google Scholar] [CrossRef]
- Jimenez, A.G.; Ellermann, M.; Abbott, W.; Sperandio, V. Diet-derived galacturonic acid regulates virulence and intestinal colonization in enterohaemorrhagic escherichia coli and Citrobacter Rodentium. Nat. Microbiol. 2019, 5, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhong, Z.; Ye, Z.; Zhang, W.; He, G.; Zheng, Y.; Huang, S. D-galacturonic acid ameliorates the intestinal mucosal permeability and inflammation of functional dyspepsia in rats. Ann. Palliat. Med. 2021, 10, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Molnár, E.; Nemestóthy, N.; Bélafi-Bakó, K. Utilisation of bipolar electrodialysis for recovery of galacturonic acid. Desalination 2010, 250, 1128–1131. [Google Scholar] [CrossRef]
- Durazzo, A.; Turfani, V.; Azzini, E.; Maiani, G.; Carcea, M. Phenols, lignans and antioxidant properties of legume and sweet chestnut flours. Food Chem. 2013, 140, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Borges, O.; Costa, H.S.; Bennett, R.; Santos, M.; Silva, A.P. Metabolite composition of chestnut (Castanea sativa Mill.) upon cooking: Proximate analysis, fibre, organic acids and phenolics. Food Chem. 2010, 122, 154–160. [Google Scholar] [CrossRef]
- Viriot, C.; Scalbert, A.; du Penhoat, C.L.H.; Moutounet, M. Ellagitannins in woods of sessile oak and sweet chestnut dimerization and hydrolysis during wood ageing. Phytochemistry 1994, 36, 1253–1260. [Google Scholar] [CrossRef]
- Carrero-Carralero, C.; Rodríguez-Sánchez, S.; Calvillo, I.; Martínez-Castro, I.; Soria, A.; Ramos, L.; Sanz, M. Gas chromatographic-based techniques for the characterization of low molecular weight carbohydrates and phenylalkanoid glycosides of sedum roseum root supplements. J. Chromatogr. A 2018, 1570, 116–125. [Google Scholar] [CrossRef]
- Bai, C.; Sun, Y.; Pan, X.; Yang, J.; Li, X.; Wu, A.; Qin, D.; Cao, S.; Wenjun, Z.; Wu, J. Antitumor effects of trimethylellagic acid isolated from Sanguisorba officinalis l. on colorectal cancer via angiogenesis inhibition and apoptosis induction. Front. Pharmacol. 2020, 10, 1646. [Google Scholar] [CrossRef] [Green Version]
- Cerulli, A.; Masullo, M.; Mari, A.; Balato, A.; Filosa, R.; Lembo, S.; Napolitano, A.; Piacente, S. Phenolics from Castanea sativa leaves and their effects on UVB-induced damage. Nat. Prod. Res. 2017, 32, 1170–1175. [Google Scholar] [CrossRef]
- Lei, L.; Chai, Y.; Lin, H.; Chen, C.; Zhao, M.; Xiong, W.; Fan, X. Dihydroquercetin activates AMPK/Nrf2/HO-1 signaling in macrophages and attenuates inflammation in LPS-induced endotoxemic mice. Front. Pharmacol. 2020, 11, 662. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, X.; Yi, X.; Liu, C.; Kong, D.; Zhang, J.; Gong, M. Myricetin: A potent approach for the treatment of type 2 diabetes as a natural class B GPCR agonist. FASEB J. 2017, 31, 2603–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalai, F.Z.; Boulaaba, M.; Ferdousi, F.; Isoda, H. Effects of isorhamnetin on diabetes and its associated complications: A review of in vitro and in vivo studies and a post hoc transcriptome analysis of involved molecular pathways. Int. J. Mol. Sci. 2022, 23, 704. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Peng, H.; Wu, X.; Xu, X.; Kuang, T.; Zhang, J.; Du, L.; Fan, G. The therapeutic effects and mechanisms of quercetin on metabolic diseases: Pharmacological Data and clinical evidence. Oxid. Med. Cell. Longev. 2021, 2021, 6678662. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.J.; Pecio, Ł.; Kowalczyk, M.; Kontek, R.; Gajek, G.; Stopinsek, L.; Mirt, I.; Stochmal, A.; Oleszek, W. Cytotoxic triterpenoids isolated from sweet chestnut heartwood (Castanea sativa) and their health benefits implication. Food Chem. Toxicol. 2017, 109, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, S.; Galasso, S.; Piccolella, S.; Kretschmer, N.; Pan, S.; Marciano, S.; Bauer, R.; Monaco, P. Seasonal variation in phenolic composition and antioxidant and anti-inflammatory activities of Calamintha nepeta (L.) savi. Food Res. Int. 2015, 69, 121–132. [Google Scholar] [CrossRef]
- Fiorentino, M.; Gravina, C.; Piccolella, S.; Pecoraro, M.T.; Formato, M.; Stinca, A.; Pacifico, S.; Esposito, A. Calendula arvensis (Vaill.) L.: A Systematic Plant Analysis of the Polar Extracts from Its Organs by UHPLC-HRMS. Foods 2022, 11, 247. [Google Scholar] [CrossRef]
- Formato, M.; Piccolella, S.; Zidorn, C.; Pacifico, S. UHPLC-HRMS Analysis of Fagus sylvatica (Fagaceae) Leaves: A Renewable Source of Antioxidant Polyphenols. Antioxidants 2021, 10, 1140. [Google Scholar] [CrossRef]
- Pacifico, S.; D’Abrosca, B.; Scognamiglio, M.; D’Angelo, G.; Gallicchio, M.; Galasso, S.; Monaco, P.; Fiorentino, A. NMR-based metabolic profiling and in vitro antioxidant and hepatotoxic assessment of partially purified fractions from Golden Germander (Teucrium polium L.) methanolic extract. Food Chem. 2012, 135, 1957–1967. [Google Scholar] [CrossRef]
- Formato, M.; Piccolella, S.; Zidorn, C.; Vastolo, A.; Calabrò, S.; Cutrignelli, M.I.; Pacifico, S. UHPLC-ESI-QQTOF analysis and in vitro rumen fermentation for exploiting Fagus sylvatica leaf in ruminant diet. Molecules 2022, 27, 2217. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and Delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VERDOLE DESCRIPTOR ATTRIBUTED | ||
|---|---|---|
 | Tree: vigor Tree: growth habit | Medium Upright |
 | Current season’s shoot: thickness Current season’s shoot: length of internodes Current season’s shoot: arrangement of leaves Current season’s shoot: color of upper side stem Current season’s shoot: density of lenticels Shoot: number of female flowers | Thick Long Opposite Yellow brown Dense Many |
 | Male flower: length of filament Catkin: length | Very long Long |
 | Young leaf: bronze coloration Leaf: size Leaf: profile in cross-section Leaf: symmetry Leaf: length/width ratio Leaf: attitude in relation to shoot Leaf blade: green color intensity of upper side Leaf: color of lower side Leaf: shape Leaf: shape of apex Leaf: the shape of base Leaf: shape of margin Leaf: symmetry of base Leaf: color of petiole Leaf: ratio length of leaf blade/length of petiole | Absent Large Slightly concave Moderately asymmetric High Upwards Medium Light green Narrow elliptic Broad acuminate Cordate Acute Symmetric or slightly asymmetric Green High |
 | Bur: shape Bur: density of prickles | Obloid Sparse |
 | Nut: embryony Nut: degree of seed coat penetration into embryo Nut: shape Nut: area of pubescence on upper part Nut: area of hilum Nut: shape of border line of hilum and pericarp Nut: conspicuousness of hilum Nut: glossiness Nut: color of skin Nut: size | Mono-embryonic Medium Circular Small Small Wavy Conspicuous Present Reddish brown Medium |
 | Seed coat: adherence to kernel Kernel: color of flesh | Weak Yellow |
 | Time of leaf bud burst Time of male flowering Time of female flowering Time of maturity for consumption | Medium Early Medium Early |
| Cultivar | State | Extraction Method | Extraction Solvent | Analytical Tools | TPC | TFC | TcTC | AA | Refs. |
|---|---|---|---|---|---|---|---|---|---|
| – | Italy | Maceration (∅ 1–2 mm) | EtOH: H2O (different ratios v:v) | HPLC-UV-VIS | X | x | [22] | ||
| – | Spain | Acid hydrolysis (∅ 0.4 mm) | – | FTIR; TGA; MALDI-TOF-MS | X | x | x | [38] | |
| Marrone di Roccadaspide | Italy | Maceration (RT) | MeOH, PET, CHCl3 | LC-ESI/QTrap/MSn | X | x | x | x | [39] |
| – | Spain | Soxhlet | Several solvents | FT-IR; UV/Vis | X | x | [40] | ||
| – | Italy | SWE | H2O | LC-DAD/ESI-MS | X | x | [37] | ||
| – | Portugal | UAM | H2O | LC-UV-MS; 1H NMR | X | x | [41] | ||
| – | Portugal | Microwave | EtOH: H2O (different ratios, v:v) | LC-ESI/MS | x | x | x | [42] | |
| – | Italy | Decoction | Boiling H2O (1 h) | UHPLC-MS/MS | x | x | [43] | ||
| Palomina | Italy | Ultra Turrax homogenizer; Decoction | Boiling H2O (30 min) | HPLC-UV | x | x | [44] | ||
| – | Spain | N.I.H | Alkaline solution or H2O | HPLC | x | x | [45] | ||
| – | Italy | Maceration (RT) | MeOH (60%—1:10, w:v); EtOH (60%—1:10, w:v); H2O (1:40, w:v) | HPLC-DAD | x | x | x | [46] | |
| – | – | UAM (59 kHz) | EtOH: H2O (7:3, v:v) | HPLC-MS/MS | x | x | x | [47] | |
| – | Bosnia | UAM (50 Hz and 125 W) | EtOH | – | x | x | x | [48] | |
| Napoletana; Mercogliana Tempestiva; Bouche | Italy | Decoction | Boiling H2O (40 min) | HPLC-UV-DAD | x | x | x | [49] | |
| – | Portugal | Maceration (RT) | EtOH | HPLC-DAD/ESI-MS | x | x | x | [50] | |
| – | Italy | Maceration | Boiling H2O (1 h) | x | x | x | x | [16] | |
| Judia | Maceration | EtOH | HPLC-UV | x | x | x | [51] | ||
| – | Italy | UAM (35 kHz) | MeOH | HPLC DAD/ESI-MS | x | x | [52] | ||
| – | Italy | – | MeOH (after defatting) EtOH:H2O (1:1, v:v) | UHPLC-UV/ESI-HRMS | x | x | [53] |
| Peak | RT (Min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) and Relative Intensity (%) |
|---|---|---|---|---|---|---|---|
| 1 | 0.398 | Hexahydroxydiphenoyl hexose | C20H18O14 | 481.0640 | 3.4 | 12 | 481.0638; 300.9986; 275.0193; 257.0083 |
| 2 | 0.491 | Gallic acid hexoside | C13H16O10 | 331.0663 | −2.3 | 6 | 169.0137; 168.0073; 125.0245 |
| 3 | 0.674 | Gallic acid | C7H6O5 | 169.0146 | 2.1 | 2 | 169.0146; 125.0248 |
| 4 | 0.829 | Galloyl-HHDP-hexose | C27H22O18 | 633.0739 | 0.9 | 17 | 633.0761; 300.9978; 275.0193 |
| 5 | 0.967 | Crenatin | C13H18O9 | 317.0886 | 2.5 | 5 | 317.0891; 155.0362; 137.0242 |
| 7 | 1.187 | Dihydroxybenzoic acid | C7H6O4 | 153.0200 | 4.4 | 5 | 109.0299; 108.0220; |
| 9 | 1.579 | diHHDP-hexose | C34H24O22 | 783.0709 | 2.9 | 23 | 783.0707; 481.0627; 300.9984; 275.0191 |
| 11 | 1.823 | Castalagin/ vescalagin | C34H30O31 | 933.0673 | −2.7 | 20 | 933.0677; 631.0593; 300.9977 |
| 14 | 4.838 | Castacrenin | C27H18O17 | 613.0487 | 2.6 | 10 | 613.0511; 493.0076; 300.9988 |
| 15 | 5.665 | Digalloyl hexose | C20H20O14 | 483.0780 | −0.1 | 11 | 483.0757; 313.0554; 211.0241; 271.0462 169.0150; 125.0250; |
| 18 | 8.019 | Trigalloylhexose 1 | C27H24O18 | 635.0890 | 0 | 16 | 635.0947; 483.0800; 465.0701; 423.0607; 313.0567; 169.0143 |
| 19 | 8.085 | Trigalloylhexose 2 | C27H24O18 | 635.0891 | 0.2 | 16 | 635.0932; 589.1926; 521.2044; 465.0707; 359.1537; 313.0587; 169.0142 |
| 26 | 9.680 | Ellagic acid | C14H6O8 | 300.9986 | −1.3 | 12 | 301.0000; 283.9974; 185.0243 |
| 31 | 10.069 | diHDDP-deoxyhexoside | C34H22O20 | 749.0643 | 1.5 | 24 | 447.0579; 300.9991; 299.9901 |
| 41 | 11.434 | Methyl ellagic acid | C15H8O8 | 315.0145 | −0.4 | 12.0 | 299.9907; 298.9814; 216.0057; 172.0169 |
| 44 | 12.245 | Dimethyl ellagic acid deoxyhexoside | C22H20O12 | 475.0888 | 1.3 | 13.0 | 475.0912; 460.0681; 328.0228; 312.9991; 297.9757 |
| 46 | 12.855 | Dimethylellagic acid | C16H10O8 | 329.0304 | 0.3 | 12.0 | 329.0307; 314.0073; 298.9837; 285.0038; 270.9887; 242.9934; 214.9982 |
| 47 | 14.272 | Trimethylellagic acid eptuloside | C24H24O14 | 535.1109 | 2.9 | 13.0 | 343.0461; 328.0230; 312.9991; 297.9749 |
| 51 | 15.135 | Trimethyl ellagic acid | C17H12O8 | 343.0462 | 0.8 | 12.0 | 312.9988; 297.9755; 285.0036; 269.9804;213.9901 |
| Peak | RT (Min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) and Relative Intensity (%) |
|---|---|---|---|---|---|---|---|
| 6 | 1.147 | Prodelphinidin B-type I | C30H26O14 | 609.1258 | 1.3 | 18 | 609.1278; 441.0834; 423.0723; 305.0664; 177.0193; 483.0942; 261.0756 |
| D | 1.308 | Prodelphinidin B-type II | C30H26O14 | 609.1247 | −2 | 18 | 609.1258; 441.0837; 423.0724; 305.0667; 177.0194 |
| 8 | 1.503 | Gallocatechin | C15H14O7 | 305.0665 | −0.6 | 9 | 305.0650; 261.0726; 237.0735; 219.0661; 179.0337; 167.0342; 137.0240; 125.0243; 111.0446 |
| 10 | 1.805 | Procyanidin | C30H26O13 | 593.1289 | −2.0 | 18 | 593.1312; 467.1045; 425.0900; 407.0795; 303.0546; 289.0719; 177.0202 |
| E | 2.755 | (epi)Catechin derivative I | C30H26O14 | 609.1244 | −1 | 18 | 609.1249; 565.1344; 457.0764; 407.0758; 319.0452; 289.0706; 275.0553; 231.0653; 165.0189 |
| 12 | 4.387 | Catechin | C15H14O6 | 289.0719 | 0.5 | 9 | 289.0735; 271.0650; 245.0847; 221.0825; 205.0523; 203.0722; 187.0414; 175.0404; 151.0418; 137.0251; 123.0457; 109.0312 |
| 13 | 4.564 | Procyanidin B-type | C30H26O12 | 577.1349 | −0.4 | 18 | 577.1399; 451.1277; 425.0900; 407.0788; 289.0740; 245.0836; 125.0250 |
| F | 4.778 | (epi)Catechin derivative II | C30H26O14 | 609.1245 | −0.8 | 18 | 609.1257; 485.1241; 407.0771; 289.0696; 231.0656; 206.0667; 102.0995 |
| 16 | 7.229 | Procyanidin A-type | C30H24O13 | 591.1158 | 2.3 | 19 | 591.1226; 439.0687; 421.0576; 285.0419; 177.0183 |
| 21 | 8.145 | Procyanidin B-type O-gallate | C37H30O16 | 729.1475 | 1.9 | 23 | 729.1506; 577.1385; 451.1053; 425.0894; 407.0778; 289.0723; 245.0450; 125.0267 |
| 22 | 8.379 | (epi)Catechin gallate | C22H18O10 | 441.0845 | 4.0 | 14 | 441.0796; 289.0731 |
| 32 | 10.073 | Procyanidin B-type gallate | C37H30O16 | 729.1452 | −1.2 | 23 | 729.1507; 577.1343; 407.0781; 289.0715; 269.0462; 125.0245 |
| Peak | RT (Min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) and Relative Intensity (%) |
|---|---|---|---|---|---|---|---|
| 23 | 8.842 | Dihydrokaempferol deoxyhexoside | C21H22O11 | 449.1097 | 1.7 | 11 | 449.1094; 287.0557; 269.0444; 151.0035 |
| 24 | 9.395 | Myricetin hexoside | C21H20O13 | 479.0827 | −0.9 | 12 | 479.0871; 317.0336; 316.0234; 287.0219 |
| 25 | 9.494 | Dihydroquercetin galloyl hexoside (e.g., taxillusin) | C28H26O16 | 617.1152 | 0.6 | 5 | 617.1215; 491.0869; 465.1078; 313.0584; 303.0520; 194.9932 |
| 27 | 9.682 | Myricetin deoxyhexoside | C21H20O12 | 463.0882 | 0 | 12 | 463.0910; 317.0315; 316.0238; 287.0207; 316.0238; 271.0254 |
| 28 | 9.688 | Quercetin hexoside gallate | C21H28O21 | 615.1050 | −0.7 | 8.0 | 615.1046; 463.0907; 301.0373; 300.0281 |
| 29 | 10.014 | Phloretin hexoside | C21H24O10 | 435.1296 | −0.2 | 10.0 | 345.0967; 315.0860; 273.0772; 209.0455; 167.0348; 123.0450 |
| 30 | 10.033 | Quercetin hexoside | C21H20O12 | 463.0892 | 2.2 | 12.0 | 463.0892; 301.0251; 300.0280; 271.0242 |
| 33 | 10.290 | Patuletin hexoside | C22H22O13 | 493.0986 | −0.3 | 12 | 493.1081; 331.0468; 315.0141; 287.0188; 271.0237; 151.0033 |
| 34 | 10.539 | Dihydroquercetin | C15H12O7 | 303.0514 | 1.2 | 10.0 | 303.0498; 151.0409; 123.0448 |
| 35 | 10.870 | Quercetin deoxyhexoside | C21H20O11 | 447.0936 | 0.7 | 12.0 | 447.0964; 300.0291; 271.0261; 151.0028 |
| 36 | 11.055 | Isorhamnetin rutinoside | C28H32O16 | 623.1619 | 0.2 | 13.0 | 623.1675; 315.0527; 314.0440; 300.0264 |
| 37 | 11.115 | Isorhamnetin deoxyhexoside | C22H22O12 | 477.1036 | −0.5 | 12.0 | 331.0480; 314.0456; 299.0212; 271.0267 |
| 38 | 11.185 | Naringenin gallate | C22H16O9 | 423.0723 | 0.3 | 15 | 423.0747; 299.0204; 271.0253; 258.0168; 243.0303 |
| 39 | 11.331 | Trihydroxy dimethoxyflavanol hexoside | C23H24O13 | 507.1146 | 0.4 | 12 | 507.1195; 345.0639; 344.0559; 273.0418 |
| 40 | 11.653 | Isorhamnetin pentoside | C21H20O11 | 447.0937 | 0.9 | 12 | 447.0936; 315.0527; 314.0440; 300.0285; 285.0416; 271.0245; 243.0300 |
| 42 | 12.035 | Isorhamnetin hexoside | C22H22O11 | 461.1093 | 0.8 | 12.0 | 461.1110; 315.0500; 314.0433; 271.0240; 243.0284 |
| 43 | 12.315 | Dihydroxy trimethoxyflavone deoxyhexoside | C23H24O12 | 491.1205 | 2.0 | 12.0 | 491.1244; 345.0637; 344.0558; 329.0323; 315.0164; 301.0367; 273.0412 |
| 45 | 14.507 | Isokaempferide | C16H12O6 | 299.0559 | −0.7 | 11.0 | 284.0322; 256.0370; 255.0286; 227.0343; 151.0038 |
| 48 | 14.510 | Methoxyquercetin | C16H12O7 | 315.0512 | 0.6 | 11 | 315.0530; 300.0257; 283.0235; 271.0240; 255.0287; 151.0031 |
| 49 | 14.609 | Dimethoxyquercetin | C16H14O8 | 345.0615 | −0.3 | 11 | 345.0607; 330.0362; 315.0144; 287.0186; 259.0240; 203.0388; 171.0452 |
| Peak | RT (Min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) and Relative Intensity (%) |
|---|---|---|---|---|---|---|---|
| 50 | 14.665 | Trihydroxyoctadecadienoic acid | C18H32O5 | 327.2180 | 0.9 | 3 | 327.2196;229.1437; 211.1335; 171.1026 |
| 52 | 17.309 | Chestnoside | C36H54O11 | 661.3606 | 1.9 | 10.0 | 661.3650; 499.3099; 419.2973 |
| 53 | 17.557 | Bartogenic acid | C30H46O7 | 517.3182 | 2.2 | 8.0 | 517.3209; 455.3186; 437.3076 |
| 54 | 19.959 | Dihydroxyursadiendioic acid (I) | C30H44O6 | 499.3075 | 2.0 | 9.0 | 499.3075; 455.3160; 437.3050; 419.2946 |
| 55 | 19.967 | Hederagenin | C29H44O5 | 471.3116 | 2.1 | 8.0 | 471.3119; 453.3013; 409.3107; 379.3000; 363.2712 |
| 56 | 20.369 | Dihydroxyursadiendioic acid (II) | C30H44O6 | 499.3065 | 0.1 | 9.0 | 499.3085 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrara, E.; Pecoraro, M.T.; Cice, D.; Piccolella, S.; Formato, M.; Esposito, A.; Petriccione, M.; Pacifico, S. A Joint Approach of Morphological and UHPLC-HRMS Analyses to Throw Light on the Autochthonous ‘Verdole’ Chestnut for Nutraceutical Innovation of Its Waste. Molecules 2022, 27, 8924. https://doi.org/10.3390/molecules27248924
Ferrara E, Pecoraro MT, Cice D, Piccolella S, Formato M, Esposito A, Petriccione M, Pacifico S. A Joint Approach of Morphological and UHPLC-HRMS Analyses to Throw Light on the Autochthonous ‘Verdole’ Chestnut for Nutraceutical Innovation of Its Waste. Molecules. 2022; 27(24):8924. https://doi.org/10.3390/molecules27248924
Chicago/Turabian StyleFerrara, Elvira, Maria Tommasina Pecoraro, Danilo Cice, Simona Piccolella, Marialuisa Formato, Assunta Esposito, Milena Petriccione, and Severina Pacifico. 2022. "A Joint Approach of Morphological and UHPLC-HRMS Analyses to Throw Light on the Autochthonous ‘Verdole’ Chestnut for Nutraceutical Innovation of Its Waste" Molecules 27, no. 24: 8924. https://doi.org/10.3390/molecules27248924