
An Untargeted Metabolomics Approach on Carfilzomib-Induced Nephrotoxicity
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
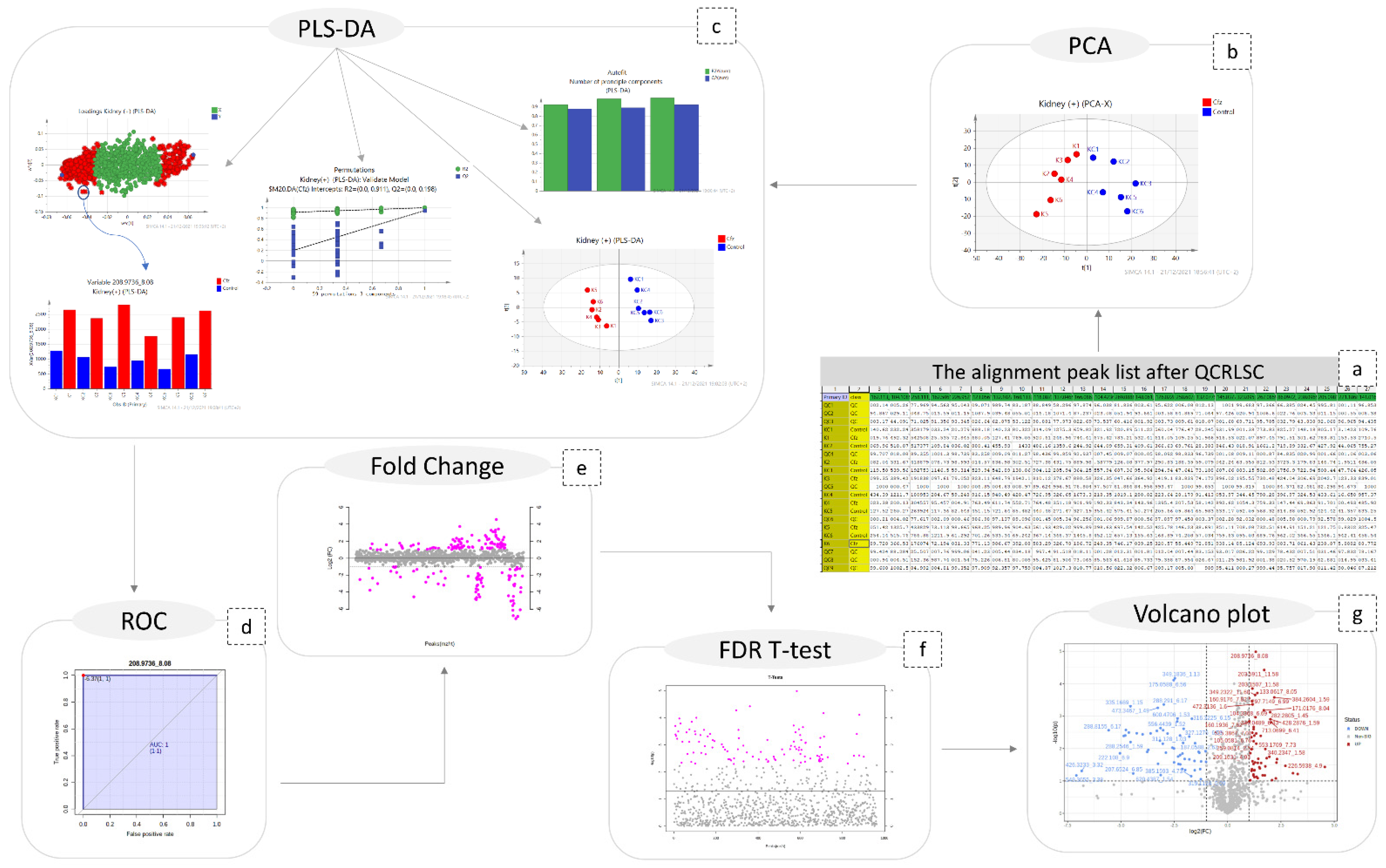
2.1. Data Pre-Processing
2.2. Statistical Analysis
2.3. Peaks Identification
3. Discussion
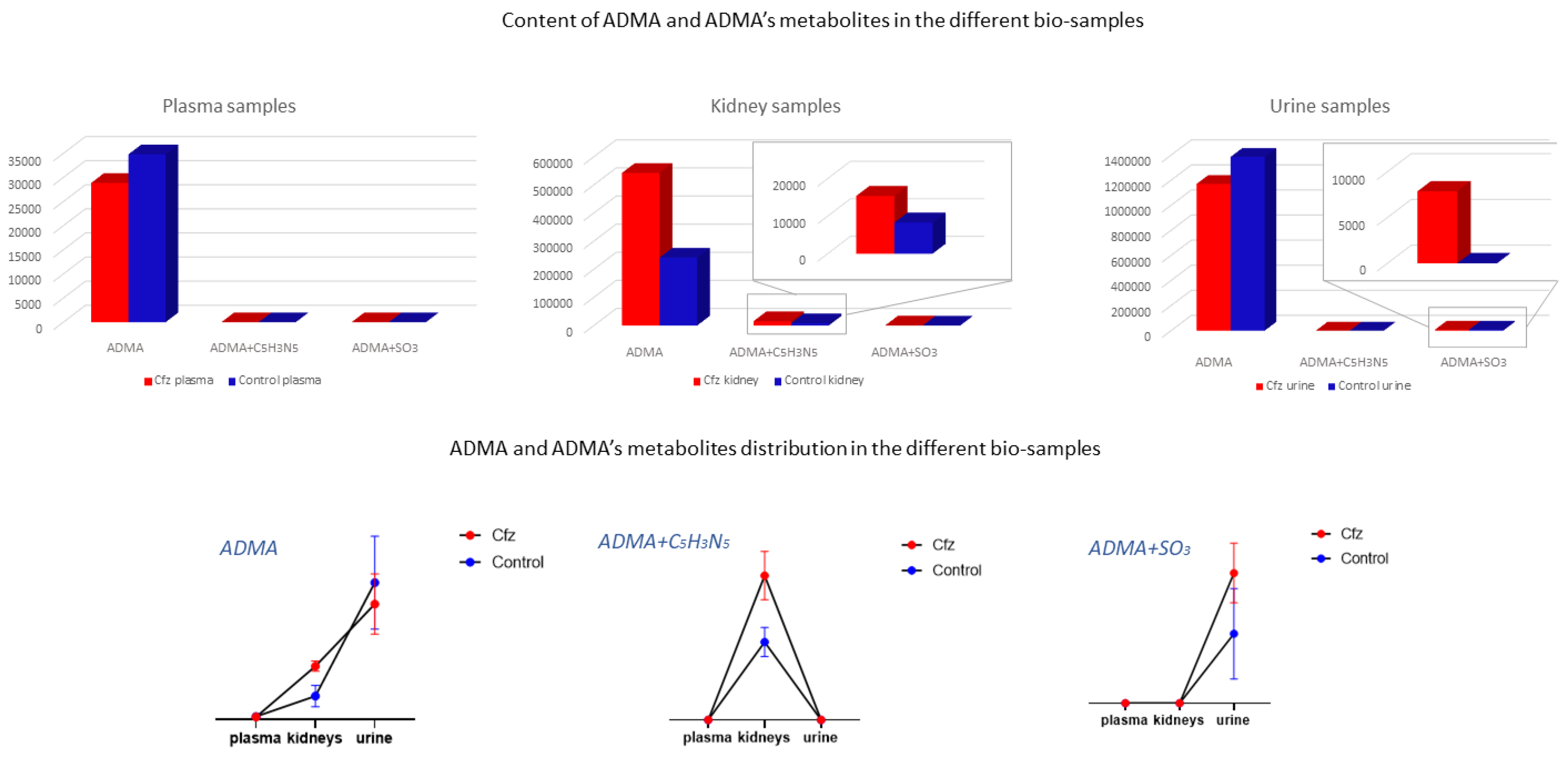
3.1. Asymetric Dimethylarginine
3.2. N1-Methyl-2-pyridone-5-carboxamide
3.3. N4-Acetylcytidine
3.4. Phenylacetic Acid
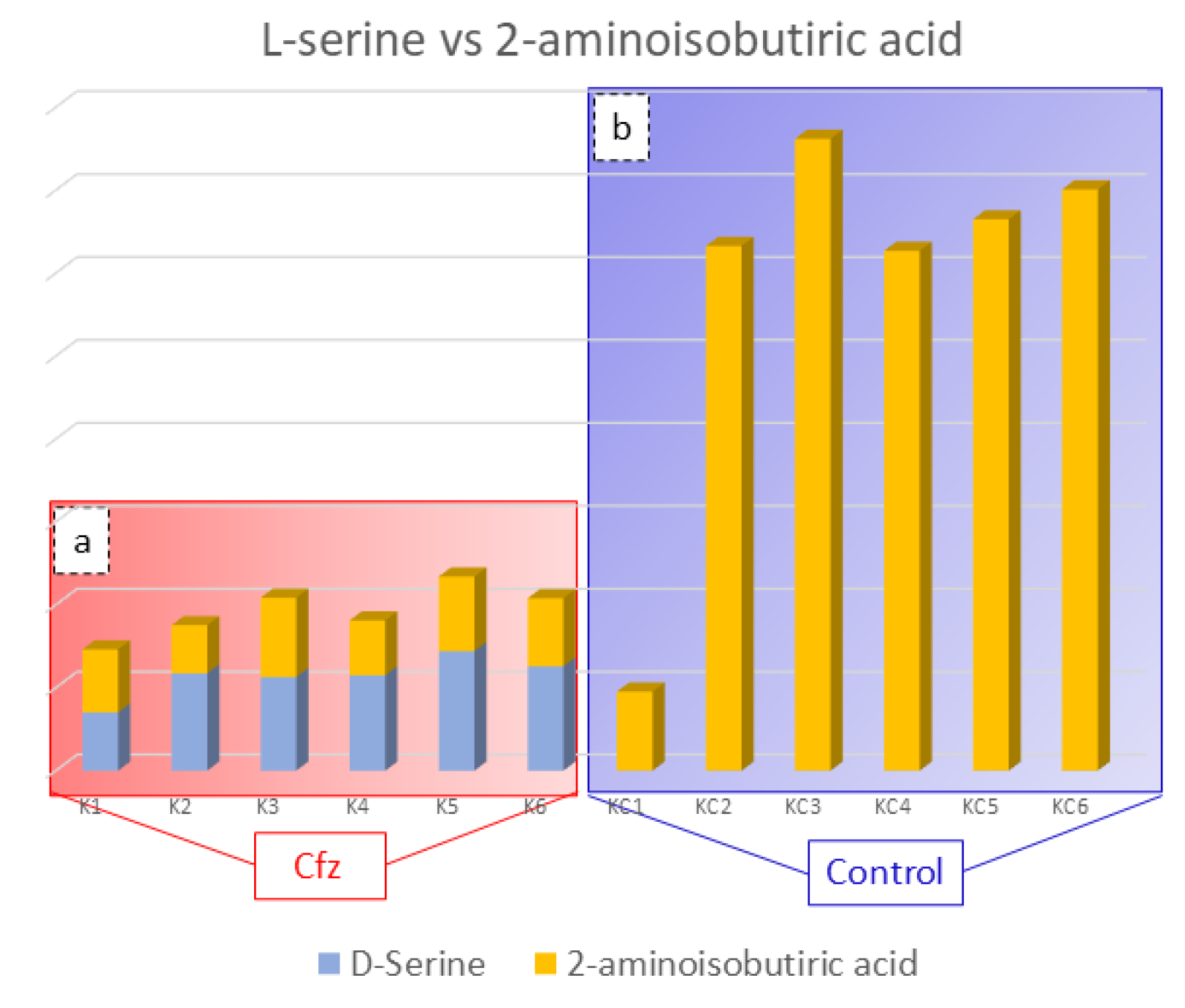
3.5. 2-Aminoisobutyric Acid
3.6. Exploration of Metabolites Alterations between Different Bio-Samples
3.7. Discovery of New Potential Biomarkers of Cfz-Related Nephrotoxicity
4. Materials and Methods
4.1. Sample Collection and Storage
4.2. Reagents and Solutions
4.3. Sample Preparation
4.4. UPLC-ESI-QTOFMS Analysis
4.5. Data Acquisition
4.6. Data Pre-Processing
4.7. Multivariate Analysis
4.8. Univariate Analysis
4.9. Peaks Identification Procedure
4.10. Data-Driven Suspect Screening of Metabolites
4.11. Exploration of Metabolites Alterations between Different Bio-Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Efentakis, P.; Kremastiotis, G.; Varela, A.; Nikolaou, P.-E.; Papanagnou, E.-D.; Davos, C.H.; Tsoumani, M.; Agrogiannis, G.; Konstantinidou, A.; Kastritis, E.; et al. Molecular mechanisms of carfilzomib-induced cardiotoxicity in mice and the emerging cardioprotective role of metformin. J. Am. Soc. Hematol. 2019, 133, 710–723. Available online: http://ashpublications.org/blood/article-pdf/133/7/710/1552545/blood858415.pdf (accessed on 17 May 2022). [CrossRef] [PubMed] [Green Version]
- Efentakis, P.; Psarakou, G.; Varela, A.; Papanagnou, E.D.; Chatzistefanou, M.; Nikolaou, P.E.; Davos, C.H.; Gavriatopoulou, M.; Trougakos, I.P.; Dimopoulos, M.A.; et al. Elucidating carfilzomib’s induced cardiotoxicity in an in vivo model of aging: Prophylactic potential of metformin. Int. J. Mol. Sci. 2021, 22, 10956. [Google Scholar] [CrossRef] [PubMed]
- Imam, F.; Al-Harbi, N.O.; Al-Harbia, M.M.; Korashy, H.M.; Ansari, M.A.; Sayed-Ahmed, M.M.; Nagi, M.N.; Iqbal, M.; Khalid Anwer, M.; Kazmi, I.; et al. Rutin Attenuates Carfilzomib-Induced Cardiotoxicity Through Inhibition of NF-κB, Hypertrophic Gene Expression and Oxidative Stress. Cardiovasc. Toxicol. 2017, 17, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Efentakis, P.; Doerschmann, H.; Witzler, C.; Siemer, S.; Nikolaou, P.E.; Kastritis, E.; Stauber, R.; Dimopoulos, M.A.; Wenzel, P.; Andreadou, I.; et al. Investigating the vascular toxicity outcomes of the irreversible proteasome inhibitor carfilzomib. Int. J. Mol. Sci. 2020, 21, 5185. [Google Scholar] [CrossRef]
- Fotiou, D.; Roussou, M.; Gakiopoulou, C.; Psimenou, E.; Gavriatopoulou, M.; Migkou, M.; Kanellias, N.; Dialoupi, I.; Eleutherakis-papaiakovou, E.; Giannouli, S.; et al. Carfilzomib-associated renal toxicity is common and unpredictable: A comprehensive analysis of 114 multiple myeloma patients. Blood Cancer J. 2020, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Tolstikov, V.; James Moser, A.; Sarangarajan, R.; Narain, N.R.; Kiebish, M.A. Current status of metabolomic biomarker discovery: Impact of study design and demographic characteristics. Metabolites 2020, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Steuer, A.E.; Brockbals, L.; Kraemer, T. Metabolomic strategies in biomarker research-new approach for indirect identification of drug consumption and sample manipulation in clinical and forensic toxicology? Front. Chem. 2019, 7, 319. [Google Scholar] [CrossRef]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [Green Version]
- Tantawy, M.; Chekka, L.M.; Huang, Y.; Garrett, T.J. Lactate Dehydrogenase B and Pyruvate Oxidation Pathway Associated With Carfilzomib-Related Cardiotoxicity in Multiple Myeloma Patients: Result of a Multi-Omics Integrative Analysis. Front. Cardiovasc. Med. 2021, 8, 645122. [Google Scholar] [CrossRef]
- Efentakis, P.; Lamprou, S.; Makridakis, M.; Barla, I.; Nikolaou, P.-E.; Christodoulou, A.; Dimitriou, C.; Kostomitsopoulos, N.; Ntanasis-Stathopoulos, I.; Theochari, I.; et al. Mineralocorticoid Receptor Pathway Is a Key Mediator of Carfilzomib-induced Nephrotoxicity: Preventive Role of Eplerenone. HemaSphere 2022, 6, e791. [Google Scholar] [CrossRef]
- Jones, O.A.H. Illuminating the dark metabolome to advance the molecular characterisation of biological systems. Metabolomics 2018, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Huan, T. Comparison of Full-Scan, Data-Dependent, and Data-Independent Acquisition Modes in Liquid Chromatography-Mass Spectrometry Based Untargeted Metabolomics. Anal. Chem. 2020, 92, 8072–8080. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Xiao, J.F.; Tuli, L.; Ressom, H.W. LC-MS-based metabolomics. Mol. Biosyst. 2012, 8, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Albert, F.; Llorach, R.; Garcia-Aloy, M.; Ziyatdinov, A.; Andres-Lacueva, C.; Perera, A. Intensity drift removal in LC/MS metabolomics by common variance compensation. Bioinformatics 2014, 30, 2899–2905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Liu, Q.; Li, Q.; Zhang, S.; Qu, X.; Zhu, J.; Zhong, G.; Huang, M. Signal Drift in Liquid Chromatography Tandem Mass Spectrometry and Its Internal Standard Calibration Strategy for Quantitative Analysis. Anal. Chem. 2020, 92, 7690–7698. [Google Scholar] [CrossRef]
- Luan, H.; Ji, F.; Chen, Y.; Cai, Z. statTarget: A streamlined tool for signal drift correction and interpretations of quantitative mass spectrometry-based omics data. Anal. Chim. Acta 2018, 1036, 66–72. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-Mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Vallance, P.; Leiper, J. Cardiovascular biology of the asymmetric dimethylarginine:dimethylarginine dimethylaminohydrolase pathway. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni, F.; Ahmetaj, B.; Leiper, J. The Biology and Therapeutic Potential of the DDAH/ADMA Pathway. Curr. Pharm. Des. 2011, 16, 4089–4102. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.P. ADMA: Its role in vascular disease. Vasc. Med. 2005, 10 (Suppl. S1), 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fliser, D.; Kielstein, J.T.; Haller, H.; Bode-Böger, S.M. Asymmetric dimethylarginine: A cardiovascular risk factor in renal disease? Kidney Int. Suppl. 2003, 63, S37–S40. [Google Scholar] [CrossRef] [Green Version]
- Basile, D.P.; Anderson, M.D.; Sutton, T.A. Pathophysiology of acute kidney injury. Compr. Physiol. 2012, 2, 1303–1353. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, Y.; Ueda, S.; Yamagishi, S.I.; Obara, N.; Taguchi, K.; Ando, R.; Kaida, Y.; Iwatani, R.; Kaifu, K.; Yokoro, M.; et al. Asymmetric dimethylarginine accumulates in the kidney during ischemia/reperfusion injury. Kidney Int. 2014, 85, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Lenglet, A.; Liabeuf, S.; Bodeau, S.; Louvet, L.; Mary, A.; Boullier, A.; Lemaire-Hurtel, A.S.; Jonet, A.; Sonnet, P.; Kamel, S.; et al. N-methyl-2-pyridone-5-carboxamide (2PY)—Major metabolite of nicotinamide: An update on an old uremic Toxin. Toxins 2016, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Ray Chaudhuri, A.; Nussenzweig, A. The multifaceted roles of PARP1 in DNA repair and chromatin remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 610–621. [Google Scholar] [CrossRef]
- Sethi, G.S.; Dharwal, V.; Naura, A.S. Poly(ADP-ribose)polymerase-1 in lung inflammatory disorders: A review. Front. Immunol. 2017, 8, 1172. [Google Scholar] [CrossRef] [Green Version]
- Deen, C.P.J.; van der Veen, A.; Gomes-Neto, A.W.; Geleijnse, J.M.; Borgonjen-Van Den Berg, K.J.; Heiner-Fokkema, M.R.; Kema, I.P.; Bakker, S.J.L. Urinary excretion of N1-methyl-2-pyridone-5-carboxamide and N1-methylnicotinamide in renal transplant recipients and donors. J. Clin. Med. 2020, 9, 437. [Google Scholar] [CrossRef]
- Jin, G.; Xu, M.; Zou, M.; Duan, S. The Processing, Gene Regulation, Biological Functions, and Clinical Relevance of N4-Acetylcytidine on RNA: A Systematic Review. Mol. Ther. Nucleic Acid 2020, 20, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Scholze, A.; Jankowski, V.; Henning, L.; Haass, W.; Jankowski, J. Phenylacetic Acid and Arterial Vascular Properties in Patients with Chronic Kidney Disease Stage 5 on Hemodialysis Therapy. Nephron 2007, 107, c1–c6. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.; Raupachova, J.; Hörl, W.H. The uraemic toxin phenylacetic acid contributes to in fl ammation by priming polymorphonuclear leucocytes. Nephrol. Dial. Transplant. 2013, 28, 421–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, T.; Hesaka, A.; Isaka, Y. d-Amino acids and kidney diseases. Clin. Exp. Nephrol. 2020, 24, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Krug, A.W.; Völker, K.; Dantzler, W.H.; Silbernagl, S. Why is D-serine nephrotoxic and-aminoisobutyric acid protective? Am. J. Physiol. Ren. Physiol. 2007, 293, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Titan, S.M.; Venturini, G.; Padilha, K.; Goulart, A.C.; Lotufo, P.A.; Bensenor, I.J.; Krieger, J.E.; Thadhani, R.I.; Rhee, E.P.; Pereira, A.C. Metabolomics biomarkers and the risk of overall mortality and ESRD in CKD: Results from the PRoGREDIR cohort. PLoS ONE 2019, 14, e0213764. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Chen, W.; Li, R.; Wang, M.; Chen, C.; Zeng, R.; Deng, Y. Systematic variations associated with renal disease uncovered by parallel metabolomics of urine and serum. BMC Syst. Biol. 2012, 6, S14. [Google Scholar] [CrossRef] [Green Version]
- Kaltenbach, J.P.; Carone, F.A.; Ganote, C.E. Compounds Protective against Renal Tubular Necrosis Induced by o-Serine and D2, 3-Diaminopropionic Acid in the Rat. Exp. Mol. Pathol. 1982, 37, 225–234. [Google Scholar] [CrossRef]
- Sun, J.; Shannon, M.; Ando, Y.; Schnackenberg, L.K.; Khan, N.A.; Portilla, D.; Beger, R.D. Serum metabolomic profiles from patients with acute kidney injury: A pilot study. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 893–894, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Boudonck, K.J.; Mitchell, M.W.; Német, L.; Keresztes, L.; Nyska, A.; Shinar, D.; Rosenstock, M. Discovery of Metabolomics Biomarkers for Early Detection of Nephrotoxicity. Toxicol. Pathol. 2009, 37, 280–292. [Google Scholar] [CrossRef]
- Mutsaers, H.A.M.; Wilmer, M.J.G.; Reijnders, D.; Jansen, J.; van den Broek, P.H.H.; Forkink, M.; Schepers, E.; Glorieux, G.; Vanholder, R.; van den Heuvel, L.P.; et al. Uremic toxins inhibit renal metabolic capacity through interference with glucuronidation and mitochondrial respiration. Biochim. Biophys. Acta Mol. Basis Dis. 2013, 1832, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Osswald, H.; Schnermann, J. Methylxanthines and the kidney. Handb. Exp. Pharmacol. 2011, 200, 391–412. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.; Liang, M. Renal metabolism and hypertension. Nat. Commun. 2021, 12, 963. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, M.A.; Roussou, M.; Gavriatopoulou, M.; Psimenou, E.; Ziogas, D.; Eleutherakis-Papaiakovou, E.; Fotiou, D.; Migkou, M.; Kanellias, N.; Panagiotidis, I.; et al. Cardiac and renal complications of carfilzomib in patients with multiple myeloma. Blood Adv. 2017, 1, 449–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The arrive guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Breitkopf, S.B.; Yang, X.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wawrzyniak, R.; Kosnowska, A.; Macioszek, S.; Bartoszewski, R.; Markuszewski, M.J. New plasma preparation approach to enrich metabolome coverage in untargeted metabolomics: Plasma protein bound hydrophobic metabolite release with proteinase K. Sci. Rep. 2018, 8, 9541. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; Mullin, L.G.; Wilson, I.D.; Coen, M.; Rainville, P.D.; Plumb, R.S.; Gethings, L.A.; Maker, G.; Trengove, R. Development of a rapid profiling method for the analysis of polar analytes in urine using HILIC–MS and ion mobility enabled HILIC–MS. Metabolomics 2019, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, A.; Han, Y.; Wang, P.; Sun, H.; Song, G.; Dong, T.; Yuan, Y.; Yuan, X.; Zhang, M.; et al. Urine metabolomics analysis for biomarker discovery and detection of Jaundice syndrome in patients with liver disease. Mol. Cell. Proteomics 2012, 11, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Afsar, F.A.; Neumann, S.; Ben-Hur, A.; Prenni, J.E. RAMClust: A novel feature clustering method enables spectral-matching-based annotation for metabolomics data. Anal. Chem. 2014, 86, 6812–6817. [Google Scholar] [CrossRef] [PubMed]
- Huan, T.; Tang, C.; Li, R.; Shi, Y.; Lin, G.; Li, L. MyCompoundID MS/MS Search: Metabolite Identification Using a Library of Predicted Fragment-Ion-Spectra of 383,830 Possible Human Metabolites. Anal. Chem. 2015, 87, 10619–10626. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Variables: | |||
|---|---|---|---|
| Dataset | Detected | After QC-RLCS | Used for Multivariate Analysis |
| Plasma (+) | 624 | 346 | 191 |
| Plasma (−) | 156 | 67 | 49 |
| Kidney (+) | 1079 | 964 | 964 |
| Kidney (−) | 239 | 195 | 195 |
| Urine (+) | 1769 | 1509 | 684 |
| Urine (−) | 533 | 458 | 328 |
| Number of Biomarkers | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PLS-DA Model | PLS-DA Permutations Test | PLS-DA Analysis | ROC Analysis | FDR-t-Test | Fold-Change | |||||
| PCs | Q2 | R2 | Higher VIP Value | (Q2) | (R2) | VIP > 1.5 | AUC > 0.9 | p-Value < 0.05 | log2(FC) > 2 | |
| Plasma (+) | 3 | 0.5 | 0.99 | 2.3 | 0.32 | 0.99 | 11 | 8 | 20 | 107 |
| Plasma (−) | 4 | 0.14 | 0.92 | 2.53 | 0.27 | 0.94 | 13 | 0 | 0 | 24 |
| Kidney (+) | 3 | 0.79 | 0.99 | 1.96 | 0.13 | 0.98 | 40 | 105 | 110 | 79 |
| Kidney (−) | 3 | 0.93 | 0.99 | 1.73 | 0.38 | 0.97 | 21 | 45 | 30 | 29 |
| Urine (+) | 4 | 0.86 | 1 | 2.35 | 0.58 | 0.99 | 23 | 68 | 82 | 926 |
| Urine (−) | 3 | 0.77 | 0.99 | 2.49 | 0.20 | 0.99 | 19 | 41 | 40 | 158 |
| Precursor Mass Exp. | Sample Type | ESI Polarity | RT | Cfz-Regulation ** | VIP | AUC | Formula | Compound | HMDB | MCID Initial Score | MCID Fit Score | Reactions of Metabolism * | Precursor Type | Precursor Mass Theo. | Error (mDa) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 153.0695 | urine | + | 3.14 | ↑ | 1.64 | 1 | C7H8N2O2 | N1-Methyl-2-pyridone-5-carboxamide | HMDB0004193 | 1 | 0.87 | NO REACTION | M + H | 153.0659 | −0.004 |
| 551.2665 | urine | + | 4.51 | ↑ | 1.62 | 1 | UNKNOWN | - | |||||||
| 135.0948 | urine | + | 6.1 | ↑ | 1.47 | 1 | C3H7N3O2 | Guanidoacetic acid | HMDB0001528 | 1 | 0.52 | [+NH3] | M + H | 135.0877 | −0.007 |
| 286.1103 | urine | + | 5.78 | ↑ | 1.44 | 1 | C11H15N3O6 | N4-Acetylcytidine | HMDB0005923 | 1 | 0.72 | NO REACTION | M + H | 286.1034 | 0.007 |
| 183.1168 | urine | + | 3.28 | ↑ | 1.40 | 1 | C9H13NO2 | p-Synephrine | HMDB0004826 | 0.96 | 0.59 | [+NH] | M + H | 183.1128 | −0.004 |
| 245.1014 | urine | + | 3.11 | ↑ | 1.40 | 1 | C10H16N2O3S | Biotin | HMDB0000030 | 1 | 0.74 | NO REACTION | M + H | 245.0954 | −0.006 |
| 181.1626 | urine | + | 1.35 | ↑ | 1.39 | 1 | C10H16O | Perillyl alcohol | HMDB0003634 | 0.99 | 0.68 | [+C2H4] | M + H | 181.1587 | −0.004 |
| 214.1852 | urine | + | 1.36 | ↑ | 1.38 | 1 | C13H23NO4 | 2-Hexenoylcarnitine | HMDB0013161 | 1 | 0.92 | [−CO2] | M + H | 214.1802 | −0.004 |
| 199.1739 | urine | + | 1.35 | ↑ | 1.35 | 1 | C10H20O | Decanal | HMDB0011623 | 0.86 | 0.83 | [+C2H2O] | M + H | 199.1693 | −0.005 |
| 283.1111 | urine | + | 5.13 | ↑ | 1.32 | 0.96 | C8H18N4O2 | Asymmetric dimethylarginine | HMDB0001539 | 0.96 | 0.55 | [+SO3] | M + H | 283.1037 | −0.007 |
| 112.1154 | urine | + | 6 | ↑ | 1.40 | 0.96 | UNKNOWN | - | |||||||
| 392.2368 | urine | + | 5.4 | ↑ | 1.34 | 0.9 | UNKNOWN | - | |||||||
| 144.9639 | urine | − | 1.03 | ↑ | 1.48 | 1 | C6H4Cl2O | 2,4-Dichlorophenol | HMDB0004811 | 1 | 0.79 | [−O] | M-H | 144.9606 | −0.003 |
| 305.1473 | urine | − | 6.07 | ↑ | 1.40 | 1 | C14H18N2O4 | Phenylalanyl-hydroxyproline | HMDB0011176 | 1 | 0.844 | [+C2H4] | M-H | 305.1496 | 0.002 |
| 279.0149 | urine | − | 1.14 | ↑ | 1.33 | 1 | UNKNOWN | - | |||||||
| 258.9891 | urine | − | 4.22 | ↑ | C9H8O4 | 4-Hydroxyphenylpyruvic acid | HMDB0000707 | 0.99 | 0.831 | [+HPO3] | M-H | 259.0002 | 0.011 | ||
| 215.0002 | urine | − | 1.17 | ↑ | 1.29 | 1 | C8H8O2 | Phenylacetic acid | HMDB0000209 | 1 | 0.77 | [+HPO3] | M-H | 215.0104 | 0.010 |
| 363.0135 | urine | − | 1.55 | ↑ | 1.28 | 1 | C3H6O3S | 3-Mercaptolactic acid | HMDB0002127 | 0.81 | 0.78 | [+C6H11O8P] | M-H | 363.0145 | 0.001 |
| 123.0116 | urine | − | 5.27 | ↑ | 1.23 | 0.9 | UNKNOWN | - | |||||||
| 324.9654 | urine | − | 4.75 | ↑ | 1.23 | 0.96 | UNKNOWN | - | |||||||
| 365.0294 | urine | − | 1.12 | ↑ | 1.21 | 0.9 | C10H15N2O9P | Imidazoleacetic acid-ribotide | HMDB0006032 | 0.86 | 0.75 | [+CO] | M-H | 365.0381 | 0.009 |
| 199.9947 | urine | − | 2.1 | ↓ | 1.30 | 1 | C3H8NO6P | Phosphoserine | HMDB0000272 | 1 | 0.75 | [+O] | M-H | 199.9955 | 0.001 |
| 230.9946 | urine | − | 1.37 | ↓ | 1.30 | 1 | C8H8O3 | 4-Hydroxy-3-methylbenzoic acid | HMDB0004815 | 1 | 0.9 | [+SO3] | M-H | 230.9958 | 0.001 |
| 144.0655 | urine | − | 2.27 | ↓ | 1.20 | 1 | C6H11NO2 | Pipecolic acid | HMDB0000070 | 1 | 0.76 | [+O] | M-H | 144.0655 | 0.000 |
| 337.0345 | urine | − | 1.63 | ↓ | 1.29 | 1 | C5H11O8P | D-Arabinose 5-phosphate | HMDB0011734 | 0.72 | 0.8 | [ +C5H4N2O] | M-H | 337.0431 | 0.009 |
| 208.9736 | kidney | + | 8.08 | ↑ | 1.38 | 1 | UNKNOWN | - | |||||||
| 349.2322 | kidney | + | 11.68 | ↑ | 1.35 | 1 | C15H29NO4 | Octanoylcarnitine | HMDB0000791 | 1 | 0.71 | [+CO2] | M + NH4 | 349.2333 | 0.001 |
| 336.1931 | kidney | + | 6.97 | ↑ | 1.31 | 1 | C8H18N4O2 | Asymmetric dimethylarginine | HMDB0001539 | 1 | 0.46 | [+C5H3N5] | M + H | 336.1891 | −0.004 |
| 133.0617 | kidney | + | 8.06 | ↑ | 1.31 | 1 | C3H8N2O2 | 2,3-Diaminopropionic acid | HMDB0002006 | 0.9 | 0.52 | [+CO] | M + H | 133.0608 | −0.001 |
| 245.0777 | kidney | + | 5.82 | ↑ | 1.30 | 1 | C9H12N2O6 | Uridine | HMDB0000296 | 1 | 0.4 | NO REACTION | M + H | 245.0768 | −0.001 |
| 160.5245 | kidney | + | 7.82 | ↑ | 1.29 | 1 | UNKNOWN | - | |||||||
| 384.2604 | kidney | + | 1.59 | ↑ | 1.28 | 1 | UNKNOWN | - | |||||||
| 203.1507 | kidney | + | 11.58 | ↑ | 1.28 | 1 | C8H18N4O2 | Asymmetric dimethylarginine | HMDB0001539 | 1 | 0.76 | NO REACTION | M + H | 203.1503 | 0.000 |
| 297.7149 | kidney | + | 6.99 | ↑ | 1.25 | 1 | C10H20O3 | 3-Hydroxycapric acid | HMDB0002203 | 1 | 0.58 | [+C5H4N2O] | M + H | 297.1809 | −0.534 |
| 258.4705 | kidney | + | 7.72 | ↑ | 1.24 | 1 | UNKNOWN | - | |||||||
| 166.0867 | kidney | + | 6.97 | ↑ | 1.24 | 1 | C9H10O2 | 4-Ethylbenzoic acid | HMDB0002097 | 1 | 0.63 | [+NH] | M + H | 166.0863 | 0.000 |
| 120.42 | kidney | + | 6.97 | ↑ | 1.24 | 0.97 | C8H11N | 1-Phenylethylamine | HMDB0002017 | 0.96 | 0.71 | [−H2] | M + H | 120.0808 | −0.339 |
| 331.1662 | kidney | + | 6.96 | ↑ | 1.23 | 1 | C18H21NO4 | (S)-3-Hydroxy-N-methylcoclaurine | HMDB0006921 | 1 | 0.7 | [+NH] | M + H | 331.1652 | −0.001 |
| 166.8851 | kidney | + | 6.95 | ↑ | 1.23 | 1 | UNKNOWN | - | |||||||
| 171.0176 | kidney | + | 8.04 | ↑ | 1.22 | 1 | C4H8N2O3 | Ureidopropionic acid | HMDB0000026 | 0.77 | 0.61 | NO REACTION | M + K | 171.0167 | −0.001 |
| 600.4706 | kidney | + | 1.53 | ↓ | 1.25 | 1 | C34H68NO6P | CerP(d18:1/16:0) | HMDB0010700 | 1 | 0.83 | [−H2O] | M + H | 600.4751 | 0.005 |
| 556.4439 | kidney | + | 1.52 | ↓ | 1.25 | 1 | UNKNOWN | - | |||||||
| 288.291 | kidney | + | 6.17 | ↓ | 1.23 | 1 | C18H39NO2 | Sphinganine | HMDB0000269 | 1 | 0.74 | [−CH2] | M + H | 288.2897 | −0.001 |
| 166.4847 | kidney | + | 6.96 | ↑ | 1.28 | 0.97 | UNKNOWN | - | |||||||
| 132.1027 | kidney | + | 7 | ↑ | 1.27 | 0.97 | C6H12O2 | L-alpha-Aminobutyric acid | HMDB0000452 | 0.98 | 0.82 | [+NH] | M + H | 132.1019 | −0.001 |
| 609.2826 | kidney | + | 6.15 | ↑ | 1.26 | 0.97 | C26H45NO8S2 | Taurolithocholic acid 3-sulfate | HMDB0002580 | 1 | 0.59 | [+CO] | M + NH4 | 609.2874 | 0.005 |
| 263.1976 | kidney | + | 6.94 | ↑ | 1.26 | 0.97 | C12H23NO4 | Valerylcarnitine | HMDB0013128 | 1 | 0.63 | [+NH3] | M + H | 263.1965 | −0.001 |
| 86.3838 | kidney | + | 6.94 | ↑ | 1.26 | 0.97 | UNKNOWN | - | |||||||
| 160.9176 | kidney | + | 7.82 | ↑ | 1.25 | 0.97 | UNKNOWN | - | |||||||
| 179.0616 | kidney | − | 7.46 | ↑ | 1.33 | 1 | C6H6N4O2 | 1-Methylxanthine | HMDB0010738 | 1 | 0.85 | [+CH2] | M-H | 179.0564 | −0.005 |
| 132.0344 | kidney | − | 7.83 | ↑ | 1.26 | 1 | C4H4O4 | Fumaric acid | HMDB0000134 | 1 | 0.74 | [+NH3] | M-H | 132.0291 | −0.005 |
| 225.0678 | kidney | − | 7.9 | ↑ | 1.21 | 1 | C7H16NO2 | 4-Trimethylammoniobutanoic acid | HMDB0001161 | 1 | 0.73 | [+SO3] | M-H | 225.0665 | −0.001 |
| 130.0914 | kidney | − | 6.86 | ↑ | 1.20 | 1 | C6H10O2 | delta-Hexanolactone | HMDB0000453 | 1 | 1 | [+NH3] | M-H | 130.0863 | −0.005 |
| 267.0796 | kidney | − | 6.6 | ↑ | 1.20 | 1 | C10H12N4O4 | Deoxyinosine | HMDB0000071 | 1 | 0.92 | [+O] | M-H | 267.0724 | −0.007 |
| 124.0114 | kidney | − | 7.25 | ↑ | 1.20 | 1 | UNKNOWN | - | M-H | ||||||
| 180.0716 | kidney | − | 7.34 | ↑ | 1.20 | 1 | C9H8O3 | Phenylpyruvic acid | HMDB0000205 | 1 | 0.82 | [+NH3] | M-H | 180.0655 | −0.006 |
| 289.0737 | kidney | − | 6.03 | ↑ | 1.19 | 1 | C6H12O7 | Galactonic acid | HMDB0000565 | 0.92 | 0.74 | [+C4H2N2O] | M-H | 289.0666 | −0.007 |
| 203.0883 | kidney | − | 6.78 | ↑ | 1.19 | 1 | C11H12N2O2 | L-Tryptophan | HMDB0000929 | 1 | 0.85 | NO REACTION | M-H | 203.0815 | −0.007 |
| 306.0639 | kidney | − | 6.03 | ↑ | 1.18 | 1 | C14H15NO7 | Indoxyl glucuronide | HMDB0010319 | 1 | 0.78 | [−H2] | M-H | 306.0608 | −0.003 |
| 296.8881 | kidney | − | 7.07 | ↑ | 1.18 | 1 | UNKNOWN | - | M-H | ||||||
| 171.0116 | kidney | − | 7.82 | ↑ | 1.18 | 0.97 | C7H8O3S | p-Cresol sulphate | HMDB0011635 | 1 | 0.79 | NO REACTION | M-H | 171.0110 | −0.001 |
| 145.0664 | kidney | − | 7.88 | ↑ | 1.16 | 0.97 | C5H10N2O3 | L-Glutamine | HMDB0000641 | 1 | 0.6 | NO REACTION | M-H | 145.0608 | −0.006 |
| 303.056 | kidney | − | 6.61 | ↑ | 1.15 | 1 | C10H12N2O8 | Orotidine | HMDB0000788 | 1 | 0.93 | [+O] | M-H | 303.0459 | −0.010 |
| 128.9636 | kidney | − | 7.07 | ↑ | 1.14 | 1 | UNKNOWN | - | M-H | ||||||
| 164.0767 | kidney | − | 6.83 | ↑ | 1.13 | 1 | C9H13NO3 | Normetanephrine | HMDB0000819 | 0.87 | 0.83 | [−H2O] | M-H | 164.0706 | −0.006 |
| 243.0685 | kidney | − | 6.03 | ↑ | 1.10 | 1 | C4H8O5 | Threonic acid | HMDB0000943 | 0.96 | 0.85 | [+C5H4N2O] | M-H | 243.0612 | −0.007 |
| 379.107 | kidney | − | 6.07 | ↑ | 1.11 | 1 | C15H15NO4 | L-Thyronine | HMDB0000667 | 0.96 | 0.66 | [+C2H5NO2S] | M-H | 379.0958 | −0.011 |
| 302.1068 | kidney | − | 7.68 | ↑ | 1.11 | 0.97 | C10H17N3O6 | N2-gamma-Glutamylglutamine | HMDB0011738 | 1 | 0.72 | [+CO] | M-H | 302.0983 | −0.009 |
| 294.9404 | plasma | + | 7.16 | ↑ | 1.45 | 1 | Na(NaCOOH)4 | Formate | HMDB0303296 | M+ | 294.9389 | −0.001 | |||
| 362.9282 | plasma | + | 7.16 | ↑ | 1.45 | 0.97 | Na(NaCOOH)5 | Formate | HMDB0303297 | M+ | 362.9263 | −0.002 | |||
| 430.9158 | plasma | + | 7.16 | ↑ | 1.37 | 0.91 | Na(NaCOOH)6 | Formate | HMDB0303298 | M+ | 430.9138 | −0.002 | |||
| 226.9524 | plasma | + | 7.16 | ↑ | 1.09 | 0.91 | Na(NaCOOH)3 | Formate | HMDB0303299 | M+ | 226.9515 | −0.001 | |||
| 332.3335 | plasma | + | 3.36 | ↓ | 1.13 | 1 | UNKNOWN | - | |||||||
| 304.3021 | plasma | + | 3.44 | ↓ | 1.23 | 0.97 | UNKNOWN | - | |||||||
| 326.3804 | plasma | + | 3.17 | ↓ | 1.35 | 0.94 | UNKNOWN | - | |||||||
| 717.0657 | plasma | + | 6.46 | ↑ | 1.36 | 0.91 | UNKNOWN | - |
| UPLC Gradient Conditions | ||
|---|---|---|
| % B-(pos/neg) | Time (min) | Flow (mL/min) |
| 100 | 0 | 0.2 |
| 100 | 2 | 0.2 |
| 5 | 10 | 0.2 |
| 5 | 15 | 0.2 |
| 100 | 15.1 | 0.2 |
| 100 | 22 | 0.2 |
| ESI–QTOF Settings | ||
| Nebulizer gas | N2 | |
| Nebulizer gas pressure | 2 bar | |
| Drying gas | N2 | |
| Drying gas flow | 10 L/min | |
| Drying temperature | 200 °C | |
| Capillary voltage—positive | 3.5 kV | |
| Capillary voltage—positive | 2.5 kV | |
| End plate offset | 0.5 kV | |
| bbCID collision energy | 24 V–36 V (ramp) | |
| m/z scan range | 100–1000 Da | |
| Step | Algorithm | Parameters | Comments |
|---|---|---|---|
| Mass Detection | Centroid Mass detector | Noise level: 500–1500 | adjusted to each dataset |
| Chromatogram building | ADAP Chromatogram builder | Number of scans: 10 | |
| Group Intensity threshold: 1000–2500 | adjusted to each dataset | ||
| Minimum highest intensity: 1000–4000 | adjusted to each dataset | ||
| m/z tolerance: 5 mDa | |||
| Chromatogram deconvolution | Noise amplitude/AUTO mz centre calculation | Minimum peak height: 1100–4100 abs | adjusted to each dataset |
| Peak duration range: 0.1–1 min | |||
| Amplitude of noise: 100–1000 | adjusted to each dataset | ||
| Isotopes | Isotopic peaks grouper | m/z tolerance: 5 mDa | |
| Retention time tolerance: 0.1 | |||
| Maximum charge: 2 | |||
| Adducts | Adducts search | [M + Na], [M + K], [M + MeOH], [M + HCOOH], [M + ACN], etc. | |
| RT tolerance: 0.1 min | |||
| m/z tolerance: 10 mDa | |||
| Max relative-adduct peak height: 100% | |||
| Normalization | Retention time calibration | m/z: tolerance 5 mDa | |
| Retention time tolerance: 0.2 min | |||
| Minimum standard intensity: 5000–10,000 abs | adjusted to each dataset | ||
| Alignment | Join aligner | m/z tolerance: 10 mDa | |
| Retention time tolerance: 0.5 min | |||
| Remove duplicates | Duplicate peak finder/New Average | m/z tolerance: 20 mDa | |
| RT tolerance: 0.8 min | |||
| Gap filling | Peak Finder | Intensity tolerance: 20% | |
| m/z tolerance: 10 mDa | |||
| Retention time tolerance: 0.8 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barla, I.; Efentakis, P.; Lamprou, S.; Gavriatopoulou, M.; Dimopoulos, M.-A.; Terpos, E.; Andreadou, I.; Thomaidis, N.; Gikas, E. An Untargeted Metabolomics Approach on Carfilzomib-Induced Nephrotoxicity. Molecules 2022, 27, 7929. https://doi.org/10.3390/molecules27227929
Barla I, Efentakis P, Lamprou S, Gavriatopoulou M, Dimopoulos M-A, Terpos E, Andreadou I, Thomaidis N, Gikas E. An Untargeted Metabolomics Approach on Carfilzomib-Induced Nephrotoxicity. Molecules. 2022; 27(22):7929. https://doi.org/10.3390/molecules27227929
Chicago/Turabian StyleBarla, Ioanna, Panagiotis Efentakis, Sofia Lamprou, Maria Gavriatopoulou, Meletios-Athanasios Dimopoulos, Evangelos Terpos, Ioanna Andreadou, Nikolaos Thomaidis, and Evangelos Gikas. 2022. "An Untargeted Metabolomics Approach on Carfilzomib-Induced Nephrotoxicity" Molecules 27, no. 22: 7929. https://doi.org/10.3390/molecules27227929