
Use of MALDI-TOF MS to Discriminate between Aflatoxin B1-Producing and Non-Producing Strains of Aspergillus flavus
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Detection of AFB1
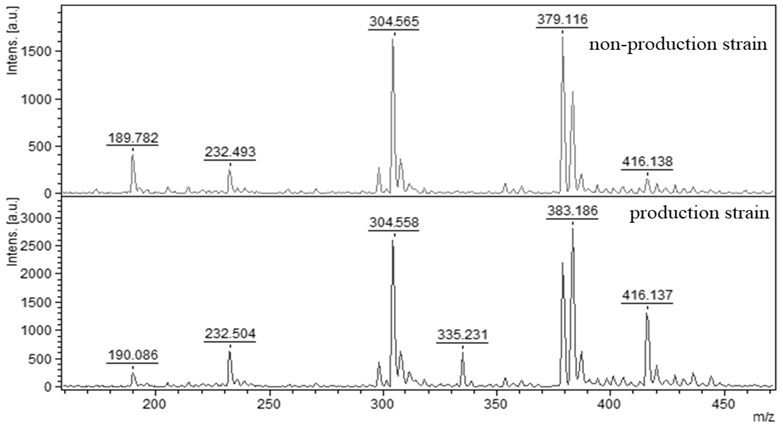
2.2. Confirmation of AFB1 Producing Strains of Aspergillus flavus
2.2.1. Direct AFB1 Detection from TLC Scrap
2.2.2. Direct AFB1 Detection from A. flavus Mycelia
2.2.3. Direct AFB1 Detection from Medium around the A. flavus
2.2.4. Direct AFB1 Detection from Grain Samples
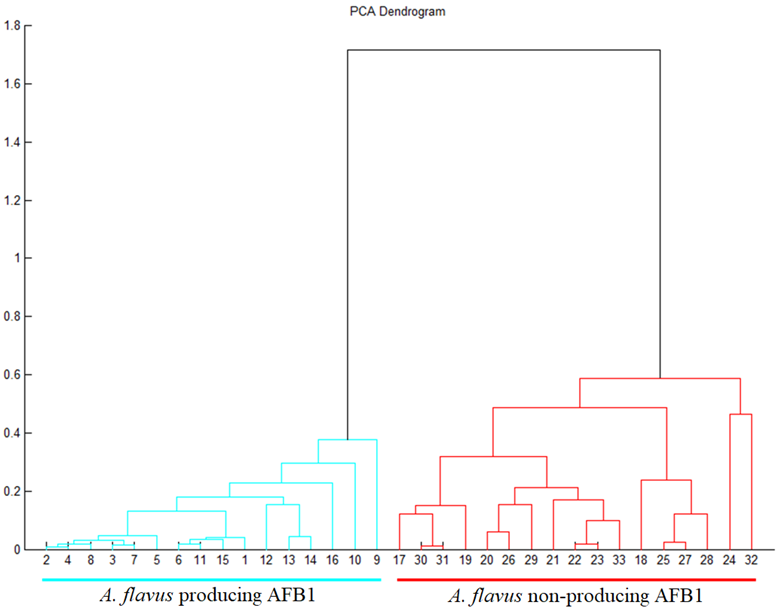
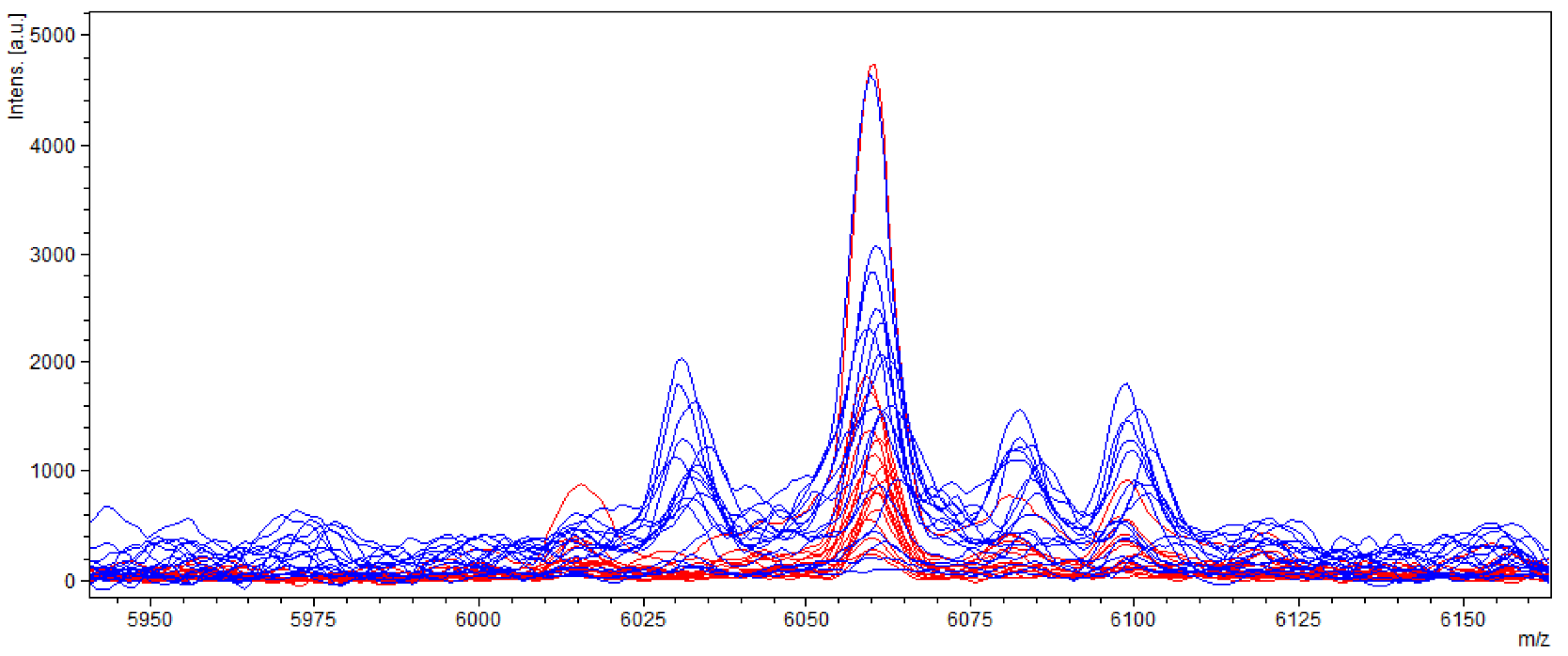
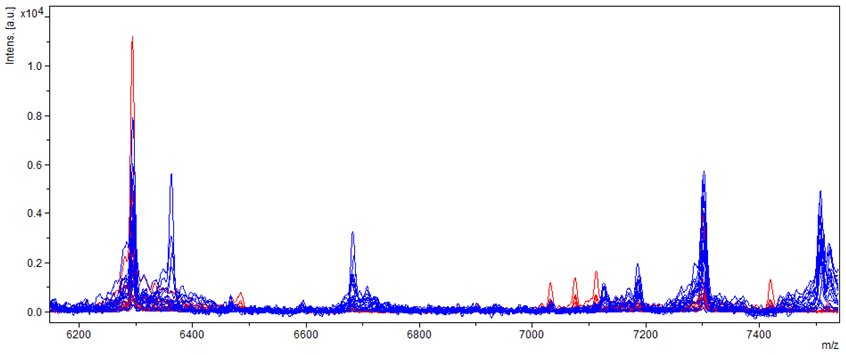
2.3. Indirect Detection of Aflatoxin-Producing Strains of A. flavus by Mass Spectrum Discrimination
Aspergillus flavus Entire Spectrum Observation
3. Materials and Methods
3.1. AFB1 Analysis
3.1.1. Preparation of AFB1
3.1.2. MALDI-TOF Detection of AFB1
3.1.3. Analysis of Mass Spectra
3.2. MALDI-TOF Direct Detection of AFB1 in Producing Strains Aspergillus flavus
3.2.1. Microorganisms, Cultivation, and Identification
3.2.2. MALDI-TOF MS A. flavus Identification
3.2.3. Verification of AFB1 Producers by TLC Method
3.2.4. Mass Spectrometry Methodology
3.3. MALDI-TOF MS Detection AFB1 in Grain Samples Directly
3.4. MALDI-TOF Detection of AFB1 Producing Strains of Aspergillus flavus Based on Comparison of Entire Mass Spectra (Fingerperint Method)
3.4.1. Proteins Isolation and MALDI-TOF Protocol
3.4.2. Creating an Aspergillus flavus Local Database
3.4.3. Verification of Reproducibility
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodrigues, A.I.; Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactants as Biocontrol Agents Against Mycotoxigenic Fungi. In Biosurfactants for a Sustainable Future; John Wiley and Sons: Hoboken, NJ, USA, 2021. [Google Scholar]
- Klapec, T.; Wócik-Falta, A.; Farian, E.; Kowalczyk, K.; Cholewa, G.; Cholewa, A.; Dutkiewicz, J. Mycobiota of Berry Fruits—Levels of Filamentous Fungi and Mycotoxins, Composition of Fungi, and Analysis of Potential Health Risk for Consumers. Ann. Agric. Environ. Med. 2022, 29, 28. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Nisa, S.; Daud, S. Mycotoxins in Environment and Its Health Implications. In Hazardous Environmental Micro-Pollutants, Health Impacts and Allied Treatment Technologies; Springer: Cham, Switzerland, 2022; pp. 289–318. [Google Scholar]
- Gallo, M.; Ferrara, L.; Calogero, A.; Montesano, D.; Naviglio, D. Relationships between Food and Diseases: What to Know to Ensure Food Safety. Food Res. Int. 2020, 137, 109414. [Google Scholar] [CrossRef] [PubMed]
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef] [Green Version]
- Winter, G.; Pereg, L. A Review on the Relation between Soil and Mycotoxins: Effect of Aflatoxin on Field, Food and Finance. Eur. J. Soil Sci. 2019, 70, 882–897. [Google Scholar] [CrossRef]
- Singh, P.; Orbach, M.J.; Cotty, P.J. Aspergillus Texensis: A Novel Aflatoxin Producer with s Morphology from the United States. Toxins 2018, 10, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klich, M.A.; Mullaney, E.J.; Daly, C.B.; Cary, J.W. Molecular and Physiological Aspects of Aflatoxin and Sterigmatocystin Biosynthesis by Aspergillus tamarii and A. ochraceoroseus. Appl. Microbiol. Biotechnol. 2000, 53, 605–609. [Google Scholar] [CrossRef]
- Peterson, S.W.; Ito, Y.; Horn, B.W.; Goto, T. Aspergillus Bombycis, a New Aflatoxigenic Species and Genetic Variation in Its Sibling Species, A. nomius. Mycologia 2001, 93, 689. [Google Scholar] [CrossRef]
- Fumagalli, F.; Ottoboni, M.; Pinotti, L.; Cheli, F. Integrated Mycotoxin Management System in the Feed Supply Chain: Innovative Approaches. Toxins 2021, 13, 572. [Google Scholar] [CrossRef]
- Darwish, A. Fungal Mycotoxins and Natural Antioxidants: Two Sides of the Same Coin and Significance in Food Safety. Microb. Biosyst. 2019, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Litoriya, N.S.; Modi, A. Mycotoxin-Associated Food Safety Concerns of Agriculture Crops. In Food Security and Plant Disease Management; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF Mass Spectrometry: An Emerging Technology for Microbial Identification and Diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Oyedeji, A.B.; Green, E.; Adebiyi, J.A.; Ogundele, O.M.; Gbashi, S.; Adefisoye, M.A.; Oyeyinka, S.A.; Adebo, O.A. Metabolomic Approaches for the Determination of Metabolites from Pathogenic Microorganisms: A Review. Food Res. Int. 2021, 140, 110042. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Chen, Y.C. Characterization of Intact Penicillium Spores by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Chalupová, J.; Raus, M.; Sedlářová, M.; Šebela, M. Identification of Fungal Microorganisms by MALDI-TOF Mass Spectrometry. Biotechnol. Adv. 2014, 32, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Hleba, L.; Císarová, M.; Shariati, M.A.; Tancinová, D. Detection of Mycotoxins Using MALDI-TOF Mass Spectrometry. J. Microbiol. Biotechnol. Food Sci. 2017, 7, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Elosta, S.; Gajdošová, D.; Hégrová, B.; Havel, J. MALDI TOF Mass Spectrometry of Selected Mycotoxins in Barley. J. Appl. Biomed. 2007, 5, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Blechová, P.; Havlová, P.; Gajdošová, D.; Havel, J. New Possibilities of Matrix-Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry to Analyze Barley Malt Quality. Highly Sensitive Detection of Mycotoxins. Environ. Toxicol. 2006, 21, 403–408. [Google Scholar] [CrossRef]
- Lanyasunya, T.P.; Wamae, L.W.; Musa, H.H.; Olowofeso, O.; Lokwaleput, I.K. The Risk of Mycotoxins Contamination of Dairy Feed and Milk on Smallholder Dairy Farms in Kenya. Pak. J. Nutr. 2005, 4, 162–169. [Google Scholar] [CrossRef]
- Trail, F.; Mahanti, N.; Rarick, M.; Mehigh, R.; Liang, S.H.; Zhou, R.; Linz, J.E. Physical and Transcriptional Map of an Aflatoxin Gene Cluster in Aspergillus Parasiticus and Functional Disruption of a Gene Involved Early in the Aflatoxin Pathway. Appl. Environ. Microbiol. 1995, 61, 2665–2673. [Google Scholar] [CrossRef] [Green Version]
- Kew, M.C. Aflatoxins as a Cause of Hepatocellular Carcinoma. J. Gastrointest. Liver Dis. 2013, 22, 305–310. [Google Scholar]
- Feucherolles, M.; Poppert, S.; Utzinger, J.; Becker, S.L. MALDI-TOF Mass Spectrometry as a Diagnostic Tool in Human and Veterinary Helminthology: A Systematic Review. Parasit Vectors 2019, 12, 1–13. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, Z.; Guo, Y. Chemical and Biochemical Applications of MALDI TOF-MS Based on Analyzing the Small Organic Compounds. In Applications of MALDI-TOF Spectroscopy; Topics in Current Chemistry Series; Springer: Berlin/Heidelberg, Germany, 2012; Volume 331, pp. 165–192. [Google Scholar]
- Catharino, R.R.; de Marques, L.A.; Santos, L.S.; Baptista, A.S.; Glória, E.M.; Calori-Domingues, M.A.; Facco, E.M.P.; Eberlin, M.N. Aflatoxin Screening by MALDI-TOF Mass Spectrometry. Anal. Chem. 2005, 77, 8155–8157. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Gómez, A.; Anaya, I.; Díaz, J.; Broto, F.; Agut, M.; Comellas, L. Determination of Aflatoxins B1, G1, B2 and G2 in Medicinal Herbs by Liquid Chromatography-Tandem Mass Spectrometry. J. Chromatogr. A 2004, 1048, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Var, I.; Kabak, B.; Gök, F. Survey of Aflatoxin B1 in Helva, a Traditional Turkish Food, by TLC. Food Control 2007, 18, 59–62. [Google Scholar] [CrossRef]
- Reddy, K.R.N.; Reddy, C.S.; Muralidharan, K. Characterization of Aflatoxin B1 Produced by Aspergillus Flavus Isolated from Discolored Rice Grains. J. Mycol. Plant Pathol. 2005, 35, 470–474. [Google Scholar]
- Wu, L.; Zhou, M.; Wang, Y.; Liu, J. Nanozyme and Aptamer- Based Immunosorbent Assay for Aflatoxin B1. J. Hazard. Mater. 2020, 399, 123154. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, U.; Ahmad, M.; Anwar-ul-Haq; Mohsin Iqbal, M. Determination of Aflatoxin-B1 in Poultry Feed and Its Components Employing Enzyme-Linked Immunosorbent Assay (Elisa). Toxicol. Environ. Chem. 2004, 86, 213–218. [Google Scholar] [CrossRef]
- Lawellin, D.W.; Grant, D.W.; Joyce, B.K. Enzyme-Linked Immunosorbent Analysis for Aflatoxin B1. Appl. Environ. Microbiol. 1977, 34, 94–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdanpanah, H.; Zarghi, A.; Shafaati, A.R.; Foroutan, S.M.; Aboul-Fathi, F.; Khoddam, A.; Nazari, F.; Shaki, F. Analysis of Aflatoxin B1 in Iranian Foods Using HPLC and a Monolithic Column and Estimation of Its Dietary Intake. Iran J. Pharm. Res. 2013, 12, 83–89. [Google Scholar]
- Ammida, N.H.S.; Micheli, L.; Piermarini, S.; Moscone, D.; Palleschi, G. Detection of Aflatoxin B1 in Barley: Comparative Study of Immunosensor and HPLC. Anal. Lett. 2006, 39, 1559–1572. [Google Scholar] [CrossRef]
- Jangampalli Adi, P.; Matcha, B. Analysis of Aflatoxin B1 in Contaminated Feed, Media, and Serum Samples of Cyprinus Carpio L. by High-Performance Liquid Chromatography. Food Qual. Saf. 2018, 2, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Císarová, M.; Hleba, L.; Medo, J.; Tančinová, D.; Mašková, Z.; Čuboň, J.; Kováčik, A.; Foltinová, D.; Božik, M.; Klouček, P. The In Vitro and In Situ Effect of Selected Essential Oils in Vapour Phase against Bread Spoilage Toxicogenic Aspergilli. Food Control 2020, 110, 107007. [Google Scholar] [CrossRef]
- Tsai, F.J.; Chen, S.Y.; Liu, Y.C.; Liao, H.Y.; Chen, C.J. The Comparison of CHCA Solvent Compositions for Improving LC-MALDI Performance and Its Application to Study the Impact of Aflatoxin B1 on the Liver Proteome of Diabetes Mellitus Type 1 Mice. PLoS ONE 2017, 12, e0181423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, V.; Adhikary, R.; Maiti, P.K.; Mandal, S.; Mandal, V. Morpho-Biochemical and Molecular Characterization of Two New Strains of Aspergillus Fumigatus NHF-01 and A. Fumigatus PPR-01 Producing Broad-Spectrum Antimicrobial Compounds. Braz. J. Microbiol. 2021, 52, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Tuzimski, T. Application of Different Modes of Thin-Layer Chromatography and Mass Spectrometry for the Separation and Detection of Large and Small Biomolecules. J. Chromatogr. A 2011, 1218, 8799–8812. [Google Scholar] [CrossRef]
- Zimcik, P.; Malkova, A.; Hruba, L.; Miletin, M.; Novakova, V. Bulky 2,6-Diphenylphenylsulfanyl Substituents Efficiently Inhibit Aggregation in Phthalocyanines and Tetrapyrazinoporphyrazines and Control Their Photophysical and Electrochemical Properties. Dye. Pigment. 2017, 136, 715–723. [Google Scholar] [CrossRef]
- Davis, N.D.; Diener, U.L.; Eldridge, D.W. Production of Aflatoxins B1 and G1 by Aspergillus flavus in a Semisynthetic Medium. Appl. Microbiol. 1966, 14, 378–380. [Google Scholar] [CrossRef]
- Liu, D.L.; Yao, D.S.; Liang, R.; Ma, L.; Cheng, W.Q.; Gu, L.Q. Detoxification of Aflatoxin B1 by Enzymes Isolated from Armillariella tabescens. Food Chem. Toxicol. 1998, 36, 563–574. [Google Scholar] [CrossRef]
- Gell, R.M.; Carbone, I. HPLC Quantitation of Aflatoxin B1 from Fungal Mycelium Culture. J. Microbiol. Methods 2019, 158, 14–17. [Google Scholar] [CrossRef]
- Wilson, B.J. Toxins Other than Aflatoxins Produced by Aspergillus flavus. Bacteriol. Rev. 1966, 30, 478–484. [Google Scholar] [CrossRef]
- Gourama, H.; Bullerman, L.B. Aspergillus flavus and Aspergillus parasiticus: Aflatoxigenic Fungi of Concern in Foods and Feeds: A Review. J. Food Prot. 1995, 58, 1395–1404. [Google Scholar] [CrossRef]
- Helal, G.E.D.A.; Sarhan, M.M.; Abu Shahla, A.N.K.; Abou El-Khair, E.K. Effects of Cymbopogon citratus L. Essential Oil on the Growth, Morphogenesis and Aflatoxin Production of Aspergillus flavus ML2-Strain. J. Basic Microbiol. 2007, 47, 5–15. [Google Scholar] [CrossRef]
- Garon, D.; Richard, E.; Sage, L.; Bouchart, V.; Pottier, D.; Lebailly, P. Mycoflora and Multimycotoxin Detection in Corn Silage: Experimental Study. J. Agric. Food Chem. 2006, 54, 3479–3484. [Google Scholar] [CrossRef]
- Jerome Jeyakumar, J.M.; Zhang, M.; Thiruvengadam, M. Determination of Mycotoxins by HPLC, LC-ESI-MS/MS, and MALDI-TOF MS in Fusarium Species-Infected Sugarcane. Microb. Pathog. 2018, 123, 98–110. [Google Scholar] [CrossRef]
- Sivagnanam, K.; Komatsu, E.; Rampitsch, C.; Perreault, H.; Gräfenhan, T. Rapid Screening of Alternaria Mycotoxins Using MALDI-TOF Mass Spectrometry. J. Sci. Food Agric. 2017, 97, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Kobylis, P.; Stepnowski, P.; Caban, M. Review of the Applicability of Ionic Liquid Matrices for the Quantification of Small Molecules by MALDI MS. Microchem. J. 2021, 164, 105983. [Google Scholar] [CrossRef]
- Carbonnelle, E.; Mesquita, C.; Bille, E.; Day, N.; Dauphin, B.; Beretti, J.L.; Ferroni, A.; Gutmann, L.; Nassif, X. MALDI-TOF Mass Spectrometry Tools for Bacterial Identification in Clinical Microbiology Laboratory. Clin. Biochem. 2011, 44, 104–109. [Google Scholar] [CrossRef]
- Li, Y.; Shan, M.; Zhu, Z.; Mao, X.; Yan, M.; Chen, Y.; Zhu, Q.; Li, H.; Gu, B. Application of MALDI-TOF MS to Rapid Identification of Anaerobic Bacteria. BMC Infect. Dis. 2019, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Paauw, A.; Jonker, D.; Roeselers, G.; Heng, J.M.E.; Mars-Groenendijk, R.H.; Trip, H.; Molhoek, E.M.; Jansen, H.J.; van der Plas, J.; de Jong, A.L.; et al. Rapid and Reliable Discrimination between Shigella Species and Escherichia Coli Using MALDI-TOF Mass Spectrometry. Int. J. Med. Microbiol. 2015, 305, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Cassagne, C.; Normand, A.C.; L’Ollivier, C.; Ranque, S.; Piarroux, R. Performance of MALDI-TOF MS Platforms for Fungal Identification. Mycoses 2016, 59, 678–690. [Google Scholar] [CrossRef]
- Patel, R. A Moldy Application of MALDI: MALDI-ToF Mass Spectrometry for Fungal Identification. J. Fungi 2019, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Bader, O. MALDI-TOF-MS-Based Species Identification and Typing Approaches in Medical Mycology. Proteomics 2013, 13, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Taverna, C.G.; Mazza, M.; Bueno, N.S.; Alvarez, C.; Amigot, S.; Andreani, M.; Azula, N.; Barrios, R.; Fernández, N.; Fox, B.; et al. Development and Validation of an Extended Database for Yeast Identification by Maldi-Tof Ms in Argentina. Med. Mycol. 2019, 57, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, M.; Mewes, A.; Maggipinto, M.; Schmidt, W.; Messelhäußer, U.; Balsliemke, J.; Hörmansdorfer, S.; Busch, U.; Huber, I. MALDI-TOF MS Based Identification of Food-Borne Yeast Isolates. J. Microbiol. Methods 2014, 106, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Fidelis, C.E.; Franke, M.; de Abreu, L.C.R.; Jagielski, T.; Ribeiro, M.G.; dos Santos, M.v.; Gonçalves, J.L. MALDI-TOF MS Identification of Prototheca Algae Associated with Bovine Mastitis. J. Vet. Diagn. Investig. 2021, 33, 1168–1171. [Google Scholar] [CrossRef] [PubMed]
- Volta, P.; Riccardi, N.; Lauceri, R.; Tonolla, M. Discrimination of Freshwater Fish Species by Matrix-Assisted Laser Desorption/Ionization- Time of Flight Mass Spectrometry (MALDI-TOF MS): A Pilot Study. J. Limnol. 2012, 71, e17. [Google Scholar] [CrossRef]
- Yssouf, A.; Socolovschi, C.; Leulmi, H.; Kernif, T.; Bitam, I.; Audoly, G.; Almeras, L.; Raoult, D.; Parola, P. Identification of Flea Species Using MALDI-TOF/MS. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 153–157. [Google Scholar] [CrossRef]
- Welker, M.; van Belkum, A.; Girard, V.; Charrier, J.P.; Pincus, D. An Update on the Routine Application of MALDI-TOF MS in Clinical Microbiology. Expert Rev. Proteom. 2019, 16, 695–710. [Google Scholar] [CrossRef]
- Seng, P.; Rolain, J.M.; Fournier, P.E.; la Scola, B.; Drancourt, M.; Raoult, D. MALDI-TOF-Mass Spectrometry Applications in Clinical Microbiology. Future Microbiol. 2010, 5, 1733–1754. [Google Scholar] [CrossRef]
- Angeletti, S. Matrix Assisted Laser Desorption Time of Flight Mass Spectrometry (MALDI-TOF MS) in Clinical Microbiology. J. Microbiol. Methods 2017, 138, 20–29. [Google Scholar] [CrossRef]
- Nakano, S.; Matsumura, Y.; Ito, Y.; Fujisawa, T.; Chang, B.; Suga, S.; Kato, K.; Yunoki, T.; Hotta, G.; Noguchi, T.; et al. Development and Evaluation of MALDI-TOF MS-Based Serotyping for Streptococcus Pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2191–2198. [Google Scholar] [CrossRef] [Green Version]
- Kyritsi, M.A.; Kristo, I.; Hadjichristodoulou, C. Serotyping and Detection of Pathogenecity Loci of Environmental Isolates of Legionella Pneumophila Using MALDI-TOF MS. Int. J. Hyg. Environ. Health 2020, 224, 113441. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Yamamoto, N.; Takahashi, H.; Tamura, H. Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) Can Precisely Discriminate the Lineages of Listeria Monocytogenes and Species of Listeria. PLoS ONE 2016, 11, e0159730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojima-Kato, T.; Yamamoto, N.; Nagai, S.; Shima, K.; Akiyama, Y.; Ota, J.; Tamura, H. Application of Proteotyping Strain SolutionTM Ver. 2 Software and Theoretically Calculated Mass Database in MALDI-TOF MS Typing of Salmonella Serotype. Appl. Microbiol. Biotechnol. 2017, 101, e015973. [Google Scholar] [CrossRef] [PubMed]
- Zautner, A.E.; Masanta, W.O.; Tareen, A.M.; Weig, M.; Lugert, R.; Groß, U.; Bader, O. Discrimination of Multilocus Sequence Typing-Based Campylobacter Jejuni Subgroups by MALDI-TOF Mass Spectrometry. BMC Microbiol. 2013, 13, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Kim, I.; Chung, S.H.; Chung, Y.; Han, M.; Kim, J.S. Rapid Discrimination of Methicillin-Resistant Staphylococcus aureus by MALDI-TOF MS. Pathogens 2019, 8, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hleba, L.; Hlebová, M.; Kováčik, A.; Čuboň, J.; Medo, J. Carbapenemase Producing Klebsiella pneumoniae (Kpc): What Is the Best Maldi-Tof Ms Detection Method. Antibiotics 2021, 10, 1549. [Google Scholar] [CrossRef]
- Hrabák, J.; Chudáčková, E.; Walková, R. Matrix-Assisted Laser Desorption Ionization-Time of Flight (MALDITOF) Mass Spectrometry for Detection of Antibiotic Resistance Mechanisms: From Research to Routine Diagnosis. Clin. Microbiol. Rev. 2013, 26, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Caputo, P.; di Martino, M.; Perfetto, B.; Iovino, F.; Donnarumma, G. Use of MALDI-TOF MS to Discriminate between Biofilm-Producer and Non-Producer Strains of Staphylococcus epidermidis. Int. J. Environ. Res. Public Health 2018, 15, 1695. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, P.A.D.F.; Menezes, R.P.; Penatti, M.P.A.; Moreira, T.A.; Pimenta, J.P.; Silva, N.B.S.; Röder, D.V.D.B. Rapid Detection of Biofilm-Producing Candida Species via MALDI-TOF Mass Spectrometry. J. Appl. Microbiol. 2021, 131, 2049–2060. [Google Scholar] [CrossRef]
- Kuhns, M.; Zautner, A.E.; Rabsch, W.; Zimmermann, O.; Weig, M.; Bader, O.; Groß, U. Rapid Discrimination of Salmonella enterica Serovar Typhi from Other Serovars by MALDI-TOF Mass Spectrometry. PLoS ONE 2012, 7, e40004. [Google Scholar] [CrossRef]
- Calderaro, A.; Piccolo, G.; Gorrini, C.; Montecchini, S.; Buttrini, M.; Rossi, S.; Piergianni, M.; de Conto, F.; Arcangeletti, M.C.; Chezzi, C.; et al. Leptospira Species and Serovars Identified by MALDI-TOF Mass Spectrometry after Database Implementation. BMC Res. Notes 2014, 7, 330. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Yamamoto, N.; Suzuki, M.; Fukunaga, T.; Tamura, H. Discrimination of Escherichia Coli O157, O26 and O111 from Other Serovars by MALDI-TOF MS Based on the S10-Germs Method. PLoS ONE 2014, 9, e113458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hleba, L.; Charousova, I.; Cisarova, M.; Kovacik, A.; Kormanec, J.; Medo, J.; Bozik, M.; Javorekova, S. Rapid Identification of Streptomyces Tetracycline Producers by MALDI-TOF Mass Spectrometry. J. Environ. Sci. Health Part A 2018, 53, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Nagai, S.; Fujita, A.; Ido, Y.; Kato, K.; Saito, A.; Moriya, Y.; Tomimatsu, Y.; Kaneta, N.; Tsujimoto, Y.; et al. Discrimination of Psychrotolerant Bacillus cereus Group Based on MALDI-TOF MS Analysis of Ribosomal Subunit Proteins. Food Microbiol. 2020, 91, 103542. [Google Scholar] [CrossRef] [PubMed]
- Paauw, A.; Trip, H.; Niemcewicz, M.; Sellek, R.; Heng, J.M.E.; Mars-Groenendijk, R.H.; de Jong, A.L.; Majchrzykiewicz-Koehorst, J.A.; Olsen, J.S.; Tsivtsivadze, E. OmpU as a Biomarker for Rapid Discrimination between Toxigenic and Epidemic Vibrio cholerae O1/O139 and Non-Epidemic Vibrio cholerae in a Modified MALDI-TOF MS Assay. BMC Microbiol. 2014, 14, 158. [Google Scholar] [CrossRef] [Green Version]
- Verwer, P.E.B.; van Leeuwen, W.B.; Girard, V.; Monnin, V.; van Belkum, A.; Staab, J.F.; Verbrugh, H.A.; Bakker-Woudenberg, I.A.J.M.; van de Sande, W.W.J. Discrimination of Aspergillus lentulus from Aspergillus fumigatus by Raman Spectroscopy and MALDI-TOF MS. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 245–251. [Google Scholar] [CrossRef]
- Hettick, J.M.; Green, B.J.; Buskirk, A.D.; Kashon, M.L.; Slaven, J.E.; Janotka, E.; Blachere, F.M.; Schmechel, D.; Beezhold, D.H. Discrimination of Aspergillus Isolates at the Species and Strain Level by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Fingerprinting. Anal. Biochem. 2008, 380, 276–281. [Google Scholar] [CrossRef]
- Yoon, E.J.; Lee, E.H.; Hwang, D.H.; Lee, H.; Baek, J.H.; Jeong, S.H. Direct Detection of Intact Klebsiella pneumoniae Carbapenemases Produced by Enterobacterales Using MALDI-TOF MS. J. Antimicrob. Chemother. 2020, 75, 1174–1181. [Google Scholar] [CrossRef]
- de Carolis, E.; Soldini, S.; la Rosa, M.; Nucci, F.; Posteraro, B.; Sanguinetti, M. BIOF–HILO Assay: A New MALDI–TOF Mass Spectrometry Based Method for Discriminating between High-and Low-Biofilm-Producing Candida parapsilosis Isolates. Front. Microbiol. 2019, 10, 2046. [Google Scholar] [CrossRef] [Green Version]
- Dortet, L.; Bonnin, R.A.; Pennisi, I.; Gauthier, L.; Jousset, A.B.; Dabos, L.; Furniss, R.C.D.; Mavridou, D.A.I.; Bogaerts, P.; Glupczynski, Y.; et al. Rapid Detection and Discrimination of Chromosome-And MCR-Plasmid-Mediated Resistance to Polymyxins by MALDI-TOF MS in Escherichia coli: The MALDIxin Test. J. Antimicrob. Chemother. 2018, 73, 3359–3367. [Google Scholar] [CrossRef]
- Schaumann, R.; Knoop, N.; Genzel, G.H.; Losensky, K.; Rosenkranz, C.; Stîngu, C.S.; Schellenberger, W.; Rodloff, A.C.; Eschrich, K. A Step towards the Discrimination of Beta-Lactamase-Producing Clinical Isolates of Enterobacteriaceae and Pseudomonas aeruginosa by MALDI-TOF Mass Spectrometry. Med. Sci. Monit. 2012, 18, MT71–MT77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolters, M.; Rohde, H.; Maier, T.; Belmar-Campos, C.; Franke, G.; Scherpe, S.; Aepfelbacher, M.; Christner, M. MALDI-TOF MS Fingerprinting Allows for Discrimination of Major Methicillin-Resistant Staphylococcus aureus Lineages. Int. J. Med. Microbiol. 2011, 301, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Hleba, L.; Drab, S.; Cisarova, M.; Cubon, J. Barley Varieties Discrimination Using MALDI-TOF Mass Spectrometry. J. Microbiol. Biotechnol. Food Sci. 2019, 8, 1347–1351. [Google Scholar] [CrossRef]
- Šalplachta, J.; Bobálová, J. MALDI-TOF Mass Spectrometry of Hordeins: Rapid Approach for Identification of Malting Barley Varieties. J. Mass Spectrom. 2009, 44, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Reeve, M.A.; Pollard, K.M. Discrimination between Regional Biotypes of Impatiens Glandulifera Using a Simple MALDI-TOF MS-Based Method for Use with Seeds. Plant Methods 2019, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Granzotto, C.; Sutherland, K.; Arslanoglu, J.; Ferguson, G.A. Discrimination of Acacia Gums by MALDI-TOF MS: Applications to Micro-Samples from Works of Art. Microchem. J. 2019, 144, 229–241. [Google Scholar] [CrossRef]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi. In CBS Laboratory Manual Series 2; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010. [Google Scholar]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus Section Flavi and Their Production of Aflatoxins, Ochratoxins and Other Mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, Identification and Nomenclature of the Genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Iamanaka, B.T.; de Souza Lopes, A.; Martins, L.M.; Frisvad, J.C.; Medina, A.; Magan, N.; Sartori, D.; Massi, F.P.; Fungaro, M.H.P.; Taniwaki, M.H. Aspergillus Section Flavi Diversity and the Role of A. novoparasiticus in Aflatoxin Contamination in the Sugarcane Production Chain. Int. J. Food Microbiol. 2019, 293, 17–23. [Google Scholar] [CrossRef]
- Císarová, M.; Tančinová, D.; Medo, J.; Kačániová, M. The In Vitro Effect of Selected Essential Oils on the Growth and Mycotoxin Production of Aspergillus Species. J. Environ. Sci. Health B 2016, 51, 668–674. [Google Scholar] [CrossRef]
- Srivastava, B.; Singh, P.; Shukla, R.; Dubey, N.K. A Novel Combination of the Essential Oils of Cinnamomum camphora and Alpinia galanga in Checking Aflatoxin B1 Production by a Toxigenic Strain of Aspergillus flavus. World J. Microbiol. Biotechnol. 2008, 24, 693–697. [Google Scholar] [CrossRef]
- Loucif, L.; Bendjama, E.; Gacemi-Kirane, D.; Rolain, J.M. Rapid Identification of Streptomyces Isolates by MALDI-TOF MS. Microbiol. Res. 2014, 169, 940–947. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hleba, L.; Hlebova, M.; Kovacik, A.; Petrova, J.; Maskova, Z.; Cubon, J.; Massanyi, P. Use of MALDI-TOF MS to Discriminate between Aflatoxin B1-Producing and Non-Producing Strains of Aspergillus flavus. Molecules 2022, 27, 7861. https://doi.org/10.3390/molecules27227861
Hleba L, Hlebova M, Kovacik A, Petrova J, Maskova Z, Cubon J, Massanyi P. Use of MALDI-TOF MS to Discriminate between Aflatoxin B1-Producing and Non-Producing Strains of Aspergillus flavus. Molecules. 2022; 27(22):7861. https://doi.org/10.3390/molecules27227861
Chicago/Turabian StyleHleba, Lukas, Miroslava Hlebova, Anton Kovacik, Jana Petrova, Zuzana Maskova, Juraj Cubon, and Peter Massanyi. 2022. "Use of MALDI-TOF MS to Discriminate between Aflatoxin B1-Producing and Non-Producing Strains of Aspergillus flavus" Molecules 27, no. 22: 7861. https://doi.org/10.3390/molecules27227861