
Biotechnology and In Vitro Culture as an Alternative System for Secondary Metabolite Production
 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
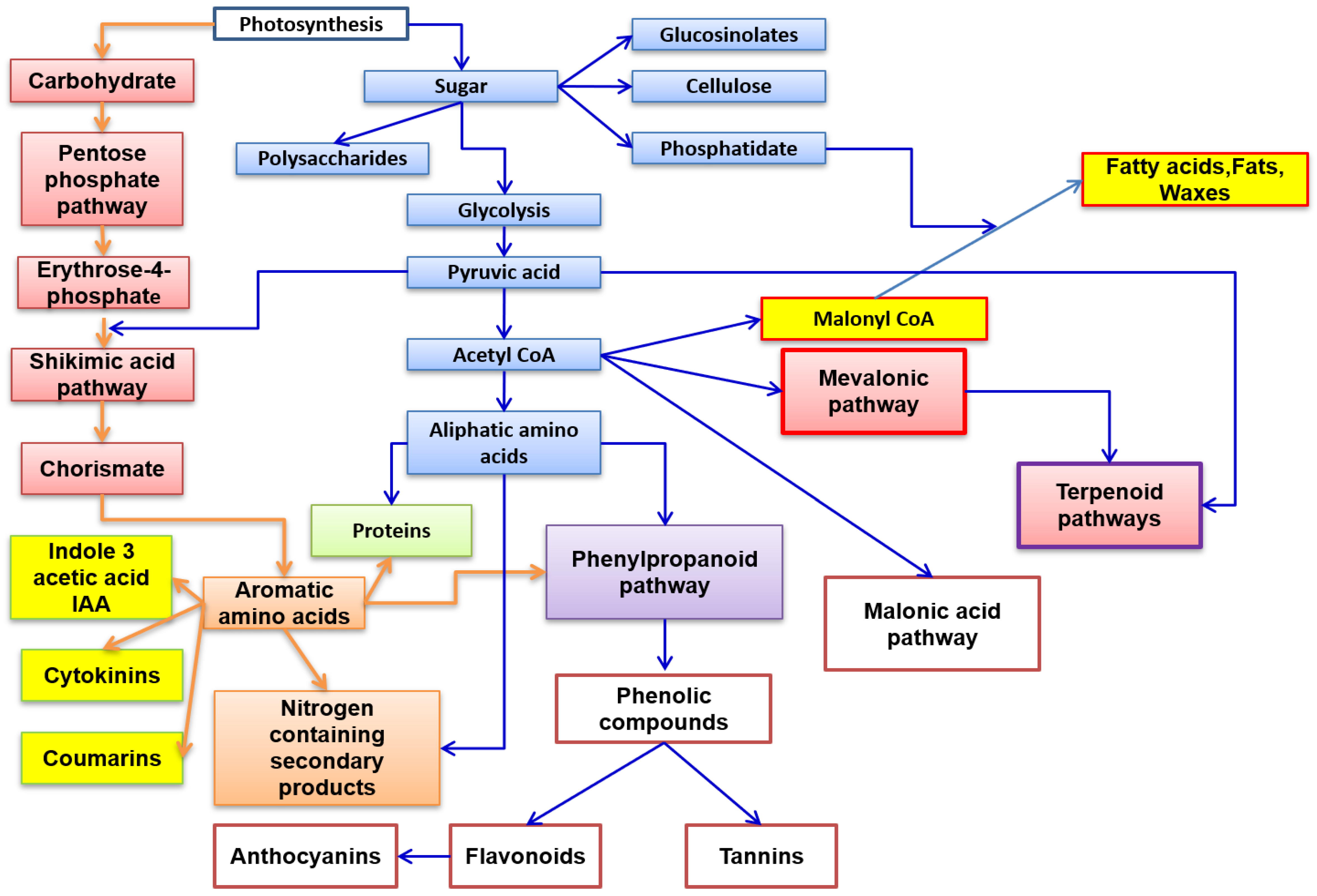
2. Plant Secondary Metabolites
3. Micropropagation as a Tool for the Production of Secondary Metabolites
4. The Importance of Cell and Suspension Culture in the Production of Plant Secondary Metabolites
5. Bioreactors: System for Large-Scale Production
6. Elicitation of In Vitro Products

{kind=link}
| Plant Species | Elicitor Factor | Culture System | Product | Key Findings | References |
|---|---|---|---|---|---|
| Abiotic elicitors | |||||
| Chelidonium majus | Methyl jasmonate (MJ) Salicylic acid (SA) | Cell suspension culture | Chelidonine, sanguinarine | Elicitation stimulated the expression of genes in the benzophenanthridine alkaloid biosynthetic pathway. | [177] |
| Ocimumbasilicum | Copper oxide (CuO) | Callus culture | Rosmarinic acid, chicoric acid, eugenol | Elicitation by nanoparticles stimulated the biosynthesis of the secondary metabolite. | [178] |
| Ocimumbasilicum | Salicylic acid (SA) + light regimes | Callus culture | Rosmarinic acid, chicoricacid, cyanidin, peonidin | Continuous light with SA increased the content of phenolic compounds and flavonoids, also antioxidant activity. | [178] |
| Coelogyne ovalis | Salicylic acid (SA) | Tissue culture | Flavonoids, anthocyanins, phenolic compounds | Elicitation stimulates chalcone synthase expression and secondary metabolites production. | [179] |
| Papaver orientale | Methyl jasmonate (MJ), salicylic acid (SA) | Hairy root culture | Thebaine, morphine, codeine | Expression of morphinan biosynthetic genes was significantly upregulated with MJ and SA. MJ and SA elicitation enhanced thebaine, morphine, and codeine biosynthesis. | [172] |
| Crocus sativus | Ultrasonic waves | Cell suspension culture | Safranal, crocin | Ultrasonic treatment acted as an effective mechanical stimulus on the production of secondary metabolites in suspension cultures. | [180] |
| Gymnemasylvestre | Sodium nitroprusside (SNP) | Cell suspension culture | Deacylgymnemic acid, gymnemagenin, gymnemic acid XVII | Significant improvement in the content of gymnemic acids in cell suspension cultures of G. sylvestre. | [161] |
| Momordica charantia | Silver nanoparticles (AgNPs) | Cell suspension culture | Hydroxybenzoic, hydroxycinnamic | The significant increase in bioactive compounds as well as pharmacological activities was enhanced by the application of elicitation. | [181] |
| Biotic elicitors | |||||
| Corylus avellana | Chaetommiuglobosum | Cell suspension cultures | Paclitaxel | Increased extracellular portion of paclitaxel (44.0%). | [170] |
| Bletilla striata | Byssochlamys spectabilis | Tissue culture | Total phenolic content | Increased total phenolic compounds. | [182] |
| Panax ginseng | Aspergillus niger | Adventitious root culture | Ginsenosides | A. Niger triggered the defense response of plants and enhanced the accumulation of nitric oxide (NO), SA, and JA. Significantly upregulated the gene expression of terpenoid biosynthesis. | [183] |
| Panax ginseng | Alternaria panax | Adventitious root culture | Ginsenosides | Nitric oxide (NO), putrescine (Put), and hydrogen peroxide (H2O2) are involved in regulating ginsenoside synthesis in fungal elicitor-treated Adventitious root of P. ginseng. | [184] |
| Trichosanthescucumerina | Chitosan | Callus and suspension culture | Bryonolic acid | Callus and suspension cultures presented higher levels of Bryonolic acid than the natural roots ones. | [185] |
| Psammosilenetunicoides | Chitosan | Hairy root culture | Quillaic acid, gypsogenin, gypsogenin 3-O-β-D-glucuronopyranoside | Chitosan elicitor promotes triterpenoid saponin biosynthesis by enhancing antioxidant activities and differential gene expression. | [175] |
| Iberis amara | Chitosan | Cell suspension culture | Total phenol, flavonoid, flavonol, anthocyanin | Chitosan elicitor promotes phenolic compounds’ biosynthesis without genetic modifications in medicinal herbs. | [186] |
| Plumbago zeylanica | Chitosan and yeast extract | Root callus | Plumbagin | Increase of 12.08-fold plumbagin content compared to control. | [187] |
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harborne, J.B. Classes and Functions of Secondary Products from Plants. Chem. Plants 1999, 26, 1–25. [Google Scholar]
- Aboukhalid, K.; Lamiri, A.; Agacka-Moldoch, M.; Doroszewska, T.; Douaik, A.; Bakha, M.; Casanova, J.; Tomi, F.; Machon, N.; Faiz, C.A. Chemical Polymorphism of Origanum Compactum Grown in All Natural Habitats in Morocco. Chem. Biodivers. 2016, 13, 1126–1139. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-C.; Bao, H.-Y.; Liu, Y.-Y.; Nie, Y.-Y.; Yang, J.-M.; Hong, P.-Z.; Zhang, Y. Depsidone Derivatives and a Cyclopeptide Produced by Marine Fungus Aspergillus unguis under Chemical Induction and by Its Plasma Induced Mutant. Molecules 2018, 23, 2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New Concepts in Nutraceuticals as Alternative for Pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487. [Google Scholar] [PubMed]
- Elazzouzi, H.; Soro, A.; Elhilali, F.; Bentayeb, A.; El Belghiti, M.A.; Zair, T. Phytochemical Study of Anacyclus pyrethrum (L.) of Middle Atlas (Morocco), and in Vitro Study of Antibacterial Activity of Pyrethrum. Adv. Nat. Appl. Sci. 2014, 8, 131–141. [Google Scholar]
- Kikowska, M.; Thiem, B.; Szopa, A.; Ekiert, H. Accumulation of Valuable Secondary Metabolites: Phenolic Acids and Flavonoids in Different in Vitro Systems of Shoot Cultures of the Endangered Plant Species-Eryngium alpinum L. Plant Cell Tissue Organ Cult. 2020, 141, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Gurib-Fakim, A. Medicinal Plants: Traditions of Yesterday and Drugs of Tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef] [PubMed]
- Isah, T. Anticancer Alkaloids from Trees: Development into Drugs. Pharmacogn. Rev. 2016, 10, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Paek, K.-Y. Bioreactor Culture of Shoots and Somatic Embryos of Medicinal Plants for Production of Bioactive Compounds. Prod. Biomass Bioact. Compd. Using Bioreact. Technol. 2014, 3, 337–368. [Google Scholar]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of Plant Secondary Metabolites: A Historical Perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Bouyahya, A.; Abrini, J.; Bakri, Y.; Dakka, N. Essential Oils as Anticancer Agents: News on Mode of Action. Phytothérapie 2016, 146, 1–14. [Google Scholar]
- Bouyahya, A.; Guaouguaou, F.-E.; El Omari, N.; El Menyiy, N.; Balahbib, A.; El-Shazly, M.; Bakri, Y. Anti-Inflammatory and Analgesic Properties of Moroccan Medicinal Plants: Phytochemistry, in Vitro and in Vivo Investigations, Mechanism Insights, Clinical Evidences and Perspectives. J. Pharm. Anal. 2021, 12, 35–57. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; El Omari, N.; Elmenyiy, N.; Guaouguaou, F.-E.; Balahbib, A.; Belmehdi, O.; Salhi, N.; Imtara, H.; Mrabti, H.N.; El-Shazly, M. Moroccan Antidiabetic Medicinal Plants: Ethnobotanical Studies, Phytochemical Bioactive Compounds, Preclinical Investigations, Toxicological Validations and Clinical Evidences; Challenges, Guidance and Perspectives for Future Management of Diabetes Worldwide. Trends Food Sci. Technol. 2021, 115, 147–254. [Google Scholar]
- Bulugahapitiya, V.P. Plants Based Natural Products; University of Ruhuna: Fribourg, Switzerland, 2013. [Google Scholar]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for Extraction and Isolation of Natural Products: A Comprehensive Review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary Metabolism of Pharmaceuticals in the Plant in Vitro Cultures: Strategies, Approaches, and Limitations to Achieving Higher Yield. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 239–265. [Google Scholar] [CrossRef]
- Nalawade, S.M.; Tsay, H.-S. In Vitro Propagation of Some Important Chinese Medicinal Plants and Their Sustainable Usage. Vitr. Cell. Dev. Biol.-Plant 2004, 40, 143–154. [Google Scholar] [CrossRef]
- Gaosheng, H.; Jingming, J. Production of Useful Secondary Metabolites through Regulation of Biosynthetic Pathway in Cell and Tissue Suspension Culture of Medicinal Plants. Recent Adv. Plant Vitr. Cult. 2012, 10, 53038. [Google Scholar]
- Gonçalves, S.; Romano, A. Production of Plant Secondary Metabolites by Using Biotechnological Tools. In Secondary Metabolites—Sources and Applications; IntechOpen: London, UK, 2018; pp. 81–99. [Google Scholar]
- Guerriero, G.; Berni, R.; Muñoz-Sanchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.A.; Cantini, C.; Cai, G.; Hausman, J.-F. Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulabagal, V.; Tsay, H.-S. Plant Cell Cultures-an Alternative and Efficient Source for the Production of Biologically Important Secondary Metabolites. Int. J. Appl. Sci. Eng. 2004, 2, 29–48. [Google Scholar]
- Yue, W.; Ming, Q.; Lin, B.; Rahman, K.; Zheng, C.-J.; Han, T.; Qin, L. Medicinal Plant Cell Suspension Cultures: Pharmaceutical Applications and High-Yielding Strategies for the Desired Secondary Metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef]
- Yahya, N.A.; Attan, N.; Wahab, R.A. An Overview of Cosmeceutically Relevant Plant Extracts and Strategies for Extraction of Plant-Based Bioactive Compounds. Food Bioprod. Process. 2018, 112, 69–85. [Google Scholar] [CrossRef]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A.; Ghafoor, K.; Norulaini, N.A.N.; Omar, A.K.M. Techniques for Extraction of Bioactive Compounds from Plant Materials: A Review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Azwanida, N.N. A Review on the Extraction Methods Use in Medicinal Plants, Principle, Strength and Limitation. Med. Aromat Plants 2015, 4, 1000196. [Google Scholar]
- Belwal, T.; Ezzat, S.M.; Rastrelli, L.; Bhatt, I.D.; Daglia, M.; Baldi, A.; Devkota, H.P.; Orhan, I.E.; Patra, J.K.; Das, G. A Critical Analysis of Extraction Techniques Used for Botanicals: Trends, Priorities, Industrial Uses and Optimization Strategies. TrAC Trends Anal. Chem. 2018, 100, 82–102. [Google Scholar] [CrossRef]
- Jovanović, A.A.; DJordjević, V.B.; Zdunić, G.M.; Pljevljakušić, D.S.; Šavikin, K.P.; Godjevac, D.M.; Bugarski, B.M. Optimization of the Extraction Process of Polyphenols from Thymus serpyllum L. Herb Using Maceration, Heat-and Ultrasound-Assisted Techniques. Sep. Purif. Technol. 2017, 179, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Dzah, C.S.; Duan, Y.; Zhang, H.; Wen, C.; Zhang, J.; Chen, G.; Ma, H. The Effects of Ultrasound Assisted Extraction on Yield, Antioxidant, Anticancer and Antimicrobial Activity of Polyphenol Extracts: A Review. Food Biosci. 2020, 35, 100547. [Google Scholar] [CrossRef]
- Haloui, I.; Meniai, A.-H. Supercritical CO2 Extraction of Essential Oil from Algerian Argan (Argania spinosa L.) Seeds and Yield Optimization. Int. J. Hydrog. Energy 2017, 42, 12912–12919. [Google Scholar] [CrossRef]
- Meireles, M.A.A. Supercritical Extraction from Solid: Process Design Data (2001–2003). Curr. Opin. Solid State Mater. Sci. 2003, 7, 321–330. [Google Scholar] [CrossRef]
- Souza, M.A.; Guzatti, J.G.; Martello, R.H.; Schindler, M.S.; Calisto, J.F.; Morgan, L.V.; Aguiar, G.P.; Locateli, G.; Scapinello, J.; Müller, L.G. Supercritical CO2 Extraction of Aloysia Gratissima Leaves and Evaluation of Anti-Inflammatory Activity. J. Supercrit. Fluids 2020, 159, 104753. [Google Scholar] [CrossRef]
- Rahmalia, W.; Fabre, J.-F.; Mouloungui, Z. Effects of Cyclohexane/Acetone Ratio on Bixin Extraction Yield by Accelerated Solvent Extraction Method. Procedia Chem. 2015, 14, 455–464. [Google Scholar] [CrossRef]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A Biotechnological Tool for Enhanced Production of Secondary Metabolites in Hairy Root Cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [Green Version]
- Kawka, M.; Bubko, I.; Koronkiewicz, M.; Gruber-Bzura, B.; Graikou, K.; Chinou, I.; Jeziorek, M.; Pietrosiuk, A.; Syklowska-Baranek, K. Polyurethane Foam Rafts Supported in Vitro Cultures of Rindera Graeca Roots for Enhanced Production of Rinderol, Potent Proapoptotic Naphthoquinone Compound. Int. J. Mol. Sci. 2021, 23, 56. [Google Scholar] [CrossRef]
- Nowak, B.; Kawka, M.; Wierzchowski, K.; Syklowska-Baranek, K.; Pilarek, M. MTMS-Based Aerogel Constructs for Immobilization of Plant Hairy Roots: Effects on Proliferation of Rindera Graeca Biomass and Extracellular Secretion of Naphthoquinones. J. Funct. Biomater. 2021, 12, 19. [Google Scholar] [CrossRef] [PubMed]
- Syklowska-Baranek, K.; Rymaszewski, W.; Gawel, M.; Rokicki, P.; Pilarek, M.; Grech-Baran, M.; Hennig, J.; Pietrosiuk, A. Comparison of Elicitor-Based Effects on Metabolic Responses of Taxus× Media Hairy Roots in Perfluorodecalin-Supported Two-Phase Culture System. Plant Cell Rep. 2019, 38, 85–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syklowska-Baranek, K.; Pilarek, M.; Cichosz, M.; Pietrosiuk, A. Liquid Perfluorodecalin Application for in Situ Extraction and Enhanced Naphthoquinones Production in Arnebia Euchroma Cell Suspension Cultures. Appl. Biochem. Biotechnol. 2014, 172, 2618–2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H. Discovery and Resupply of Pharmacologically Active Plant-Derived Natural Products: A Review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oladimeji, A.V.; Valan, M.F. HPLC Techniques for Phytochemistry. IJCS 2020, 8, 2590–2596. [Google Scholar] [CrossRef]
- Nisar, B.; Sultan, A.; Rubab, S.L. Comparison of Medicinally Important Natural Products versus Synthetic Drugs-a Short Commentary. Nat. Prod. Chem. Res 2018, 6, 308. [Google Scholar] [CrossRef]
- Stratton, C.F.; Newman, D.J.; Tan, D.S. Cheminformatic Comparison of Approved Drugs from Natural Product versus Synthetic Origins. Bioorganic Med. Chem. Lett. 2015, 25, 4802–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hann, M.M.; Leach, A.R.; Harper, G. Molecular Complexity and Its Impact on the Probability of Finding Leads for Drug Discovery. J. Chem. Inf. Comput. Sci. 2001, 41, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Selzer, P.; Roth, H.-J.; Ertl, P.; Schuffenhauer, A. Complex Molecules: Do They Add Value? Curr. Opin. Chem. Biol. 2005, 9, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordell, G.A. Sustainable Medicines and Global Health Care. Planta Med. 2011, 77, 1129–1138. [Google Scholar] [CrossRef] [Green Version]
- Efferth, T. Biotechnology Applications of Plant Callus Cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Lautie, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Villarreal, M.; Howat, S.; Hong, S.; Jang, M.O.; Jin, Y.-W.; Lee, E.-K.; Loake, G.J. Plant Cell Culture Strategies for the Production of Natural Products. BMB Rep. 2016, 49, 149. [Google Scholar] [CrossRef] [PubMed]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant Tissue Culture as a Perpetual Source for Production of Industrially Important Bioactive Compounds. Biotechnol. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef] [PubMed]
- Eibl, R.; Meier, P.; Stutz, I.; Schildberger, D.; Hühn, T.; Eibl, D. Plant Cell Culture Technology in the Cosmetics and Food Industries: Current State and Future Trends. Appl. Microbiol. Biotechnol. 2018, 102, 8661–8675. [Google Scholar] [CrossRef] [PubMed]
- Alamgir, A.N.M. Cultivation of Herbal Drugs, Biotechnology, and in Vitro Production of Secondary Metabolites, High-Value Medicinal Plants, Herbal Wealth, and Herbal Trade. In Therapeutic Use of Medicinal Plants and Their Extracts: Volume 1; Springer: Berlin/Heidelberg, Germany, 2017; pp. 379–452. [Google Scholar]
- Tasheva, K.; Kosturkova, G. Role of Biotechnology for Protection of Endangered Medicinal Plants. In Environmental Biotechnology—New Approaches and Prospective Applications; IntechOpen: London, UK, 2013; pp. 235–238. [Google Scholar]
- Vanisree, M.; Lee, C.-Y.; Lo, S.-F.; Nalawade, S.M.; Lin, C.Y.; Tsay, H.-S. Studies on the Production of Some Important Secondary Metabolites from Medicinal Plants by Plant Tissue Cultures. Bot. Bull. Acad. Sin 2004, 45, 1–22. [Google Scholar]
- Sood, H. Production of Medicinal Compounds from Endangered and Commercially Important Medicinal Plants through Cell and Tissue Culture Technology for Herbal Industry. In Bioactive Compounds in Nutraceutical and Functional Food for Good Human Health; IntechOpen: London, UK, 2020. [Google Scholar]
- Davoodi, A.; Khoshvishkaie, E.; Azadbakht, M. Plant Cells Technology as an Effective Biotechnological Approach for High Scale Production of Pharmaceutical Natural Compounds; A Meta-Analysis Study. Pharm. Biomed. Res. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Karuppusamy, S. A Review on Trends in Production of Secondary Metabolites from Higher Plants by in Vitro Tissue, Organ and Cell Cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Rao, S.R.; Ravishankar, G.A. Plant Cell Cultures: Chemical Factories of Secondary Metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [PubMed]
- Kolewe, M.E.; Gaurav, V.; Roberts, S.C. Pharmaceutically Active Natural Product Synthesis and Supply via Plant Cell Culture Technology. Mol. Pharm. 2008, 5, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.; Singh, N.B.; Singh, A.; Hussain, I.; Niharika, K.; Yadav, V.; Bano, C.; Yadav, R.K.; Amist, N. Plant Secondary Metabolites Synthesis and Their Regulations under Biotic and Abiotic Constraints. J. Plant Biol. 2020, 63, 203–216. [Google Scholar] [CrossRef]
- Olivoto, T.; Nardino, M.; Carvalho, I.R.; Follmann, D.N.; Szareski, V.J.; Ferrari, M.; de Pelegrin, A.J.; de Souza, V.Q. Plant Secondary Metabolites and Its Dynamical Systems of Induction in Response to Environmental Factors: A Review. Afr. J. Agric. Res. 2017, 12, 71–84. [Google Scholar]
- Balandrin, M.F.; Klocke, J.A. Medicinal, Aromatic, and Industrial Materials from Plants. In Medicinal and Aromatic Plants I; Springer: Berlin/Heidelberg, Germany, 1988; pp. 3–36. [Google Scholar]
- Phillipson, J.D. Plants as source of valuable products. In Secondary Products from Plant Tissue Culture; Charlwood, B.V., Rhodes, M.J.C., Eds.; Clarendon Press: Oxford, UK, 1990; pp. 1–21. [Google Scholar]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: London, UK, 2019; p. 1. [Google Scholar]
- Arumugam, G.; Sinniah, U.R.; Swamy, M.K.; Lynch, P.T. Micropropagation and Essential Oil Characterization of Plectranthus Amboinicus (Lour.) Sprengel, an Aromatic Medicinal Plant. Vitr. Cell. Dev. Biol.-Plant 2020, 56, 491–503. [Google Scholar] [CrossRef]
- Galán-Ávila, A.; García-Fortea, E.; Prohens, J.; Herraiz, F.J. Development of a Direct in Vitro Plant Regeneration Protocol From Cannabis sativa L. Seedling Explants: Developmental Morphology of Shoot Regeneration and Ploidy Level of Regenerated Plants. Front. Plant Sci. 2020, 11, 645. [Google Scholar] [CrossRef]
- Singh, D.K.; Nirwan, S.; Babbar, S.B. Micropropagation of Anacyclus Pyrethrum and Chemical Profiling of the Regenerated Plants for Pellitorine, the Active Principle. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 122, 249–255. [Google Scholar] [CrossRef]
- Sottile, F.; Giuggioli, N.R.; Marinoni, D.T.; Peano, C.; Del Signore, M.B. Selection and Micropropagation of Valuable Caper Genotypes. Hortic. Sci. 2020, 47, 110–116. [Google Scholar] [CrossRef]
- Wróbel, T.; Dreger, M.; Wielgus, K.; Slomski, R. Modified Nodal Cuttings and Shoot Tips Protocol for Rapid Regeneration of Cannabis sativa L. J. Nat. Fibers 2020, 19, 536–545. [Google Scholar] [CrossRef]
- Bayarmaa, G.-A.; Lee, N.N.; Kang, H.D.; Oyuntsetseg, B.; Moon, H.K. Micropropagation of the Mongolian Medicinal Plant Zygophyllum Potaninii via Somatic Embryogenesis. Plant Biotechnol. Rep. 2018, 12, 187–194. [Google Scholar] [CrossRef]
- Bertero, V.G.; Beznec, A.; Faccio, P.; Auteri, M.; Arteaga, M.; Bonafede, M.; Bossio, E. High-Efficiency Direct Somatic Embryogenesis and Plant Regeneration from Leaf Base Explants of “Peperina” (Minthostachys Verticillata). Vitr. Cell. Dev. Biol.-Plant 2020, 56, 915–919. [Google Scholar] [CrossRef]
- Lema-Rumińska, J.; Kulus, D.; Tymoszuk, A.; Varejão, J.M.; Bahcevandziev, K. Profile of Secondary Metabolites and Genetic Stability Analysis in New Lines of Echinacea purpurea (L.) Moench Micropropagated via Somatic Embryogenesis. Ind. Crops Prod. 2019, 142, 111851. [Google Scholar] [CrossRef]
- Yamada, Y.; Shoyama, Y.; Nishioka, I.; Kohda, H.; Namera, A.; Okamoto, T. Clonal Micropropagation of Gentiana Scabra Bunge Var. Buergeri Maxim. and Examination of the Homogeneity Concerning the Gentiopicroside Content. Chem. Pharm. Bull. 1991, 39, 204–206. [Google Scholar] [CrossRef] [Green Version]
- Siahsar, B.; Rahimi, M.; Tavassoli, A.; Raissi, A. Application of Biotechnology in Production of Medicinal Plants. Am. Eurasian J. Agric Environ. Sci. 2011, 11, 439–444. [Google Scholar]
- Zahid, N.A.; Jaafar, H.Z.; Hakiman, M. Micropropagation of Ginger (Zingiber Officinale Roscoe)‘Bentong’and Evaluation of Its Secondary Metabolites and Antioxidant Activities Compared with the Conventionally Propagated Plant. Plants 2021, 10, 630. [Google Scholar] [CrossRef] [PubMed]
- Al Khateeb, W.; Kanaan, R.; El-Elimat, T.; Alu’datt, M.; Lahham, J.; El-Oqlah, A. In Vitro Propagation, Genetic Stability, and Secondary Metabolite Analysis of Wild Lavender (Lavandula Coronopifolia Poir.). Hortic. Environ. Biotechnol. 2017, 58, 393–405. [Google Scholar] [CrossRef]
- Devrnja, N.; Krstić-Milošević, D.; Janošević, D.; Tešević, V.; Vinterhalter, B.; Savić, J.; Ćalić, D. In Vitro Cultivation of Tansy (Tanacetum vulgare L.): A Tool for the Production of Potent Pharmaceutical Agents. Protoplasma 2021, 258, 587–599. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. In Vitro Mass Propagation of Cannabis sativa L.: A Protocol Refinement Using Novel Aromatic Cytokinin Meta-Topolin and the Assessment of Eco-Physiological, Biochemical and Genetic Fidelity of Micropropagated Plants. J. Appl. Res. Med. Aromat. Plants 2016, 3, 18–26. [Google Scholar] [CrossRef]
- Muraseva, D.S.; Kostikova, V.A. In Vitro Propagation of Spiraea Betulifolia Subsp. Aemiliana (Rosaceae) and Comparative Analysis of Phenolic Compounds of Microclones and Intact Plants. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 144, 493–504. [Google Scholar] [CrossRef]
- Erişen, S.; Kurt-Gür, G.; Servi, H. In Vitro Propagation of Salvia sclarea L. by Meta-Topolin, and Assessment of Genetic Stability and Secondary Metabolite Profiling of Micropropagated Plants. Ind. Crops Prod. 2020, 157, 112892. [Google Scholar] [CrossRef]
- Castilho, C.V.; Leitão, S.G.; Silva, V.D.; Miranda, C.d.O.; Santos, M.C.d.S.; Bizzo, H.R.; da Silva, N.C. In Vitro Propagation of a Carvacrol-Producing Type of Lippia Origanoides Kunth: A Promising Oregano-like Herb. Ind. Crops Prod. 2019, 130, 491–498. [Google Scholar] [CrossRef]
- Zimmerman, J.L. Somatic Embryogenesis: A Model for Early Development in Higher Plants. Plant Cell 1993, 5, 1411. [Google Scholar] [CrossRef] [Green Version]
- Razdan, M.K. Introduction To Plant Tissue Culture, 2/E; Oxford and IBH Publishing: Oxford, UK, 2002. [Google Scholar]
- Sharma, S.; Rathi, N.; Kamal, B.; Pundir, D.; Kaur, B.; Arya, S. Conservation of Biodiversity of Highly Important Medicinal Plants of India through Tissue Culture Technology-a Review. Agric. Biol. J. N. Am. 2010, 1, 827–833. [Google Scholar] [CrossRef]
- Sharma, R.K.; Khajuria, A.K. Somatic Embryogenesis and Plant Regeneration in Ferula Jaeschkeana Vatke: A Threatened Medicinal Herb. Vegetos 2020, 33, 658–664. [Google Scholar] [CrossRef]
- Soliman, H.I.; Abo-El-Hasan, F.M.; Ayman, S.; Mabrouk, Y.M. Influence of Plant Growth Regulators on Somatic Embryogenesis Induction in Seriphidium Herba-Album. Int. J. Environ. Agric. Biotechnol. 2018, 3, 264401. [Google Scholar] [CrossRef] [Green Version]
- Kulus, D.; Tymoszuk, A. Induction of Callogenesis, Organogenesis, and Embryogenesis in Non-Meristematic Explants of Bleeding Heart and Evaluation of Chemical Diversity of Key Metabolites from Callus. Int. J. Mol. Sci. 2020, 21, 5826. [Google Scholar] [CrossRef]
- Bhusare, B.P.; John, C.K.; Bhatt, V.P.; Nikam, T.D. Induction of Somatic Embryogenesis in Leaf and Root Explants of Digitalis Lanata Ehrh.: Direct and Indirect Method. S. Afr. J. Bot. 2020, 130, 356–365. [Google Scholar] [CrossRef]
- Debnath, M.; Malik, C.P.; Bisen, P.S. Micropropagation: A Tool for the Production of High Quality Plant-Based Medicines. Curr. Pharm. Biotechnol. 2006, 7, 33–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paek, K.Y.; Chakrabarty, D.; Hahn, E.J. Application of Bioreactor Systems for Large Scale Production of Horticultural and Medicinal Plants. In Liquid Culture Systems for In Vitro Plant Propagation; Springer: Berlin/Heidelberg, Germany, 2005; pp. 95–116. [Google Scholar]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In Vitro Plant Tissue Culture: Means for Production of Biological Active Compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef]
- DiCosmo, F.; Misawa, M. Plant Cell and Tissue Culture: Alternatives for Metabolite Production. Biotechnol. Adv. 1995, 13, 425–453. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Weber, J.; Maciuk, A. Bioprocessing of Plant Cell Cultures for Mass Production of Targeted Compounds. Appl. Microbiol. Biotechnol. 2009, 83, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Lee, E.-J.; Paek, K.-Y. Production of Secondary Metabolites from Cell and Organ Cultures: Strategies and Approaches for Biomass Improvement and Metabolite Accumulation. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Wu, C.-F.; Karioti, A.; Rohr, D.; Bilia, A.R.; Efferth, T. Production of Rosmarinic Acid and Salvianolic Acid B from Callus Culture of Salvia Miltiorrhiza with Cytotoxicity towards Acute Lymphoblastic Leukemia Cells. Food Chem. 2016, 201, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Akula, R.; Ravishankar, G.A. Influence of Abiotic Stress Signals on Secondary Metabolites in Plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Anusha, T.S.; Joseph, M.V.; Elyas, K.K. Callus Induction and Elicitation of Total Phenolics in Callus Cell Suspension Culture of Celastrus Paniculatus–Willd, an Endangered Medicinal Plant in India. Pharmacogn. J. 2016, 8, 471–475. [Google Scholar]
- Khan, T.; Khan, T.; Hano, C.; Abbasi, B.H. Effects of Chitosan and Salicylic Acid on the Production of Pharmacologically Attractive Secondary Metabolites in Callus Cultures of Fagonia Indica. Ind. Crops Prod. 2019, 129, 525–535. [Google Scholar] [CrossRef]
- Namdeo, A.G. Plant Cell Elicitation for Production of Secondary Metabolites: A Review. Pharm. Rev. 2007, 1, 69–79. [Google Scholar]
- Sharma, M.; Ahuja, A.; Gupta, R.; Mallubhotla, S. Enhanced Bacoside Production in Shoot Cultures of Bacopa Monnieri under the Influence of Abiotic Elicitors. Nat. Prod. Res. 2015, 29, 745–749. [Google Scholar] [CrossRef]
- Dörnenburg, H.; Knorr, D. Strategies for the Improvement of Secondary Metabolite Production in Plant Cell Cultures. Enzym. Microb. Technol. 1995, 17, 674–684. [Google Scholar] [CrossRef]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant Growth Regulator Mediated Consequences of Secondary Metabolites in Medicinal Plants. J. Appl. Res. Med. Aromat. Plants 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Monfort, L.E.F.; Bertolucci, S.K.V.; Lima, A.F.; de Carvalho, A.A.; Mohammed, A.; Blank, A.F.; Pinto, J.E.B.P. Effects of Plant Growth Regulators, Different Culture Media and Strength MS on Production of Volatile Fraction Composition in Shoot Cultures of Ocimum Basilicum. Ind. Crops Prod. 2018, 116, 231–239. [Google Scholar] [CrossRef]
- Azeez, H.A.; Ibrahim, K.M. Hypericum Triquetrifolium Callus Cultures a Potential Source of Phenolics and Flavonoids. JZS-Part A Spec. Issue 2014, 16, 381–388. [Google Scholar] [CrossRef]
- Karalija, E.; Paric, A. The Effect of BA and IBA on the Secondary Metabolite Production by Shoot Culture of Thymus vulgaris L. Biol. Nyssana 2011, 2, 29–35. [Google Scholar]
- Sánchez-Ramos, M.; Alvarez, L.; Romero-Estrada, A.; Bernabé-Antonio, A.; Marquina-Bahena, S.; Cruz-Sosa, F. Establishment of a Cell Suspension Culture of Ageratina Pichinchensis (Kunth) for the Improved Production of Anti-Inflammatory Compounds. Plants 2020, 9, 1398. [Google Scholar] [CrossRef] [PubMed]
- Bulchandani, N.; Shekhawat, G.S. Salicylic Acid Mediated up Regulation of Carvone Biosynthesis during Growth Phase in Cell Suspension Cultures of Anethum Graveolens. 3 Biotech 2020, 10, 482. [Google Scholar] [CrossRef] [PubMed]
- Ardianto, C.; Khotib, J.; Purwanto, D.A.; Muslihatin, W. Production of the Secondary Metabolite Catechin by in Vitro Cultures of Camellia sinensis L. J. Basic Clin. Physiol. Pharmacol. 2020, 31. [Google Scholar] [CrossRef]
- Kianersi, F.; Abdollahi, M.R.; Mirzaie-asl, A.; Dastan, D.; Rasheed, F. Biosynthesis of Rutin Changes in Capparis Spinosa Due to Altered Expression of Its Pathway Genes under Elicitors’ Supplementation. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 141, 619–631. [Google Scholar] [CrossRef]
- Ali, A.; Mohammad, S.; Khan, M.A.; Raja, N.I.; Arif, M.; Kamil, A.; Mashwani, Z.-R. Silver Nanoparticles Elicited in Vitro Callus Cultures for Accumulation of Biomass and Secondary Metabolites in Caralluma Tuberculata. Artif. Cells Nanomed. Biotechnol. 2019, 47, 715–724. [Google Scholar] [CrossRef] [Green Version]
- Roat, C.; Ramawat, K.G. Elicitor-Induced Accumulation of Stilbenes in Cell Suspension Cultures of Cayratia trifolia (L.) Domin. Plant Biotechnol. Rep. 2009, 3, 135–138. [Google Scholar] [CrossRef]
- Abd Alhady, M.R.A.A.; Abo El-Fadl, R.E.-S.; Hegazi, G.A.E.-M.; Desoukey, S.Y. In Vitro Production of Some Secondary Metabolites from Cupressus Sempervirens. J. Adv. Biomed. Pharm. Sci. 2020, 3, 127–134. [Google Scholar] [CrossRef]
- Bernabé-Antonio, A.; Sánchez-Sánchez, A.; Romero-Estrada, A.; Meza-Contreras, J.C.; Silva-Guzmán, J.A.; Fuentes-Talavera, F.J.; Hurtado-Díaz, I.; Alvarez, L.; Cruz-Sosa, F. Establishment of a Cell Suspension Culture of Eysenhardtia Platycarpa: Phytochemical Screening of Extracts and Evaluation of Antifungal Activity. Plants 2021, 10, 414. [Google Scholar] [CrossRef] [PubMed]
- El-Ashry, A.A.E.-L.; Gabr, A.M.M.; Arafa, N.M.; El-Bahr, M.K. Rutin Accumulation in Gardenia Calli Cultures as a Response to Phenyl Alanine and Salicylic Acid. Bull. Natl. Res. Cent. 2019, 43, 141. [Google Scholar] [CrossRef] [Green Version]
- Chodisetti, B.; Rao, K.; Gandi, S.; Giri, A. Gymnemic Acid Enhancement in the Suspension Cultures of Gymnema Sylvestre by Using the Signaling Molecules—Methyl Jasmonate and Salicylic Acid. Vitr. Cell. Dev. Biol.-Plant 2015, 51, 88–92. [Google Scholar] [CrossRef]
- Duangporn, P.; Siripong, P. Effect of Auxin and Cytokinin on Phyllanthusol A Production by Callus Cultures of Phyllanthus Acidus Skeels. Am.-Eurasian J Agric Env. Sci 2009, 5, 258–263. [Google Scholar]
- Arya, D.; Patni, V.; Kant, U. In Vitro Propagation and Quercetin Quantification in Callus Cultures of Rasna (Pluchea Lanceolata Oliver & Hiern.). Indian J. Biotechnol. 2008, 7, 383–387. [Google Scholar]
- Pérez-Mendoza, M.B.; Llorens-Escobar, L.; Vanegas-Espinoza, P.E.; Cifuentes, A.; Ibáñez, E.; Villar-Martínez, A.A.D. Chemical Characterization of Leaves and Calli Extracts of Rosmarinus Officinalis by UHPLC-MS. Electrophoresis 2020, 41, 1776–1783. [Google Scholar] [CrossRef]
- Açıkgöz, M.A. Effects of Sorbitol on the Production of Phenolic Compounds and Terpenoids in the Cell Suspension Cultures of Ocimum basilicum L. Biologia 2021, 76, 395–409. [Google Scholar] [CrossRef]
- Najhah, M.Y.; Jaafar, H.Z.; Nakasha, J.J.; Hakiman, M. Shoot Multiplication and Callus Induction of Labisia Pumila Var. Alata as Influenced by Different Plant Growth Regulators Treatments and Its Polyphenolic Activities Compared with the Wild Plant. Molecules 2021, 26, 3229. [Google Scholar] [CrossRef]
- Gao, S.L.; Zhu, D.N.; Cai, Z.H.; Jiang, Y.; Xu, D.R. Organ Culture of a Precious Chinese Medicinal Plant–Fritillaria Unibracteata. Plant Cell Tissue Organ Cult. 1999, 59, 197–201. [Google Scholar] [CrossRef]
- Haida, Z.; Nakasha, J.J.; Hakiman, M. In Vitro Responses of Plant Growth Factors on Growth, Yield, Phenolics Content and Antioxidant Activities of Clinacanthus Nutans (Sabah Snake Grass). Plants 2020, 9, 1030. [Google Scholar] [CrossRef] [PubMed]
- Velayutham, P.; Karthi, C. GC-MS Profile of in Vivo, in Vitro and Fungal Elicited in Vitro Leaves of Hybanthus enneaspermus (L.) F. Muell. Int. J. Pharm. Pharm. Sci. 2015, 7, 260–267. [Google Scholar]
- Panda, A.K.; Mishra, S.; Bisaria, V.S.; Bhojwani, S.S. Plant Cell Reactors—A Perspective. Enzym. Microb. Technol. 1989, 11, 386–397. [Google Scholar] [CrossRef]
- Tanaka, H. Technological Problems in Cultivation of Plant Cells at High Density. Biotechnol. Bioeng. 2000, 67, 775–790. [Google Scholar] [CrossRef]
- Pilarek, M.; Sobieszuk, P.; Wierzchowski, K.; Dąbkowska, K. Impact of Operating Parameters on Values of a Volumetric Mass Transfer Coefficient in a Single-Use Bioreactor with Wave-Induced Agitation. Chem. Eng. Res. Des. 2018, 136, 1–10. [Google Scholar] [CrossRef]
- Stiles, A.R.; Liu, C.-Z. Hairy Root Culture: Bioreactor Design and Process Intensification. Biotechnol. Hairy Root Syst. 2013, 134, 91–114. [Google Scholar]
- Eibl, R.; Eibl, D. Design of Bioreactors Suitable for Plant Cell and Tissue Cultures. Phytochem. Rev. 2008, 7, 593–598. [Google Scholar] [CrossRef]
- Eibl-Schindler, R.; Löffelholz, C.; Eibl, D. Single-Use Bioreactors: An Overview. Single-Use Technol. Biopharm. Manuf. 2011, 4, 145–158. [Google Scholar]
- Mamun, N.H.; Egertsdotter, U.; Aidun, C.K. Bioreactor Technology for Clonal Propagation of Plants and Metabolite Production. Front. Biol. 2015, 10, 177–193. [Google Scholar] [CrossRef]
- Mehring, A.; Haffelder, J.; Chodorski, J.; Stiefelmaier, J.; Strieth, D.; Ulber, R. Establishment and Triterpenoid Production of Ocimum Basilicum Cambial Meristematic Cells. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 143, 573–581. [Google Scholar] [CrossRef]
- Valdiani, A.; Hansen, O.K.; Nielsen, U.B.; Johannsen, V.K.; Shariat, M.; Georgiev, M.I.; Omidvar, V.; Ebrahimi, M.; Tavakoli Dinanai, E.; Abiri, R. Bioreactor-Based Advances in Plant Tissue and Cell Culture: Challenges and Prospects. Crit. Rev. Biotechnol. 2019, 39, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Kreis, W.; Baron, D.; Stoll, G. Biotechnologie Der Arzneistoffe: Grundlagen Und Anwendungen; Deutscher Apotheker Verlag: Stuttgart, Germany, 2001. [Google Scholar]
- Hibino, K.; Ushiyama, K. Commercial Production of Ginseng by Plant Tissue Culture Technology. In Plant Cell and Tissue Culture for the Production of Food Ingredients; Springer: Berlin/Heidelberg, Germany, 1999; pp. 215–224. [Google Scholar]
- Wink, M.; Alfermann, A.W.; Franke, R.; Wetterauer, B.; Distl, M.; Windhövel, J.; Krohn, O.; Fuss, E.; Garden, H.; Mohagheghzadeh, A. Sustainable Bioproduction of Phytochemicals by Plant in Vitro Cultures: Anticancer Agents. Plant Genet. Resour. 2005, 3, 90–100. [Google Scholar] [CrossRef]
- Ritala, A.; Heiniö, R.; Häkkinen, S.T.; Lille, M.; Hyytiäinen-Pabst, T.; Rischer, H. Tailoring Sensory Properties of Plant Cell Cultures for Food Use. Food Res. Int. 2022, 157, 111440. [Google Scholar] [CrossRef]
- Jeong, C.S.; Murthy, H.N.; Hahn, E.J.; Lee, H.L.; Paek, K.Y. Inoculum Size and Auxin Concentration Influence the Growth of Adventitious Roots and Accumulation of Ginsenosides in Suspension Cultures of Ginseng (Panax Ginseng CA Meyer). Acta Physiol. Plant. 2009, 31, 219–222. [Google Scholar] [CrossRef]
- Kokotkiewicz, A.; Zabiegala, B.; Kubica, P.; Szopa, A.; Bucinski, A.; Ekiert, H.; Luczkiewicz, M. Accumulation of Volatile Constituents in Agar and Bioreactor Shoot Cultures of Verbena officinalis L. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 144, 671–679. [Google Scholar] [CrossRef]
- Saadah, I.N.; Kristanti, A.N.; Hardjo, P.H.; Manuhara, Y.S.W. Shoots Culture of Gynura Procumbens (Lour.) Merr. in Balloon-Type Bubble-Bioreactor Influenced by Sucrose Concentration and Inoculums Density. Asian J. Plant Sci. 2019, 18, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Savitha, B.C.; Thimmaraju, R.; Bhagyalakshmi, N.; Ravishankar, G.A. Different Biotic and Abiotic Elicitors Influence Betalain Production in Hairy Root Cultures of Beta Vulgaris in Shake-Flask and Bioreactor. Process Biochem. 2006, 41, 50–60. [Google Scholar] [CrossRef]
- Suresh, B.; Thimmaraju, R.; Bhagyalakshmi, N.; Ravishankar, G.A. Polyamine and Methyl Jasmonate-Influenced Enhancement of Betalaine Production in Hairy Root Cultures of Beta Vulgaris Grown in a Bubble Column Reactor and Studies on Efflux of Pigments. Process Biochem. 2004, 39, 2091–2096. [Google Scholar] [CrossRef]
- Shohael, A.M.; Chakrabarty, D.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Application of Bioreactor System for Large-Scale Production of Eleutherococcus Sessiliflorus Somatic Embryos in an Air-Lift Bioreactor and Production of Eleutherosides. J. Biotechnol. 2005, 120, 228–236. [Google Scholar] [CrossRef]
- Ali, M.B.; Hahn, E.-J.; Paek, K.-Y. Methyl Jasmonate and Salicylic Acid Induced Oxidative Stress and Accumulation of Phenolics in Panax Ginseng Bioreactor Root Suspension Cultures. Molecules 2007, 12, 607–621. [Google Scholar] [CrossRef]
- Cui, X.-H.; Chakrabarty, D.; Lee, E.-J.; Paek, K.-Y. Production of Adventitious Roots and Secondary Metabolites by Hypericum perforatum L. in a Bioreactor. Bioresour. Technol. 2010, 101, 4708–4716. [Google Scholar] [CrossRef] [PubMed]
- Pramita, A.D.; Kristanti, A.N.; Utami, E.S.W.; Manuhara, Y.S.W. Production of Biomass and Flavonoid of Gynura Procumbens (Lour.) Merr Shoots Culture in Temporary Immersion System. J. Genet. Eng. Biotechnol. 2018, 16, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamkhi, I.; Benali, T.; Aanniz, T.; Elmenyiy, N.; Guaouguaou, F.-E.; El Omari, N.; El-Shazly, M.; Zengin, G.; Bouyahya, A. Plant-Microbial Interaction: The Mechanism and the Application of Microbial Elicitor Induced Secondary Metabolites Biosynthesis in Medicinal Plants. Plant Physiol. Biochem. 2021, 167, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, J.; Bernstein, N. Elicitation: An Underutilized Tool in the Development of Medicinal Plants as a Source of Therapeutic Secondary Metabolites. Adv. Agron. 2014, 124, 201–230. [Google Scholar]
- Malook, S.U.; Maqbool, S.; Hafeez, M.; Karunarathna, S.C.; Suwannarach, N. Molecular and Biochemical Mechanisms of Elicitors in Pest Resistance. Life. 2022, 12, 844. [Google Scholar] [CrossRef]
- Saha, S.; Pal, D. Elicitor Signal Transduction Leading to the Production of Plant Secondary Metabolites. In Bioactive Natural Products for Pharmaceutical Applications; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–39. [Google Scholar]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving Production of Plant Secondary Metabolites through Biotic and Abiotic Elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Bhaskar, R.; Xavier, L.S.E.; Udayakumaran, G.; Kumar, D.S.; Venkatesh, R.; Nagella, P. Biotic Elicitors: A Boon for the In Vitro Production of Plant Secondary Metabolites. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 5, 1–18. [Google Scholar] [CrossRef]
- Ahmad, N.; Rab, A.; Ahmad, N. Light-Induced Biochemical Variations in Secondary Metabolite Production and Antioxidant Activity in Callus Cultures of Stevia Rebaudiana (Bert). J. Photochem. Photobiol. B Biol. 2016, 154, 51–56. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Naik, P.M. Elicitor-Induced Production of Biomass and Pharmaceutical Phenolic Compounds in Cell Suspension Culture of Date Palm (Phoenix dactylifera L.). Molecules 2020, 25, 4669. [Google Scholar] [CrossRef] [PubMed]
- Allah Shoja, A.; Ganjeali, A. Stimulation of Phenolic Compounds Accumulation and Antioxidant Activity in In Vitro Culture of Salvia Tebesana Bunge in Response to Nano-TiO2 and Methyl Jasmonate Elicitors. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 149, 423–440. [Google Scholar] [CrossRef]
- Giri, C.C.; Zaheer, M. Chemical Elicitors versus Secondary Metabolite Production in Vitro Using Plant Cell, Tissue and Organ Cultures: Recent Trends and a Sky Eye View Appraisal. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 126, 1–18. [Google Scholar] [CrossRef]
- Kamarul Zaman, M.A.; Azzeme, A.M.; Ramle, I.K.; Normanshah, N.; Ramli, S.N.; Shaharuddin, N.A.; Ahmad, S.; Abdullah, S.N.A. Induction, Multiplication, and Evaluation of Antioxidant Activity of Polyalthia Bullata Callus, a Woody Medicinal Plant. Plants 2020, 9, 1772. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Raghuvanshi, R.; Bhardwaj, P.; Sood, H.; Saxena, S.; Chaurasia, O.P. Influence of Light Quality on Growth, Secondary Metabolites Production and Antioxidant Activity in Callus Culture of Rhodiola Imbricata Edgew. J. Photochem. Photobiol. B: Biol. 2018, 183, 258–265. [Google Scholar] [CrossRef]
- Lamaoui, M.; Chakhchar, A.; Benlaouane, R.; El Kharrassi, Y.; Farissi, M.; Wahbi, S.; El Modafar, C. Uprising the Antioxidant Power of Argania spinosa L. Callus through Abiotic Elicitation. Comptes Rendus Biol. 2019, 342, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Naik, P.M.; Al-Khayri, J.M. Abiotic and Biotic Elicitors–Role in Secondary Metabolites Production through in Vitro Culture of Medicinal Plants. In Abiotic Biot. Stress Plants Recent Adv. Future Perspectives; IntechOpen: London, UK, 2016; pp. 247–277. [Google Scholar]
- Chavan, J.J.; Kshirsagar, P.R.; Jadhav, S.G.; Nalavade, V.M.; Gurme, S.T.; Pai, S.R. Elicitor-Mediated Enhancement of Biomass, Polyphenols, Mangiferin Production and Antioxidant Activities in Callus Cultures of Salacia chinensis L. 3 Biotech 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, G.; Iqbal, Z.; Kumar, D.; Verma, S.K.; Rout, P.K.; ur Rahman, L. Enhanced Gymnemic Acids Production in Cell Suspension Cultures of Gymnema Sylvestre (Retz.) R. Br. Ex Sm. through Elicitation. Ind. Crops Prod. 2021, 162, 113234. [Google Scholar] [CrossRef]
- Razavizadeh, R.; Adabavazeh, F.; Komatsu, S. Chitosan Effects on the Elevation of Essential Oils and Antioxidant Activity of Carum copticum L. Seedlings and Callus Cultures under in Vitro Salt Stress. J. Plant Biochem. Biotechnol. 2020, 29, 473–483. [Google Scholar] [CrossRef]
- Khan, A.U.; Khan, T.; Khan, M.A.; Nadhman, A.; Aasim, M.; Khan, N.Z.; Ali, W.; Nazir, N.; Zahoor, M. Iron-Doped Zinc Oxide Nanoparticles-Triggered Elicitation of Important Phenolic Compounds in Cell Cultures of Fagonia Indica. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 147, 287–296. [Google Scholar] [CrossRef]
- Du, L.; Li, D.; Zhang, J.; Du, J.; Luo, Q.; Xiong, J. Elicitation of Lonicera Japonica Thunb Suspension Cell for Enhancement of Secondary Metabolites and Antioxidant Activity. Ind. Crops Prod. 2020, 156, 112877. [Google Scholar] [CrossRef]
- Açıkgöz, M.A. Establishment of Cell Suspension Cultures of Ocimum basilicum L. and Enhanced Production of Pharmaceutical Active Ingredients. Ind. Crops Prod. 2020, 148, 112278. [Google Scholar] [CrossRef]
- Bakhtiari, M.A.; Golkar, P. The Effects of Callus Elicitation on Lepidine, Phenolic Content, and Antioxidant Activity of Lepidium Sativum L.: Chitosan and Gibberellic Acid. J. Plant Growth Regul. 2021, 3, 1–13. [Google Scholar] [CrossRef]
- Shah, M.; Jan, H.; Drouet, S.; Tungmunnithum, D.; Shirazi, J.H.; Hano, C.; Abbasi, B.H. Chitosan Elicitation Impacts Flavonolignan Biosynthesis in Silybum marianum (L.) Gaertn Cell Suspension and Enhances Antioxidant and Anti-Inflammatory Activities of Cell Extracts. Molecules 2021, 26, 791. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, S.; Moieni, A.; Safaie, N.; Sabet, M.S.; Salehi, M. Fungal Cell Wall and Methyl-β–Cyclodextrin Synergistically Enhance Paclitaxel Biosynthesis and Secretion in Corylus Avellana Cell Suspension Culture. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Sherif, F.; Albotnoor, N.; Yap, Y.-K.; Meligy, A.; Khattab, S. Enhanced Bioactive Compounds Composition in Lavandula Officinalis In-Vitro Plantlets Using NaCl and Moringa Oleifera, Aloe Vera and Spirulina Platensis Extracts. Ind. Crops Prod. 2020, 157, 112890. [Google Scholar] [CrossRef]
- Salehi, B.; Ata, S.; Kumar, N.V.A.; Sharopov, F.; Ramírez-Alarcón, K.; Ruiz-Ortega, A.; Ayatollahi, S.A.; Fokou, P.V.T.; Kobarfard, F.; Zakaria, A.; et al. Antidiabetic Potential of Medicinal Plants and Their Active Components. Biomolecules 2019, 9, 551. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liang, W.; Yao, L.; Paek, K.-Y.; Wang, J.; Gao, W. Production of Ginsenoside by Chaetomium Sp. and Its Effect on Enhancing the Contents of Ginsenosides in Panax Ginseng Adventitious Roots. Biochem. Eng. J. 2021, 174, 108100. [Google Scholar] [CrossRef]
- Hashemi, S.M.; Naghavi, M.R. Production and Gene Expression of Morphinan Alkaloids in Hairy Root Culture of Papaver orientale L. Using Abiotic Elicitors. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 125, 31–41. [Google Scholar] [CrossRef]
- Malarz, J.; Michalska, K.; Yudina, Y.V.; Stojakowska, A. Hairy Root Cultures as a Source of Polyphenolic Antioxidants: Flavonoids, Stilbenoids and Hydrolyzable Tannins. Plants 2022, 11, 1950. [Google Scholar] [CrossRef]
- Markoski, M.M.; Garavaglia, J.; Oliveira, A.; Olivaes, J.; Marcadenti, A. Molecular Properties of Red Wine Compounds and Cardiometabolic Benefits. Nutr. Metab. Insights 2016, 9, NMI-S32909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, H.; Su, L.; Wang, H.; Zhang, Z. Chitosan Elicitation of Saponin Accumulation in Psammosilene Tunicoides Hairy Roots by Modulating Antioxidant Activity, Nitric Oxide Production and Differential Gene Expression. Plant Physiol. Biochem. 2021, 166, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.R.; Gajurel, G.; Ahmed, I.; Roedel, K.; Medina-Bolivar, F. Induction of the Prenylated Stilbenoids Arachidin-1 and Arachidin-3 and Their Semi-Preparative Separation and Purification from Hairy Root Cultures of Peanut (Arachis hypogaea L.). Molecules 2022, 27, 6118. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.M.; Naghavi, M.R.; Ghorbani, M.; Priyanatha, C.; Zandi, P. Effects of Abiotic Elicitors on Expression and Accumulation of Three Candidate Benzophenanthridine Alkaloids in Cultured Greater Celandine Cells. Molecules 2021, 26, 1395. [Google Scholar] [CrossRef] [PubMed]
- Nazir, S.; Jan, H.; Zaman, G.; Ahmed, N.; Drouet, S.; Hano, C.; Abbasi, B.H. Synergistic Effects of Salicylic Acid and Light Stress on Bioactive Metabolites in Basil Callus Cultures. Biocatal. Agric. Biotechnol. 2021, 37, 102176. [Google Scholar] [CrossRef]
- Singh, N.; Kumaria, S. Deciphering the Role of Stress Elicitors on the Differential Modulation of Chalcone Synthase Gene and Subsequent Production of Secondary Metabolites in Micropropagated Coelogyne Ovalis Lindl., a Therapeutically Important Medicinal Orchid. S. Afr. J. Bot. 2021, 140, 336–348. [Google Scholar] [CrossRef]
- Taherkhani, T.; Asghari Zakaria, R.; Omidi, M.; Zare, N. Effect of Ultrasonic Waves on Crocin and Safranal Content and Expression of Their Controlling Genes in Suspension Culture of Saffron (Crocus sativus L.). Nat. Prod. Res. 2019, 33, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Elicitation of Silver Nanoparticles Enhanced the Secondary Metabolites and Pharmacological Activities in Cell Suspension Cultures of Bitter Gourd. 3 Biotech 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Chen, J.; Li, L.; Tian, P.; Xiang, W.; Lu, X.; Huang, R.; Li, L. Fungal Endophytes from Medicinal Plant Bletilla Striata (Thunb.) Reichb. F. Promote the Host Plant Growth and Phenolic Accumulation. S. Afr. J. Bot. 2021, 143, 25–32. [Google Scholar] [CrossRef]
- Li, J.; Liu, S.; Wang, J.; Li, J.; Liu, D.; Li, J.; Gao, W. Fungal Elicitors Enhance Ginsenosides Biosynthesis, Expression of Functional Genes as Well as Signal Molecules Accumulation in Adventitious Roots of Panax Ginseng CA Mey. J. Biotechnol. 2016, 239, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.-J.; An, X.-L.; Sun, H.-D.; Piao, X.-C.; Gao, R.; Lian, M.-L. Ginsenoside Synthesis of Adventitious Roots in Panax Ginseng Is Promoted by Fungal Suspension Homogenate of Alternaria Panax and Regulated by Several Signaling Molecules. Ind. Crops Prod. 2020, 150, 112414. [Google Scholar] [CrossRef]
- Lertphadungkit, P.; Suksiriworapong, J.; Satitpatipan, V.; Sirikantaramas, S.; Wongrakpanich, A.; Bunsupa, S. Enhanced Production of Bryonolic Acid in Trichosanthes cucumerina L.(Thai Cultivar) Cell Cultures by Elicitors and Their Biological Activities. Plants 2020, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, M.; Nekonan, M.S.; Setorki, M. Enhancement Production of Phenolic Compounds in The Cell Suspension Culture of Iberis amara L.: The Effect of Chitosan Elicitation. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Singh, T.; Sharma, U.; Agrawal, V. Isolation and Optimization of Plumbagin Production in Root Callus of Plumbago zeylanica L. Augmented with Chitosan and Yeast Extract. Ind. Crops Prod. 2020, 151, 112446. [Google Scholar] [CrossRef]
- Brzycki, C.M.; Young, E.M.; Roberts, S.C. Secondary Metabolite Production in Plant Cell Culture: A New Epigenetic Frontier. Explor. Plant Cells Prod. Compd. Interest 2021, 1, 1–37. [Google Scholar]

| Plant | Explant Source | Shoot Multiplication | Rooting | Phytochemical Analysis | Key Findings | References | ||
|---|---|---|---|---|---|---|---|---|
| MS Medium | Phytohormone | MS Medium | Phytohormone | |||||
| Zingiber officinale Roscoe | Rhizome sproutedbud | solid | Zeatin (10 µM) | solid | NAA (7.5 µM) | Flavonoids and phenolic acids | The total content of phytochemical components is not very different from those of conventionally propagated plants. | [74] |
| Plectranthusamboinicus | Axillarybuds | semi-solid | BAP (0.4 mg/L) | semi-solid | Without PGR | Carvacrol γ-Terpinene | Essential oil yield was improved with a higher quantity of chemical compounds in vitro cultures. The in vitro regeneration was chemically true to the parent plant type. | [64] |
| Lavandula coronopifolia | Shoot tips | solid | BA (0.5 mg/L) | solid | IBA (10 mg/L) | Caffeic acid androsmarinic acid | Micropropagation was regenerated from plants with genetic fidelity to the parent plant. A remarkable difference in the chemical profiles of the in vitro culture and the wild-type plants. | [75] |
| Tanacetum vulgare | Shoot tips | solid | without PGR | liquid half-strength | Without PGR | Monoterpenes Sesquiterpene Chlorogenic acid 3,5-O-Dicaffeoylquinic acid | Spontaneously rooted seedlings at the time of propagation. Terpenes are the most abundant in essential oils. In vitro grown roots are richest in 3,5-O-dicaffeoylquinic acid. | [76] |
| Cannabis sativa | Nodal segments | solid | mT (2 µM) | solid | mT (2 µM) | Cannabinoids | Rooting was performed on the same propagation medium. Auxin was not necessary for root induction. cannabinoid level in the micropropagated plants is comparable to the mother plant. In vitro propagated plants are identical to the mother plant. | [77] |
| Eryngiumalpinum | Shoots | solid | BAP, IAA, and GA3 (each 1.0 mg/L) | __ | __ | Phenolic acids and flavonoids | The solid MS medium with BAP, IAA, and GA3 (each 1.0 mg/L) is the optimal system for micropropagation and accumulation of phenolic acids and flavonoids. An important variability in phytochemicals between the intact plant and different in vitro culture. | [6] |
| Spiraeabetulifoliasubsp. aemiliana | Axillarybuds | solid | S1 = BAP (1.0 μM) S2 = (BAP 5.0 μM) + (NAA 1.0 μM) | half-strength | S1 = S2= IBA (0.1 µM) | Phenolic acids and flavonoids | Many differences in chemical profile between in vitro culture and intact plants. Interpopulation genotypic differences in the activity of morphogenic processes have been identified in S. betulifolia in vitro culture. | [78] |
| Salvia sclarea | Nodal segments | solid | mT (2.0 mg/L) + IAA (0.2 mg/L) | solid | NAA (1.0 mg/L) | A multitude of secondary metabolites | High genetic stability of micropropagated plants. N-alkanes, tetradecanal, octadecanal, and hentriacontane are the major components from micropropagated plants. PGRs have caused variability in the content of secondary metabolite. | [79] |
| Lippiaoriganoides | Nodal segments | solid | KIN (4.6 μM) | solid | KIN (2.3 μM) | Myrcene, p-cymene, γ-terpinene, linalool, thymol, carvacrol and (E)-caryophyllene. | The presence of PGR changed the chemical profile of the volatile organic compound. | [80] |
| Family | Plant | Explant Source | Phytohormone (mg/L) for Induction SE | Basal Medium | Somatic Embryogenesis | References | |
|---|---|---|---|---|---|---|---|
| Direct | Indirect | ||||||
| Apiaceae | Ferulajaeschkeana | Petiole | 2,4-D (4.0) | MS | - | X | [84] |
| Asteraceae | Seriphidiumherba-album | Leaves | 2,4-D (1.5) + BA (0.5) | MS | - | X | [85] |
| Fumariaceae | Lamprocapnosspectabilis | Leaves | 2,4-D (0.5) + BA (0.5) | ½ MS | - | X | [86] |
| Petioles | PIC (1.0) + BA (0.5) | ||||||
| Plantaginaceae | Digitalislanata | Leaves | 2,4-D (1.0) + Kin (1.0) | MS | - | X | [87] |
| IBA (2.0) + Kin (2.0) | X | - | |||||
| Root | IBA (2.0) + Kin (2.0) | X | - | ||||
| Plant Species | Active Ingredient | Culture Condition (MS Medium) | Culture Type | References |
|---|---|---|---|---|
| Ageratinapichinchensis | Artemesinol | NAA + KIN | Suspension | [105] |
| Anethum graveolens | Carvone | BA + NAA + SA | Suspension | [106] |
| Camellia sinensis | Catechin | BAP + 2,4-D + Ph (phenylalanine) | Callus | [107] |
| Capparis spinosa | Rutin | B5 medium + 2,4-D + BAP + MeJA + SA | Callus | [108] |
| Carallumatuberculata | Total phenolics | MS + 2,4-D + BAP + AgNPs(silver nanoparticles) | Callus | [109] |
| Total flavonoid | ||||
| Cayratia trifoliata | Stilbenes | NAA + KN + MeJA | Suspension | [110] |
| Cupressus sempervirens | RutinQuercitrin | BA + NAA + GA3 | Callus | [111] |
| Eysenhardtiaplatycarpa | Total phenolics | NAA + KIN | Suspension | [112] |
| Gardeniajasminoides | Rutin | TDZ | Callus | [113] |
| Gymnemasylvestre | Gymnemic acid | 2,4-D + BA + MeJA | Suspension | [114] |
| Phyllanthus acidus | Phyllanthusol | NAA + BA | Callus | [115] |
| Pluchealanceolata | Quercetin | NAA + BAP | Callus | [116] |
| Rosmarinus officinalis | Flavonoid | 2,4-D + BAP | Callus | [117] |
| Terpenoids | ||||
| Ocimumbasilicum | Rosmarinic acid | KIN + NAA + Sorbitol | Suspension | [118] |
| Chicoric acid | ||||
| Rutin | ||||
| Linalool | ||||
| Methyl chavicol | ||||
| Labisia pumila | Total phenolics | 2,4-D + Zea | Callus | [119] |
| Total flavonoid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohaddab, M.; El Goumi, Y.; Gallo, M.; Montesano, D.; Zengin, G.; Bouyahya, A.; Fakiri, M. Biotechnology and In Vitro Culture as an Alternative System for Secondary Metabolite Production. Molecules 2022, 27, 8093. https://doi.org/10.3390/molecules27228093
Mohaddab M, El Goumi Y, Gallo M, Montesano D, Zengin G, Bouyahya A, Fakiri M. Biotechnology and In Vitro Culture as an Alternative System for Secondary Metabolite Production. Molecules. 2022; 27(22):8093. https://doi.org/10.3390/molecules27228093
Chicago/Turabian StyleMohaddab, Marouane, Younes El Goumi, Monica Gallo, Domenico Montesano, Gokhan Zengin, Abdelhakim Bouyahya, and Malika Fakiri. 2022. "Biotechnology and In Vitro Culture as an Alternative System for Secondary Metabolite Production" Molecules 27, no. 22: 8093. https://doi.org/10.3390/molecules27228093