

Allicin Alleviates Diabetes Mellitus by Inhibiting the Formation of Advanced Glycation End Products

Abstract
:1. Introduction
2. Results
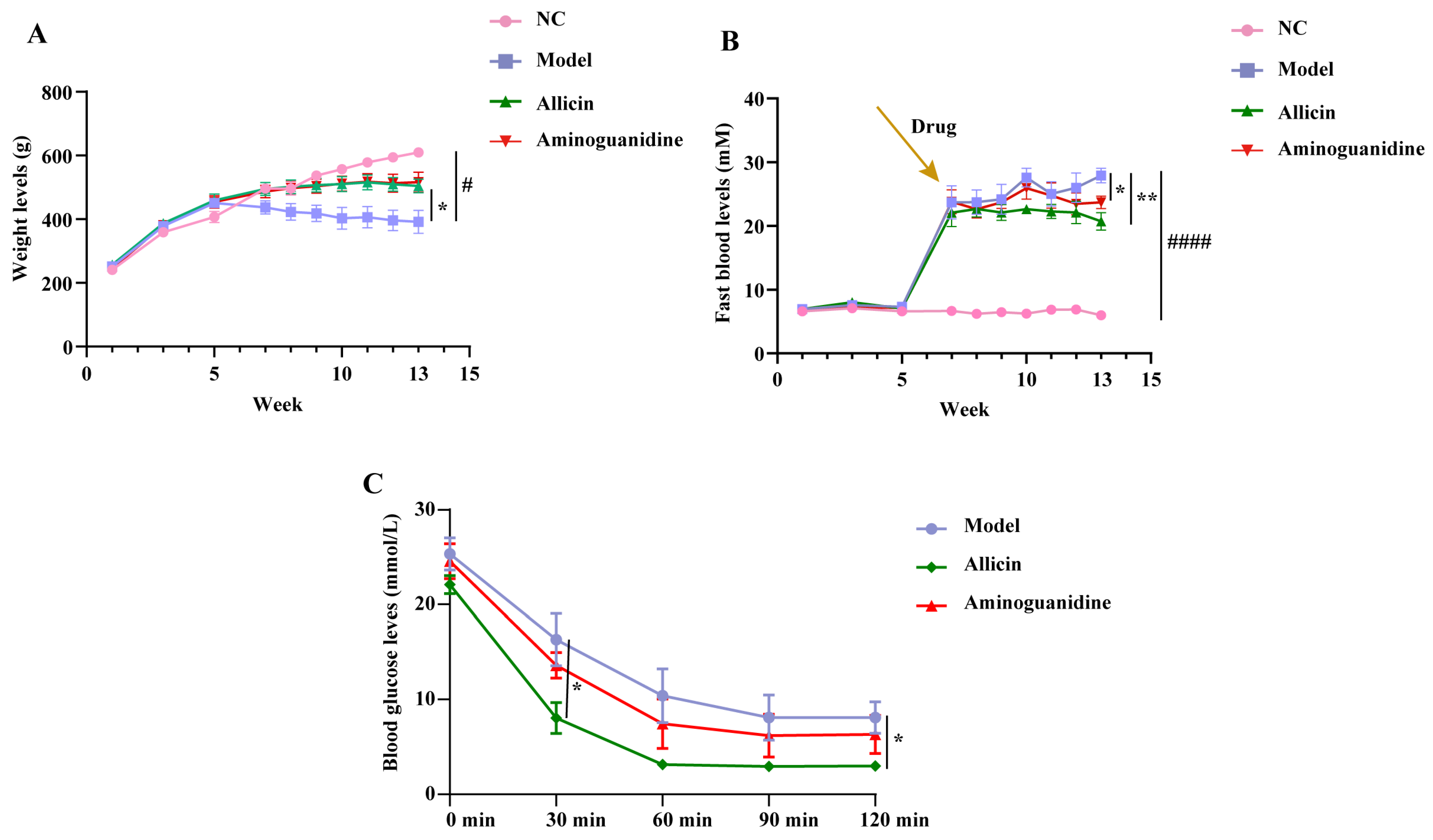
2.1. Allicin Alleviates Metabolic Dysregulation in Rats with STZ-Induced Diabetes
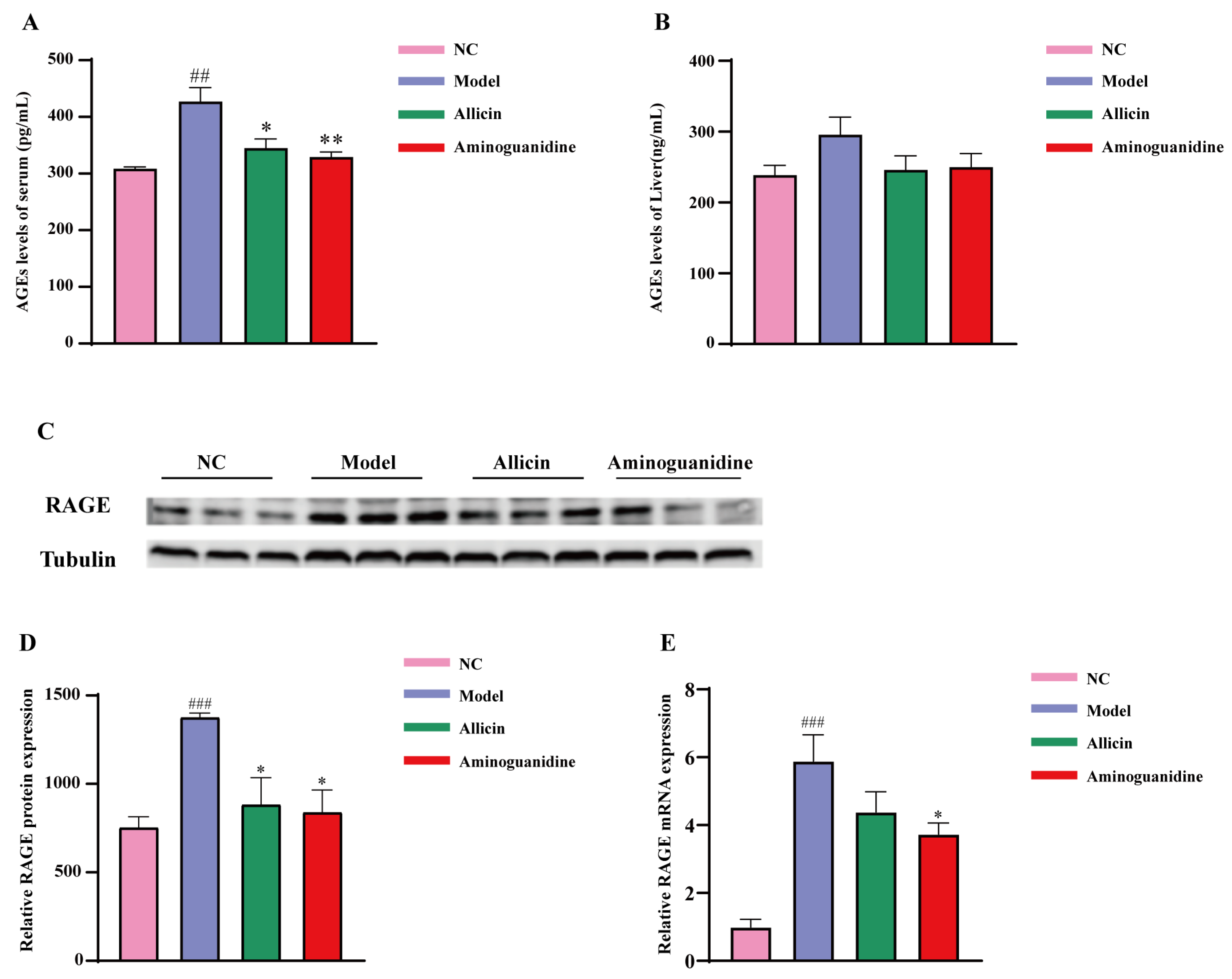
2.2. Allicin Reduces AGE Levels in Diabetic Rats
2.3. Allicin Alleviates Oxidative Stress in Rats
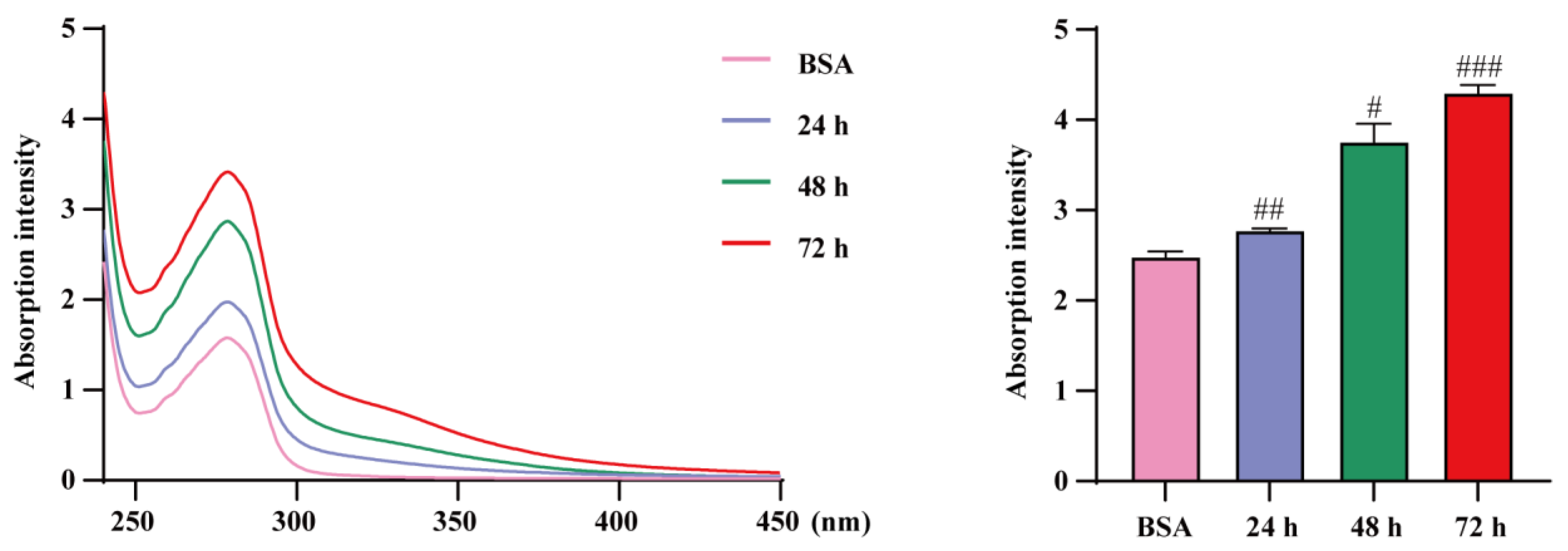
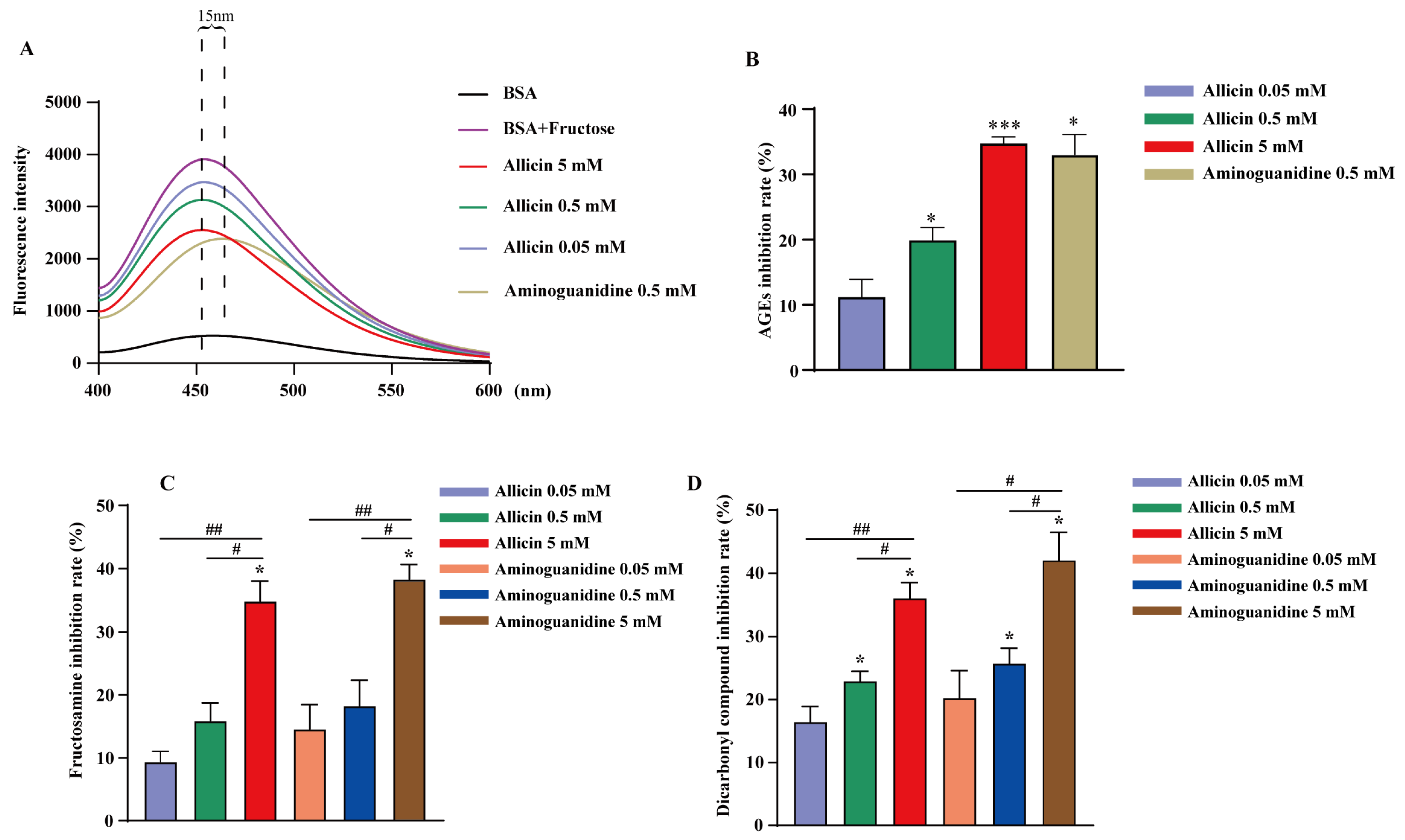
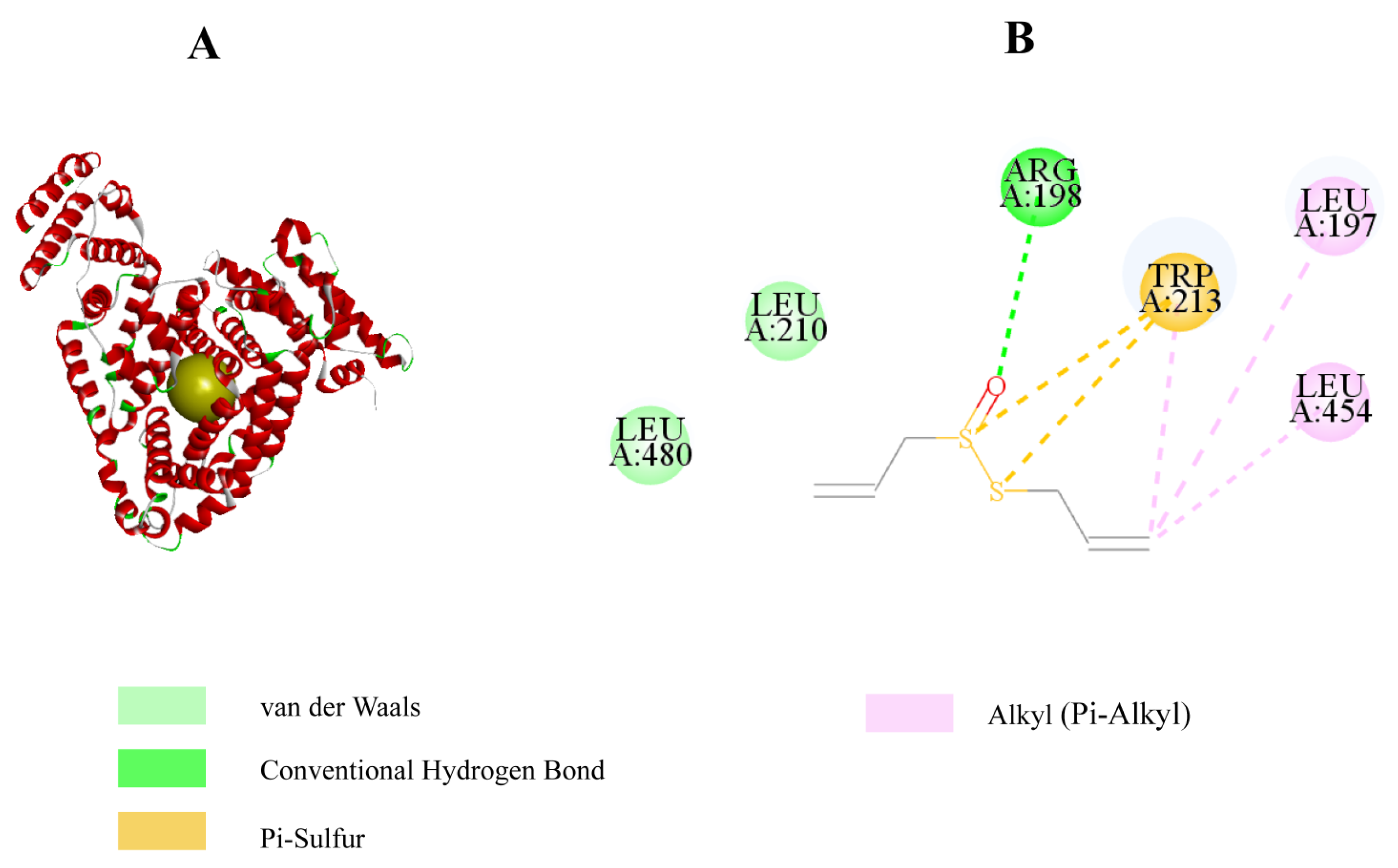
2.4. Allicin Inhibits the Process of AGE Formation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Experimental Design and Treatment
4.4. Fasting Blood Glucose and Body Weight
4.5. ITT
4.6. Serological Index Measurements
4.7. Determination of SOD, MDA, and GSH-PX
4.8. Serum and Liver AGE Determination
4.9. Inhibition of AGE Formation by Allicin In Vitro
4.10. Determination of the Inhibition Rate of Fructosamine
4.11. Determination of the Inhibition Rate of α-Dicarbonyl Compounds
4.12. Molecular Docking of Allicin with BSA
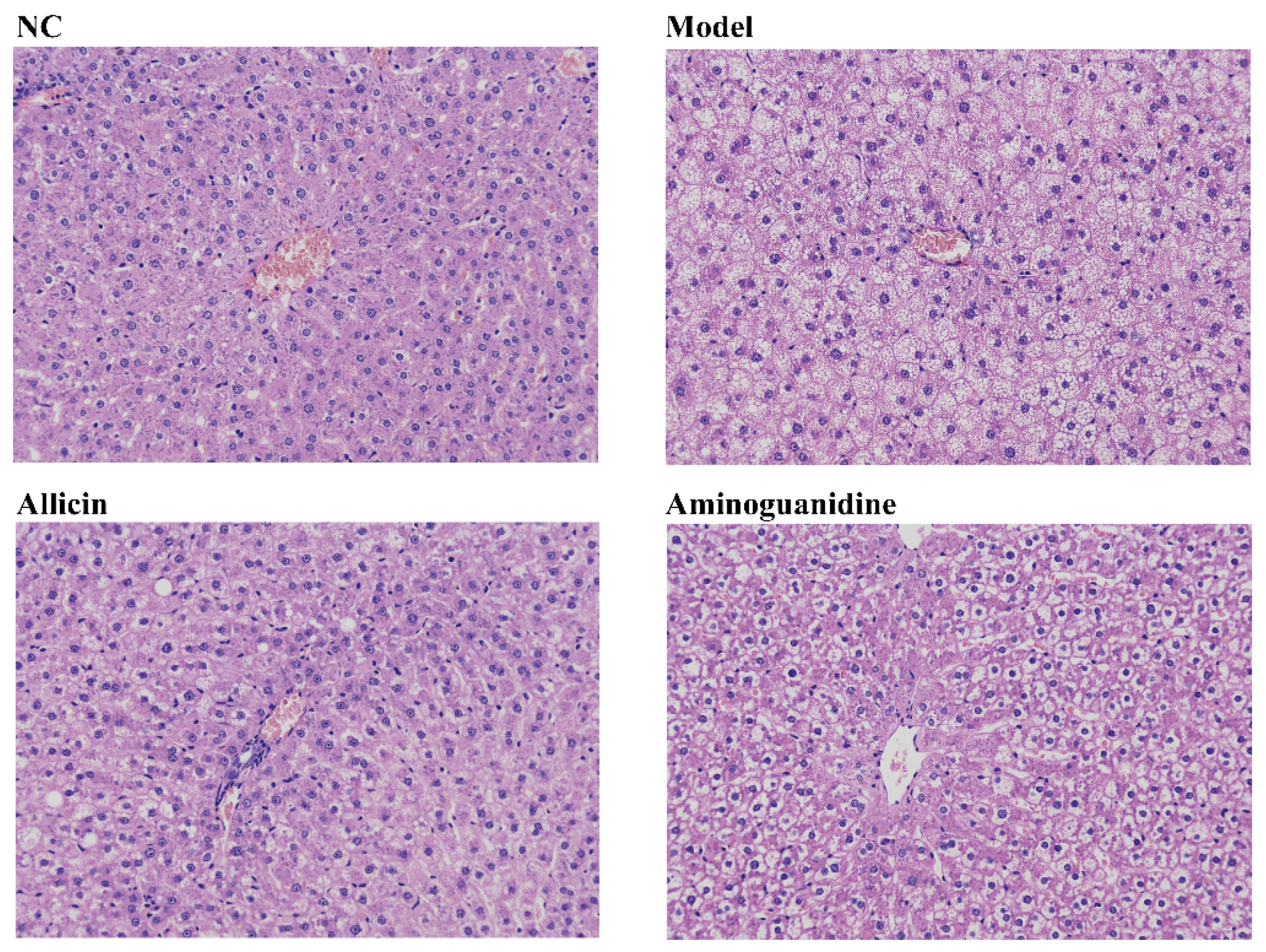
4.13. Staining of Liver Tissue Sections
4.14. Real-Time PCR
4.15. Western Blot
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Uribarri, J.; Woodruff, S.; Goodman, S.; Cai, W.; Chen, X.; Pyzik, R.; Yong, A.; Striker, G.E.; Vlassara, H. Advanced glycation end products in foods and a practical guide to their reduction in the diet. J. Am. Diet. Assoc. 2010, 110, 911–916.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deluyker, D.; Evens, L.; Bito, V. Advanced glycation end products (AGEs) and cardiovascular dysfunction: Focus on high molecular weight AGEs. Amino Acids 2017, 49, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wu, C.; Yen, G. Perspective of advanced glycation end products on human health. J. Agric. Food Chem. 2018, 66, 2065–2070. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, S. An abnormal hemoglobin in red cells of diabetics. Clin. Chim. Acta 1968, 22, 296. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9 th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Ogurtsova, K.; Da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Unnikrishnan, R.; Anjana, R.M.; Mohan, V. Diabetes mellitus and its complications in India. Nat. Rev. Endocrinol. 2016, 12, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Vlassara, H.; Striker, G.E. AGE restriction in diabetes mellitus: A paradigm shift. Nat. Rev. Endocrinol. 2011, 7, 526–539. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhao, C.; Zhang, X.H.; Zheng, F.; Cai, W.; Vlassara, H.; Ma, Z.A. Advanced glycation end products inhibit glucose-stimulated insulin secretion through nitric oxide-dependent inhibition of cytochrome c oxidase and adenosine triphosphate synthesis. Endocrinology 2009, 150, 2569–2576. [Google Scholar] [CrossRef]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bügel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef]
- Hyogo, H.; Yamagishi, S. Advanced glycation end products (AGEs) and their involvement in liver disease. Curr. Pharm. Des. 2008, 14, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.; Herath, C.B.; Jia, Z.; Andrikopoulos, S.; Brown, B.E.; Davies, M.; Rivera, L.R.; Furness, J.B.; Forbes, J.; Angus, P.W. Dietary advanced glycation end-products aggravate non-alcoholic fatty liver disease. World J. Gastroenterol. 2016, 22, 8026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šebeková, K.; Kupčová, V.; Schinzel, R.; Heidland, A. Markedly elevated levels of plasma advanced glycation end products in patients with liver cirrhosis-amelioration by liver transplantation. J. Hepatol. 2002, 36, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Yagmur, E.; Tacke, F.; Weiss, C.; Lahme, B.; Manns, M.P.; Kiefer, P.; Trautwein, C.; Gressner, A.M. Elevation of Nε-(carboxymethyl)lysine-modified advanced glycation end products in chronic liver disease is an indicator of liver cirrhosis. Clin. Biochem. 2006, 39, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Hamed, A.E.; Elwan, N.; Naguib, M.; Elwakil, R.; Esmat, G.; El Kassas, M.; Abd-Elsalam, S.; Moussa, S. Diabetes association with liver diseases: An overview for clinicians. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 274. [Google Scholar] [CrossRef]
- Song, Q.; Liu, J.; Dong, L.; Wang, X.; Zhang, X. Novel advances in inhibiting advanced glycation end product formation using natural compounds. Biomed. Pharmacother. 2021, 140, 111750. [Google Scholar] [CrossRef]
- Muthenna, P.; Akileshwari, C.; Saraswat, M.; Reddy, G.B. Inhibition of advanced glycation end-product formation on eye lens protein by rutin. Br. J. Nutr. 2012, 107, 941–949. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Singh, S.P.; Pandey, A.R.; Ansari, A.; Ahmad, S.; Sashidhara, K.V.; Tamrakar, A.K. Flavonoids from Polyalthia longifolia prevents advanced glycation end products formation and protein oxidation aligned with fructose-induced protein glycation. Nat. Prod. Res. 2019, 35, 2921–2925. [Google Scholar] [CrossRef]
- Yoon, S.; Shim, S. Inhibitory effect of polyphenols in Houttuynia cordata on advanced glycation end-products (AGEs) by trapping methylglyoxal. LWT—Food Sci. Technol. 2015, 61, 158–163. [Google Scholar] [CrossRef]
- Shi, X.; Zhou, X.; Chu, X.; Wang, J.; Xie, B.; Ge, J.; Guo, Y.; Li, X.; Yang, G. Allicin improves metabolism in high-fat diet-induced obese mice by modulating the gut microbiota. Nutrients 2019, 11, 2909. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhao, J.; Zhao, M.; Wang, K. Allicin activates autophagic cell death to alleviate the malignant development of thyroid cancer. Exp. Ther. Med. 2018, 15, 3537–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Nan, B.; Wu, X.; Yan, H.; Yuan, Y. Allicin alleviates acrylamide-induced oxidative stress in BRL-3A cells. Life Sci. 2019, 231, 116550. [Google Scholar] [CrossRef] [PubMed]
- García-Trejo, E.M.A.; Arellano-Buendía, A.S.; Argüello-García, R.; Loredo-Mendoza, M.L.; García-Arroyo, F.E.; Arellano-Mendoza, M.G.; Castillo-Hernández, M.C.; Guevara-Balcázar, G.; Tapia, E.; Sánchez-Lozada, L.G.; et al. Effects of allicin on hypertension and cardiac function in chronic kidney disease. Oxidative Med. Cell. Longev. 2016, 2016, 3850402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, X.-B.; Zhang, X.-M.; Wang, S.; Lan, Y.; Zhang, S.; Huo, L.-Y. Allicin as add-on therapy for Helicobacter pylori infection: A systematic review and meta-analysis. World J. Gastroenterol. 2019, 25, 6025–6040. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Lan, H.; Wang, W. Effect of garlic supplement in the management of type 2 diabetes mellitus (T2DM): A meta-analysis of randomized controlled trials. Food Nutr. Res. 2017, 61, 1377571. [Google Scholar] [CrossRef] [Green Version]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’Nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxidative Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.N.G.D.S.; Paula, D.P.; de Araujo, B.P.; Fonseca, M.D.J.M.D.; Diniz, M.D.F.H.S.; Daliry, A.; Griep, R.H. Advanced glycation end product: A potential biomarker for risk stratification of non-alcoholic fatty liver disease in ELSA-Brasil study. World J. Gastroenterol. 2021, 27, 4913–4928. [Google Scholar] [CrossRef]
- Srivastava, R.A.K. Life-style-induced metabolic derangement and epigenetic changes promote diabetes and oxidative stress leading to NASH and atherosclerosis severity. J. Diabetes Metab. Disord. 2018, 17, 381–391. [Google Scholar] [CrossRef]
- Li, C.L.; Liu, X.H.; Qiao, Y.; Ning, L.N.; Li, W.J.; Sun, Y.S.; Liu, D.S.; Gao, W.; Ma, C.M. Allicin alleviates inflammation of diabetic macroangiopathy via the Nrf2 and NF-kB pathway. Eur. J. Pharmacol. 2020, 876, 173052. [Google Scholar] [CrossRef]
- Shang, A.; Cao, S.-Y.; Xu, X.-Y.; Gan, R.-Y.; Tang, G.-Y.; Corke, H.; Mavumengwana, V.; Li, H.-B. Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef]
- Lisiswanti, R.; Haryanto, F.P. Allicin pada bawang putih (Allium sativum) sebagai terapi alternatif diabetes melitus tipe 2. J. Major. 2017, 6, 33–38. [Google Scholar]
- Arellano Buendía, A.S.; Tostado González, M.; Sánchez Reyes, O.; García Arroyo, F.E.; Argüello García, R.; Tapia, E.; Sánchez Lozada, L.G.; Osorio Alonso, H. Immunomodulatory effects of the nutraceutical garlic derivative allicin in the progression of diabetic nephropathy. Int. J. Mol. Sci. 2018, 19, 3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlassara, H.; Uribarri, J. Advanced glycation end products (AGE) and diabetes: Cause, effect, or both? Curr. Diabetes Rep. 2014, 14, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; He, X.; Sheng, Y.; Xu, J.; Yang, C.; Zheng, S.; Liu, J.; Li, H.; Ge, J.; Yang, M.; et al. Allicin regulates energy homeostasis through brown adipose tissue. iScience 2020, 23, 101113. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Buendía, A.S.; Castañeda-Lara, L.G.; Loredo-Mendoza, M.L.; García-Arroyo, F.E.; Rojas-Morales, P.; Argüello-García, R.; Juárez-Rojas, J.G.; Tapia, E.; Pedraza-Chaverri, J.; Sánchez-Lozada, L.G.; et al. Effects of allicin on pathophysiological mechanisms during the progression of nephropathy associated to diabetes. Antioxidants 2020, 9, 1134. [Google Scholar] [CrossRef] [PubMed]
- Sourris, K.C.; Watson, A.; Jandeleit-Dahm, K. Inhibitors of advanced glycation end product (AGE) formation and accumulation. Handb. Exp. Pharmacol. 2021, 264, 395–423. [Google Scholar] [PubMed]
- Ali, M.; Al-Qattan, K.; Al-Enezi, F.; Khanafer, R.; Mustafa, T. Effect of allicin from garlic powder on serum lipids and blood pressure in rats fed with a high cholesterol diet. Prostaglandins Leukot. Essent. Fat. Acids 2000, 62, 253–259. [Google Scholar] [CrossRef]
- Cheng, B.; Li, T.; Li, F. Use of network pharmacology to investigate the mechanism by which allicin ameliorates lipid metabolism disorder in HepG2 cells. Evid.-Based Complement. Altern. Med. 2021, 2021, 3956504. [Google Scholar] [CrossRef]
- You, M.; Arteel, G.E. Effect of ethanol on lipid metabolism. J. Hepatol. 2019, 70, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Vlassara, H.; Striker, G.E. Advanced glycation endproducts in diabetes and diabetic complications. Endocrinol. Metab. Clin. N. Am. 2013, 42, 697–719. [Google Scholar] [CrossRef]
- Ceretta, L.B.; Réus, G.Z.; Abelaira, H.M.; Ribeiro, K.F.; Zappellini, G.; Felisbino, F.F.; Steckert, A.V.; Dal-Pizzol, F.; Quevedo, J. Increased oxidative stress and imbalance in antioxidant enzymes in the brains of alloxan-induced diabetic rats. Exp. Diabetes Res. 2012, 2012, 302682. [Google Scholar] [CrossRef] [PubMed]
- Patche, J.; Girard, D.; Catan, A.; Boyer, F.; Dobi, A.; Planesse, C.; Diotel, N.; Guerin-Dubourg, A.; Baret, P.; Bravo, S.B.; et al. Diabetes-induced hepatic oxidative stress: A new pathogenic role for glycated albumin. Free Radic. Biol. Med. 2017, 102, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Schalkwijk, C.G.; Miyata, T. Early- and advanced non-enzymatic glycation in diabetic vascular complications: The search for therapeutics. Amino Acids 2012, 42, 1193–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, G. The role of polyphenols in modern nutrition. Nutr. Bull. 2017, 42, 226–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, J.M.; Shahab, U.; Tabrez, S.; Lee, E.J.; Choi, I.; Ahmad, S. Quercetin as a finer substitute to aminoguanidine in the inhibition of glycation products. Int. J. Biol. Macromol. 2015, 77, 188–192. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.H.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; He, Y.L.; Wang, S.; He, Y.; Wang, W.Y.; Li, Q.J.; Cao, X.Y. Ferulic acid inhibits advanced glycation end products (AGEs) formation and mitigates the AGEs-induced inflammatory response in HUVEC cells. J. Funct. Foods 2018, 48, 19–26. [Google Scholar] [CrossRef]
- Hou, Y.; Ding, W.; Wu, P.; Liu, C.; Ding, L.; Liu, J.; Wang, X. Adipose-derived stem cells alleviate liver injury induced by type 1 diabetes mellitus by inhibiting mitochondrial stress and attenuating inflammation. Stem Cell Res. Ther. 2022, 13, 132. [Google Scholar] [CrossRef]
- Wang, S.-H.; Chang, J.-C.; Pokkaew, R.; Lee, J.-F.; Chiou, R.Y.-Y. Modified fast procedure for the detection and screening of antiglycative phytochemicals. J. Agric. Food Chem. 2011, 59, 6906–6912. [Google Scholar] [CrossRef]
- Zeng, L.; Ding, H.; Hu, X.; Zhang, G.; Gong, D. Galangin inhibits α-glucosidase activity and formation of non-enzymatic glycation products. Food Chem. 2019, 271, 70–79. [Google Scholar] [CrossRef]
- Baker, J.R.; Zyzak, D.V.; Thorpe, S.R.; Baynes, J.W. Chemistry of the fructosamine assay: D-glucosone is the product of oxidation of Amadori compounds. Clin. Chem. 1994, 40, 1950–1955. [Google Scholar] [CrossRef] [PubMed]
- Wells-Knecht, K.J.; Zyzak, D.V.; Litchfield, J.E.; Thorpe, S.R.; Baynes, J.W. Mechanism of autoxidative glycosylation: Identification of glyoxal and arabinose as intermediates in the autoxidative modification of proteins by glucoset. Biochemistry 1995, 34, 3704–3709. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | NC | Model | Allicin | Aminoguanidine |
|---|---|---|---|---|
| TG (mmol/L) | 2.66 ± 0.54 | 8.52 ± 1.65 ## | 1.225 ± 0.32 ** | 1.85 ± 0.59 ** |
| TC (mmol/L) | 3.04 ± 0.13 | 3.42 ± 0.40 | 2.35 ± 0.19 * | 3.03 ± 0.46 |
| HDL-C (mmol/L) | 1. 64 ± 0.05 | 1.48 ± 0.13 | 1.60 ± 0.15 | 1.84 ± 0.27 |
| LDL-C (mmol/L) | 0.46 ± 0.05 | 1.00 ± 0.20 # | 0.29 ± 0.04 ** | 0.41 ± 0.09 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Song, Q.; Zhang, X.; Yan, Y.; Wang, X. Allicin Alleviates Diabetes Mellitus by Inhibiting the Formation of Advanced Glycation End Products. Molecules 2022, 27, 8793. https://doi.org/10.3390/molecules27248793
Li L, Song Q, Zhang X, Yan Y, Wang X. Allicin Alleviates Diabetes Mellitus by Inhibiting the Formation of Advanced Glycation End Products. Molecules. 2022; 27(24):8793. https://doi.org/10.3390/molecules27248793
Chicago/Turabian StyleLi, Linzehao, Qinghe Song, Xiandang Zhang, Yan Yan, and Xiaolei Wang. 2022. "Allicin Alleviates Diabetes Mellitus by Inhibiting the Formation of Advanced Glycation End Products" Molecules 27, no. 24: 8793. https://doi.org/10.3390/molecules27248793